PRINCIPIOS DEL METABOLISMO MICROBIANO
Anuncio

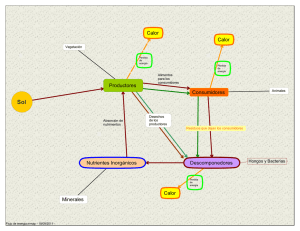
CAPITULO I PRINCIPIOS DEL METABOLISMO MICROBIANO 1.1. CARACTERÍSTICAS GENERALES DE LOS GRUPOS MICROBIANOS Las bacterias se encuentran en casi todos los ambientes e intervienen en varios procesos biológicos. El crecimiento microbiano requiere la formación de estructuras complejas como proteínas, ácidos nucleicos, polisacáridos y lípidos a partir de elementos preformados en el medio de crecimiento o ser sintetizados por la propia célula, a su vez, este crecimiento necesita de una fuente de energía para ser llevado a efecto, todo este proceso se designa con el nombre de metabolismo, que se define como todas las transformaciones químicas que ocurren en una célula. Todos los seres vivos llevan a cabo el procesamiento de los nutrientes que los mantienen vivos. A este conjunto de procesos, se le conoce como metabolismo y consiste de un gran número de reacciones químicas destinadas a transformar las moléculas nutritivas en elementos que posteriormente serán utilizados para la síntesis de los componentes estructurales; como pueden ser las proteínas. Otra parte importante del metabolismo es la de transformar y conservar la energía que está contenida en una reacción química en algún proceso que requiera de energía, como puede ser el trabajo o el movimiento. Figura N° 1.1. Es evidente que los nutrientes son transformados cuando entran en un organismo, ya que en ningún caso el alimento contiene todas las moléculas que una célula requiere. Esto se vio con claridad al observar el crecimiento normal de levaduras en un medio de cultivo que sólo contenía glucosa como única fuente de energía. Así pues, se pensó que la síntesis de todos los componentes celulares se llevaba a cabo en el interior de las levaduras. Hoy se conoce que las transformaciones que sufre la glucosa no ocurren en un solo paso, sino que, por el contrario, se forman varios productos intermedios que en muchas ocasiones no tienen una función específica a no ser la de formar parte de lo que se conoce como vía metabólica. La transformación de los nutrientes en 23 compuestos útiles para la subsistencia de un organismo se lleva a cabo por medio de las reacciones químicas que realizan unas proteínas conocidas como enzimas. El término metabolismo se refiere al conjunto de reacciones químicas que tiene lugar en la célula, y tiene tres funciones específicas a saber: - Obtener energía química del entorno, almacenarla, para utilizar luego en diferentes funciones celulares, - Convertir los nutrientes exógenos en unidades precursoras de los componentes macromoleculares de la célula bacteriana, - Formar y degradar moléculas necesarias para funciones celulares específicas, como por ejemplo, movilidad y captación de nutrientes. Figura N° 1.1. Una Visión Simplificada del Metabolismo Celular. Fuente: (M.T. Madigan, J.M. Martinko y J. Parker., 2002). 24 La energía liberada en las reacciones catabólicas se usa para fosforilar ADP, generando ATP. La energía almacenada en el ATP se utiliza en la mayoría de los trabajos celulares. Por lo tanto, el ATP acopla los procesos productores de energía de la célula a los consumidores de energía. Figura Nº 1.2. Todos los procesos que ocurren en la célula o bacteria requieren de energía. Esta energía está almacenada como moléculas de ATP, que se forma a partir de ADP y fosfato inorgánico. Figura Nº 1.2. El anabolismo y el catabolismo. Fuente: (Dreyfus Cortés Georges, 1995) 25 El metabolismo tiene lugar a través de secuencias de reacciones catalizadas enzimáticamente, y se divide en anabolismo y catabolismo. El proceso por el cual la célula bacteriana sintetiza sus propios componentes se conoce como anabolismo, y como resulta en la producción de nuevo material celular, también se denomina biosíntesis. La biosíntesis es un proceso que requiere energía, por lo tanto las bacterias deben ser capaces de obtenerla de su entorno para crecer y, eventualmente, multiplicarse. El conjunto de reacciones degradativas de los nutrientes para obtener energía o para convertirlos en unidades. Figura N° 1.3. Figura N° 1.3. El catabolismo y anabolismo Fuente: (T. Audesirk y G.Audesirk, 2003). 1.2. CLASIFICACIÓN NUTRICIONAL BÁSICA Las bacterias son seres vivos y están compuestas al igual que las células eucariotas por proteínas, polisacáridos, lípidos, ácidos nucleicos, entre otros. Estas moléculas a su vez forman parte de estructuras celulares más complejas, como por ejemplo la pared celular y la membrana citoplasmática. Una característica de los seres vivos es la capacidad para sintetizar sus propios constituyentes a partir de nutrientes que toman del medio externo. 26 El crecimiento bacteriano se define como el aumento ordenado de todos los constituyentes químicos de la célula. Se trata de un proceso complejo, que supone la replicación de todas las estructuras y componentes celulares a partir de los nutrientes exógenos. El conocimiento de la fisiología y del metabolismo bacteriano tiene algunas aplicaciones prácticas: - Permite conocer el modo de vida y el hábitat de diferentes especies bacterianas. El ser humano actuando como hospedero por ejemplo, ofrece una variedad de nichos ecológicos que se diferencian entre sí por aspectos físicos y químicos (temperatura, concentración de O2, pH, presión osmótica, etc.) en los cuales pueden crecer y multiplicarse distintas especies bacterianas, de acuerdo a sus requerimientos nutricionales. - Permite formular medios de cultivo para el aislamiento e identificación de los patógenos participantes. - Desde el punto de vista terapéutico nos permite conocer y entender el modo de acción de algunos antimicrobianos que bloquean una vía metabólica o la síntesis de alguna macromolécula esencial para la bacteria. Tipos de metabolismo microbiano En el V Curso latinoamericano de Biotecnología (1996), señalaron que el crecimiento equilibrado de un microorganismo requiere de la integración de un gran número de rutas metabólicas (alrededor de 2000 procesos bioquímicos diferentes), interconectados, que participan en un correcto flujo de energía y carbón hacia la formación de más materia celular, los cuales deben operar coordenadamente y permitirle, al menor costo energético una adaptación a condiciones ambientales cambiantes (tanto químicas como físicas). Indicaron que en general la regulación y fisiología funciona a través de controles ejercidos sobre las enzimas claves de vías metabólicas y sobre los genes que codifican para enzimas o proteínas importantes en el comportamiento microbiano. 27 Los distintos tipos de metabolismo microbiano se pueden clasificar según tres criterios distintos: I. Según la fuente de carbono que utilizan El carbono es el mayor constituyente de la célula bacteriana, por lo tanto no llama la atención que requiera más carbono que cualquier otro nutriente. Las bacterias se pueden dividir de acuerdo a la forma en la que el organismo obtiene o utiliza el carbono para la construcción de la masa celular: Autótrofo. crecen sintetizando sus materiales a partir de sustancias inorgánicas sencillas. El carbono se obtiene del dióxido de carbono (CO2). Heterótrofo. su fuente de carbono es orgánica. El carbono se obtiene de compuestos orgánicos. En este último grupo se encuentran todas las bacterias de interés médico. Mixótrofo. son aquellas bacterias con metabolismo energético litotrofo (obtienen energía de compuestos inorgánicos), pero requieren sustancias orgánicas como nutrientes para su metabolismo biosintético. El carbono se obtiene tanto de compuestos orgánicos como fijando el dióxido de carbono. II. Según el punto de vista biosintético La forma en la que organismo obtiene los equivalentes reductores para la conservación de energía o en las reacciones biosintéticas: Litotrofo. son aquellas que sólo requieren sustancias inorgánicas sencillas Los equivalentes reductores se obtienen de compuestos inorgánicos. (SH2 SO, NH3, NO2-, Fe, etc.). Organotrofo. requieren compuestos orgánicos. Los equivalentes reductores se obtienen de compuestos orgánicos. (hidratos de carbono, hidrocarburos, lípidos, proteínas, alcoholes). III. Según la fuente de energía: 28 Según la forma en la que el organismo obtiene la energía para vivir y crecer: Quimiotrofo. La energía se obtiene de compuestos químicos externos. Fototrofo. La energía se obtiene de la luz. En la práctica, estos términos se combinan casi libremente (Figura N° 1.4, Figura N° 1.5 y Cuadro N° 1.1). Los ejemplos típicos son como sigue: Los quimiolitoautótrofos obtienen energía de la oxidación de compuestos inorgánicos y el carbono de la fijación del dióxido de carbono. Ejemplos: bacterias nitrificantes, bacterias oxidantes del azufre, bacterias oxidantes del hierro, bacterias oxidantes del hidrógeno. Los fotolitoautótrofos obtienen energía de la luz y el carbono de la fijación del dióxido de carbono, usando compuestos inorgánicos como equivalentes reductores. Ejemplos: Cianobacterias (agua como equivalente reductor), Chlorobiaceae, Chromaticaceae Chloroflexus (hidrógeno). 29 (sulfuro de hidrógeno), Los quimiolitoheterótrofos obtienen energía de la oxidación de compuestos inorgánicos, pero no pueden fijar el dióxido de carbono. Ejemplos: algunos Nitrobacter spp., Wolinella (con hidrógeno como equivalente reductor), algunas bacterias oxidantes del hidrógeno. Los quimioorganoheterótrofos obtienen energía, carbono y equivalentes reductores para las reacciones biosintéticas de compuestos orgánicos. Ejemplos: la mayoría de las bacterias, como Escherichia coli, Bacillus spp., Actinobacteria. Las bacterias patógenas que viven a expensas de la materia orgánica son quimioorganótrofas. Los fotoorganotrofos obtienen energía de la luz y el carbono y los equivalentes reductores para las reacciones biosintéticas de compuestos orgánicos. Algunas especies son terminantemente heterótrofas, pero muchas otras pueden también fijar el dióxido de carbono y son mixótrofas. Ejemplos: Rhodobacter, Rhodopseudomonas, Rhodospirillum, Rhodomicrobium, Rhodocyclus, Heliobacterium, Chloroflexus (alterna con fotolitoautotrofía con hidrógeno). Figura N° 1.4. Transformaciones biológicas de moléculas carbonadas Fuente: (Smith, C.A., y Wood, E.J., 1998). 30 Figura N° 1.5. Clasificación de nutricional de los organismos Fuente: (Smith, C.A., y Wood, E.J., 1998). Cuadro N° 1.1. Clasificación nutricional de los organismos. Tipo Fuente de energía Fuente de carbono Ejemplos Fotoautotrofas Luz CO2 Algas y cianobacterias Fotoheterotrofas Luz Compuestos orgánicos Algas y bacterias fotosintéticas Compuesto inorgánicos: H2, NH3, NO2, H2S, CO2 Pocas bacterias Compuesto orgánicos: glucosa La mayoría de bacterias Quimioautotrofas o Litotrofas Química Quimioheterotrofas o Heterótrofas Química Fuente: (Smith, C.A., y Wood, E.J., 1998). 31 1.3. SUMINISTRO ENERGÉTICO Las células bacterianas, poseen una gran variedad de sustancias como fuente de energía, ilimitada I. Las bacterias necesitan de un aporte energético para desarrollarse. II. El éxito evolutivo de las bacterias se debe en parte a su versatilidad metabólica. III. Todos los mecanismos posibles de obtención de materia y energía podemos encontrarlos en las bacterias. En los seres vivos, la utilización de la energía potencial contenida en los nutrientes se produce por reacciones de oxido-reducción. (Figura N° 1.6). Figura N° 1.6. Oxidación biológica Fuente: (A. Londoño C., 2002). Químicamente la oxidación está definida por la pérdida de electrones (e-) y la reducción por la ganancia de los mismos. En bioquímica, las reacciones de oxido- 32 reducción frecuentemente incluyen no sólo la transferencia de electrones sino de átomos enteros de hidrógeno, por lo que se conocen también con el nombre de reacciones de deshidrogenación. En reacciones de este tipo hay sustancias que ceden e- (dadoras) y otras que los aceptan (aceptoras). Figura N° 1.7. En las bacterias de interés médico los sistemas de oxido-reducción que transforman la energía química de los nutrientes en una forma biológicamente útil, incluyen la fermentación y la respiración. En la fermentación tanto la molécula dadora como la aceptora de electrones, son compuestos orgánicos, mientras que en la respiración hay un aceptor final exógeno, que cuando es el oxígeno hablamos de respiración aerobia, y cuando es un compuesto inorgánico hablamos de respiración anaerobia. Todas las formas de vida están basadas en prácticamente las mismas reacciones bioquímicas. Cada uno de los compuestos que se generan en este conjunto de reacciones se le denominan compuestos endógenos o metabolitos y al conjunto de todas las reacciones que suceden en una célula se le denomina metabolismo. Las bacterias y los animales superiores usan: Básicamente las mismas reacciones para producir la energía que necesitan para sostener los procesos vitales, Los mismos tipos de compuestos y mecanismos para construir sus macromoléculas y Los mismos conjuntos de reacciones para sintetizar los compuestos que intervienen en las diferentes reacciones bioquímicas. Se puede generalizar diciendo que todas las células tienen básicamente el mismo metabolismo, aunque obviamente hay diferencias entre ellas. 33 Figura N° 1.7. Visión General del Metabolismo. Se muestran las rutas metabólicas centrales y algunos de los intermediarios claves. Fuente: (C.K. Mathews, K.E. Van Holde y K.B. Ahern, 2002). Algunas células tienen mayor capacidad bioquímica que otras: Hay bacterias que sintetizan todos sus metabolitos a partir de compuestos inorgánicos y se les denomina autótrofos. Las células vegetales también pueden vivir a base de solo precursores inorgánicos. 34 Hay microorganismos que necesitan que en el medio de cultivo existan fuentes de carbono orgánico (azúcares) y se les denomina heterótrofos, Otros microorganismos necesitan que se les suministren además otros compuestos orgánicos que ellos no tienen la capacidad de sintetizar (a estos compuestos se les denomina factores de crecimiento) Las células de los animales necesitan un gran número de compuestos preformados los cuales deben estar en la dieta (se le denominan vitaminas, aminoácidos esenciales o ácidos grasos esenciales). En el proceso de diferenciación celular, durante la formación de un nuevo organismo, las distintas células que constituyen el embrión se especializan y sólo expresan parte de la información genética que contienen pasando a formar los distintos tejidos y órganos. El conjunto de reacciones que suceden en forma secuencial y que dan lugar a un compuesto o a una función integran un camino metabólico y se le da un nombre específico. Por ejemplo, 1) la glicólisis, es el camino metabólico por medio del cual se oxidan los azúcares produciendo piruvato y equivalentes reducidos NADH (Figura Nº 1.8); 2) la transformación de la acetil-coenzima A, proveniente de la descarboxilación del piruvato o de la beta-oxidación de los ácidos grasos, en anhídrido carbónico y equivalentes reducidos se le denomina ciclo de Krebs; 3) la transferencia de electrones de los equivalentes reducidos hasta el oxígeno molecular, acoplado con la síntesis de ATP, se le llama cadena de transporte de electrones o fosforilación oxidativa. Este último proceso está formado por un conjunto de enzimas complejas que catalizan varias reacciones de óxido-reducción, donde el oxígeno es el aceptor final de electrones. 35 Figura Nº 1.8. Metabolismo Intermediario. Fuente: (Ganong, W. F.,1996). 1.4. REGULACIÓN METABÓLICA Cada reacción metabólica está regulada no sólo con respecto a otras reacciones sino también con respecto a la concentración de nutrientes en el medio. La regulación se realiza a diferentes niveles: - Regulación de la actividad enzimática a través de: enzimas alostéricas, inhibición por retroalimentación, activación alostérica, y cooperatividad. - Regulación de la síntesis de enzimas por: inducción enzimática y represión por productos finales. 36 1.4.1. Mecanismos de Regulación enzimática En la célula operan dos mecanismos deferentes de regulación: la regulación de la síntesis enzimática y la regulación de la actividad enzimática. En ambos casos, actúan de mediadores componentes de bajo peso molecular, los cuales o bien son formados en la célula como metabolitos intermediarios, o bien entran en ella procedentes del medio. En ambos mecanismos reguladores actúan proteínas alostericas. Las proteínas alostericas son aquellas cuyas propiedades cambian si se les unen moléculas específicas denominadas efectores. Existen dos clases de proteínas alostericas: las enzimas alostericas cuya actividad se incrementa o se inhibe cuando se combinan con sus efectores, y las proteínas alostericas reguladoras que modulan la actividad de enzimas específicas. a. Regulación de la actividad enzimática Las proteínas alostericas estudiadas con mayor detalle han sido las enzimas alostericas, cuyo ejemplo lo constituye la aspartato transcarbamilasas (ATCasa), que cataliza la primera reacción de la biosíntesis de las pirimidinas y cuya actividad es inhibida por un producto final, la citidina trifosfato (CTP). concentraciones elevadas de CTP inhiben el funcionamiento de la ATCasa y, en consecuencia, la formación de más CTP hasta que su concentración disminuye a un nivel optimo. el ATP, un segundo efector de la ATCasa, activa la enzima, sirviendo para coordinar la síntesis de nucleótidos de purina y pirimidina. las enzimas alostericas son siempre proteínas de peso molecular relativamente elevado, compuestas por múltiples subunidades. por regla general estas subunidades son idénticas poseyendo cada una un lugar catalítico y otro alostérico. no obstante, la ATCasa se compone de dos clases diferentes de subunidades, una con función catalítica y otra con funciones reguladoras. En las bacterias anaerobias facultativas la fermentación (como única vía de generación de energía) es bloqueada en presencia de oxígeno, asegurando que el suministro de energía se produzca por la respiración, que consume menos glucosa y acumula menos lactato. En este fenómeno, conocido como efecto Pasteur, la 37 enzima fosfofructoquinasa es activada o inhibida según la relación ATP/ADP, regulando así el consumo de glucosa. Este es un ejemplo de regulación de la actividad enzimática por una enzima alostérica. El ejemplo clásico de regulación a nivel de la síntesis de enzimas lo constituye el operón lactosa. Hay 3 enzimas que participan en la utilización de la lactosa (ß-galactosidasa, galactósido permeasa y galactósido transacetilasa) que tienen un promotor único. En ausencia de lactosa, la transcripción para estas enzimas está bloqueada por acción de un represor que se une al promotor inhibiendo la acción de la ARN polimerasa. Cuando se agrega lactosa al medio, ésta se une al represor, bloqueando de este modo su unión al promotor, permitiendo así la acción de la ARN polimerasa y la síntesis de las tres enzimas. B. Regulación de la síntesis enzimática La inhibición por el producto final mediada por enzimas alostericas es en gran medida suficiente para asegurar que todas las rutas catabólicas y biosinteticas operen en equilibrio. Sin embargo, cuando no es necesario el producto de una ruta metabólica, las enzimas que catalizan las reacciones de las mismas resultan innecesarias. Muchas bacterias son capaces de utilizar un amplio espectro de compuestos orgánicos diferentes como fuentes de carbono y energía, pero en un momento dado, puede que solo uno de estos compuestos esté presente en el medio. Aunque la información genética necesaria para sintetizar las enzimas importantes se encuentra siempre presente, su expresión fenotípica está determinada por el medio, y una enzima determinada se sintetiza en respuesta a la presencia de su sustrato. 1.4.2. Crecimiento bacteriano Puede ser definido como el aumento ordenado de todos los constituyentes químicos de la célula. Las bacterias como grupo son extremadamente versátiles y tienen una capacidad enorme para utilizar una amplia gama de nutrientes que va desde compuestos inorgánicos simples a compuestos orgánicos más complejos. Los nutrientes se pueden dividir en dos clases: esenciales, sin los cuales la célula no puede crecer, y no esenciales que se utilizan cuando están presentes pero no 38 son indispensables. Algunos nutrientes son utilizados sólo como precursores de macromoléculas celulares y otros sólo como fuente de energía sin ser incorporados directamente al material celular, mientras que otros cumplen las dos funciones a la vez. También se pueden clasificar como macro y micronutrientes según la cantidad requerida. MACRONUTRIENTES La glucosa, por ejemplo, es utilizada como fuente de carbono y fuente de energía. También existen bacterias que pueden usar una variedad de otras sustancias orgánicas como fuente parcial o exclusiva de carbono. Entre las bacterias más versátiles se encuentra las del género Pseudomonas, muchas de las cuales pueden utilizar más de 100 compuestos orgánicos. Después del carbono el elemento más abundante en la célula es el nitrógeno, representa entre el 12 y el 15% del peso seco, y es el constituyente principal de proteínas y ácidos nucleicos. La mayoría de las bacterias son capaces de utilizar el amonio como fuente de nitrógeno, mientras que otras pueden usar los nitratos. La reducción de nitratos, se puede lograr por 2 mecanismos diferentes: 1. Reducción asimiladora: En la cual es reducido por la vía del nitrito, y 2. Reducción desasimiladora: Donde el nitrato sirve como aceptor final de electrones. La primera está bastante extendida entre las bacterias mientras que la segunda sólo es común en bacterias anaerobias y anaerobias facultativas. El fósforo es utilizado para la síntesis de ácidos nucleicos y fosfolípidos. La mayoría de las bacterias lo usan en forma inorgánica como PO4=. Los fosfatos orgánicos están ampliamente distribuidos en la naturaleza, pero para ser utilizados deben ser atacados primero por fosfatasas, enzimas que clivan estos compuestos liberando fósforo inorgánico. 39 Aunque requeridos en cantidades muy pequeñas los micronutrientes son importantes para la nutrición. Entre estos destacamos, cobalto, cobre y manganeso. 1.4.3. Factores de crecimiento Son sustancias que deben ser aportadas preformadas, ya que la bacteria que las requiere no las puede sintetizar a partir de los nutrientes más simples, por falla o ausencia de una vía metabólica. Estas sustancias incluyen vitaminas del complejo B, aminoácidos, purinas y pirimidinas. Las bacterias que no requieren factores de crecimiento de denominan prototróficas y las que los requieren, auxotróficas para ese factor. I. Oxígeno: Las exigencias de oxígeno de una bacteria en particular reflejan en parte el tipo de metabolismo productor de energía. De acuerdo a su relación con el oxígeno tenemos: - Anaerobios obligados: Hay de 2 tipos, estrictos y aerotolerantes, los primeros crecen en ausencia de O2 y este es sumamente tóxico, incluso letal cuando la exposición es breve. Los segundos también crecen sólo en ausencia de O2 pero toleran más que los anteriores su presencia; - Anaerobios facultativos: Son capaces de crecer en presencia o ausencia de oxígeno. - Aerobios obligados: Requieren oxígeno para su desarrollo. - Microaerófilos: Crecen mejor con tensiones de oxígeno bajas (3%-5%), las concentraciones elevadas (21%) tienen un efecto inhibidor para estas bacterias. En los aerobios, anaerobios facultativos y anaerobios aerotolerantes la enzima superoxidodismutasa impide la acumulación del radical superóxido; esta enzima está ausente en los anaerobios estrictos. El peróxido de hidrógeno formado por la acción de la superoxidodismutasa es destruido con rapidez por la enzima catalasa o por peroxidasas, como vimos más arriba. II. Dióxido de carbono: Algunas bacterias como Neisseria y Brucella tienen varias enzimas con baja afinidad por el CO2 y requieren una concentración más elevada 40 (10%) de la que habitualmente está presente en la atmósfera (0.03%). Estos requerimientos atmosféricos deben ser tenidos en cuenta cuando se realice el cultivo de estas bacterias. 1.4.4. Requerimientos físicos. POTENCIAL DE OXIDOREDUCCION: El potencial de óxido-reducción de un medio de cultivo es un factor crítico para determinar si se producirá o no el desarrollo de un inóculo sembrado en dicho medio. Para la mayor parte de los medios de cultivo en contacto con el aire, el potencial de óxido-reducción es de +0,2 a +0,4V a Ph 7. Los anaerobios obligados son incapaces de crecer a menos que el potencial sea tan bajo como -0,2 V como mínimo. Para establecer estas condiciones en un medio de cultivo se puede eliminar el oxígeno, recurriendo a sistemas de cultivo anaerobio o agregando al propio medio compuestos que contengan sulfidrilo como por ejemplo el tioglicolato de sodio. TEMPERATURA: Para cada bacteria existe una temperatura óptima de desarrollo y un rango en el cual este puede ocurrir. Las bacterias se dividen en tres grupos de acuerdo al rango de temperatura en el que pueden desarrollarse: - Psicrófilas: -5 a 30 ºC, óptimo:15ºC - Mesófilas: 10 a 45ºC, óptimo: 30ºC - Termófilas: 25 a 80ºC, óptimo: 55ºC. CONCENTRACION DE HIDROGENO (pH): También aquí existe un valor de pH óptimo dentro de un rango más amplio en el cual el crecimiento puede ocurrir. Para la mayor parte de las bacterias de interés médico, el pH óptimo es de 7,2 a 7,6. Sin embargo hay patógenos humanos como M. tuberculosus que resisten valores muy bajos de pH. CONDICIONES OSMOTICAS: La concentración de solutos con actividad osmótica dentro de la célula bacteriana es superior a la concentración exterior. Con excepción de los Micoplasmas y las formas lister (L), que no tienen pared celular, la mayor parte de las bacterias tienen una tolerancia osmótica importante lo que les permite soportar grandes cambios de la osmolaridad. 41 1.5. METABOLISMO HETERÓTROFO El metabolismo en los seres vivos es muy flexible y se puede ajustar a la cantidad y tipo de nutrientes disponibles en el medio que rodea a un microorganismo. Un ejemplo lo podemos obtener de las levaduras, las cuales se adaptan a las condiciones en que se les mantenga, pues poseen todo el conjunto de enzimas necesario para sintetizar sus elementos básicos como los aminoácidos y todos los demás componentes celulares, de tal manera que si crecen en un medio con glucosa como única fuente de energía, pueden sintetizar todas las moléculas que necesitan, aunque esto implica un gran gasto en energía. La levadura puede evitarlo si le proporcionamos los aminoácidos y nutrientes requeridos, ya que economiza el aparato enzimático responsable de la síntesis de todos los compuestos que necesita para vivir. Actualmente es posible tener en el laboratorio cultivos controlados de bacterias en los cuales el investigador modifica a voluntad la composición de los nutrientes. Esto ha permitido conocer y entender cómo los microorganismos modifican su metabolismo en función de la calidad nutritiva del medio en el cual se les hace crecer. Si, por ejemplo, la concentración de nutrientes ricos en nitrógeno baja, el microorganismo iniciará inmediatamente la síntesis de las enzimas necesarias para elaborar sus propios ácidos nucleicos y aminoácidos; todo está finalmente regulado y enfocado hacia la máxima economía metabólica y, en última instancia, al ahorro de energía. 1.5.1. Características metabólicas El metabolismo microbiano es el conjunto de procesos por los cuales un microorganismo obtiene la energía y los nutrientes (carbono, por ejemplo) que necesita para vivir y reproducirse. Los microorganismos utilizan numerosos tipos de estrategias metabólicas distintas y las especies pueden a menudo distinguirse en base a estas estrategias. Las características metabólicas específicas de un microorganismo constituyen el principal criterio para determinar su papel ecológico, su responsabilidad en los ciclos biogeoquímicos y su utilidad en los procesos industriales. 42 - Las bacterias tienden versatilidad a intercalar fácilmente los nutrientes y productos metabólicos terminales - Esto les permite desarrollar procesos metabólicos con gran rapidez El metabolismo de la célula comprende dos grandes tipos de reacciones: La mayoría de los microorganismos son heterótrofos (o más exactamente quimiorganoheterótrofos), con compuestos orgánicos como fuentes de carbono y de energía (Figura Nº 1.9). Los microorganismos heterótrofos viven de los alimentos que roban a anfitriones vivos (como comensales o parásitos) o de la materia orgánica muerta de todo tipo (saprófagos). Este metabolismo microbiano constituye el principal factor de descomposición de todos los organismos después de muerte. Muchos microorganismos eucariontes son heterótrofos por depredación o parasitismo, características también encontradas en algunas bacterias tales como Bdellovibrio (un parásito intracelular de otras bacterias, causando la muerte de sus víctimas) y algunas Myxobacteria tales como Myxococcus (depredadora de otras bacterias a las que mata y succiona mediante la cooperación de enjambres de numerosas células). La mayoría de las bacterias patógenas son parásitos heterótrofos de seres humanos o de otras especies eucariontes. Los microorganismos heterótrofos son extremadamente abundantes en naturaleza y responsables de la degradación de los polímeros orgánicos tales como celulosa, quitina o lignina que son generalmente indigeribles para los animales más grandes. Esta degradación, generalmente, requiere la colaboración de varios organismos distintos, cada uno de los cuales realiza uno de los pasos de la degradación hasta obtener dióxido de carbono. Hay muchas variaciones en este tema, pues diversos organismos pueden degradar diversos polímeros y secretar diversos residuos. Algunos organismos pueden incluso degradar los compuestos más recalcitrantes tales como pesticidas y petróleo, realizando su reciclado. 43 Figura Nº 1.9. Metabolismo Intermediario. Fuente: (J. Koolman y K–H Röhm, 2004). 44 Bioquímicamente, el metabolismo heterótrofo procarionte es mucho más versátil que el de los organismos eucariontes, aunque muchos procariontes comparten los modelos metabólicos más básicos con los eucariontes, por ejemplo, usando la glicolisis (también llamada ruta EMP) para el metabolismo del azúcar y el ciclo del ácido cítrico en la degradación del acetato, produciendo energía bajo la forma de ATP y reduciendo energía bajo la forma de NADH o quinona. Estas rutas metabólicas básicas están muy extendidas porque también están implicadas en la biosíntesis de muchos componentes necesarios para el crecimiento de la célula (a veces en la dirección contraria). Sin embargo, muchas bacterias y archaea utilizan rutas metabólicas alternativas con la excepción de la glicólisis y del ciclo del ácido cítrico. Un ejemplo bien estudiado es el metabolismo del azúcar por la ruta del ceto-desoxi-fosfogluconato (también llamada ruta ED) en Pseudomonas en vez de la ruta glicolítica. Por otra parte, hay incluso una tercera ruta alternativa catabólica del azúcar usado por algunas bacterias, la ruta de la pentosa fosfato. Los elementos constitutivos o macromoléculas tienen su génesis en unos pocos precursores denominados metabolitos focales: glucosa 6-fosfato, fosfoenolpiruvaro, oxalacetato y l cetoglutarato, estos se interrelacionan y originan compuestos intermediarios: fosfatos de azucares, piruvato, acetil CoA, aspartato, glutamato, etc., y productos terminales como: aminoácidos, bases pirimidinicas, polisacáridos, lípidos, entre otros. (Figura Nº 1.10). Figura Nº 1.10. Familias biosinteticas de aminoácidos 45 * En Bacterias, ** en Hongos Fuente: (Pares I.F. y A. Juárez, 1997). Esta diversidad y capacidad metabólicas de los procariontes que les permite utilizar una variedad enorme de compuestos orgánicos se debe a una historia y diversidad evolutivas mucho más profundas que las de los eucariontes. Es también significativo que las mitocondrias se originaron en los eucariontes por endosimbiosis de una bacteria relacionada con los parásitos intracelulares Rickettsia, y también los simbiontes de las plantas Rhizobium o Agrobacterium. Por lo tanto, no es sorprendente que todas las mitocondrias eucariotas compartan características metabólicas con estas Proteobacteria. La mayoría de los microorganismos respiran (usando una cadena de transporte de electrones), aunque el oxígeno no es el único receptor terminal de electrones que puede usarse. El uso de receptores terminales de electrones distintos al oxígeno tiene consecuencias biogeoquímicas importantes. (Cuadro Nº 1.2 1.11.). 46 y Figura Nº Cuadro Nº 1.2. Patrones de reacciones de oxido reducción suministradoras de energía utilizadas por las bacterias. Donador de H2 Sustancias inorgánicas Sustancias orgánicas O2 (Respiración anaerobia) I. Respiración aerobia de sustancias inorgánicas Ej.: (Nitrosomonas) NH3 NO2= O2 H2O II. Respiración aerobia de sustancias orgánicas. Ej.: (muchos organismos) Glucosa CO2 O2 H2O Aceptor de H2 NO3=, SO4=, fumarato (Respiración anaerobia) Compuestos orgánicos (Fermentación) III. Respiración anaerobia de sustancias inorgánicas. Ej.: (Thiobacillus denitrificans) S SO4= NO3= N2 IV. Respiración anaerobia de sustancias orgánicas. Ej. (Desulfovibrio) Ácido láctico CO2 SO4= H2S (Ninguno) V.Fermentación de sustancias orgánicas Ej.: (Streptococcus) -4 Glucosa 2-piruvato +4H 2 ácido láctico FUENTE: (Brooks. G. F., J. S. Butel y S. A. Morse, 2005) 47 Figura Nº 1.11. Alternativas del metabolismo microbiano. Fuente: (M.T. Madigan, J.M. Martinko y J. Parker., 2002). 1.5.2. Mecanismos de generación de ATP La conservación intracelular de energía ocurre principalmente por medio de la síntesis de ATP. Los métodos usados por las bacterias para generar ATP son principalmente: Fosforilación a nivel de sustrato Fosforilación oxidativa Fotofosforilación (durante la fotosíntesis). 48 La fosforilación a nivel de sustrato es un sistema usado por ciertas bacterias quimiorganotrofas. el sustrato orgánico (donador de electrones) pasa por una ruta catabólica (p.ej., la ruta glucolítica), y uno de los intermediarios de esa ruta es oxidado por un coenzima (p. ej., NAD+), de manera que se origina un intermediario no fosforilado con una gran energía de hidrólisis. Dicho intermediario experimenta enseguida una sustitución con un fosfato, para dar la correspondiente forma acilfosfato (siendo este enlace de alta energía). Finalmente, este acil-fosfato dona su fosfato de alta energía al ADP, que pasa a ATP. Ejemplo: gliceraldehido-3-p à 1,3difosfoglicérico à 3-fosfoglicérico. La fosforilación oxidativa es el proceso por el que se forma ATP como resultado de la transferencia de electrones desde el NADH o del FADH2 al O2 a través de una serie de transportadores de electrones. (Figura Nº 1.12). Figura Nº 1.12. Flujo de electrones durante la fosforilacion oxidativa Fuente: (Stryer, 1995). 49 El flujo de electrones desde el NADH o el FADH2 al O2 a través de complejos proteicos localizados en la membrana interna mitocondrial, provoca el bombeo de protones hacia el exterior de la matriz mitocondrial. Se genera una fuerza protomotriz que está formada por un gradiente de pH y por un potencial eléctrico transmembranal. Cuando los protones regresan a la matriz mitocondrial a través de un complejo enzimático, se sintetiza ATP (fig. 1). De esta forma, la oxidación y la fosforilación están acopladas por un gradiente de protones a través de la membrana interna mitocondrial. (Figura Nº 1.13). Figura Nº 1.13. Fosforilación a nivel de sustrato y Fosforilación oxidativa. Fuente: (J. Koolman y K–H Röhm, 2004). 50 1.5.3. La fermentación La fosforilacion a nivel de sustrato es el único modelo posible de formación de ATP como resultado de una fermentación, la cual puede ser realizada por los siguientes grupos de microorganismos: 1. Anaerobios estrictos. 2. Facultativos. 3. Anaerobios aerotolerantes. En los dos primeros, la presencia de oxigeno modifica el metabolismo generador de ATP; en el tercero esto no ocurre. La fermentación es un tipo específico de metabolismo heterótrofo que utiliza carbono orgánico en vez de oxígeno como receptor terminal de electrones. Esto significa que estos organismos no utilizan una cadena de transporte de electrones para oxidar NADH a NAD+ y por lo tanto deben tener un método alternativo para usar esta energía reductora y mantener una fuente de NAD+ para el funcionamiento apropiado de las rutas metabólicas normales, como por ejemplo, la glicólisis (Figura Nº 1.14). Puesto que no requieren oxígeno, los organismos fermentantes son anaerobios. Muchos organismos pueden utilizar fermentación bajo ciertas condiciones anaerobias y respiración cuando el oxígeno está presente. Figura Nº 1.14. Destinos metabólicos del piruvato. Fuente: (T. Audesirk y G.Audesirk, 2003). 51 Estos organismos son anaerobios facultativos. Para evitar la superproducción de NADH, los organismos fermentantes obligados generalmente no tienen un ciclo completo del ácido cítrico. En vez de usar ATPasas como en la respiración, el ATP en organismos fermentantes es producido por la fosforilación a nivel de substrato donde un grupo fosfato se transfiere de un compuesto orgánico de gran energía al ADP para formar el ATP. (Figura Nº 1.15). Figura Nº 1.15. Modelo propuesto para el estado energizado de la membrana Fuente: (Pares I.F. y A. Juárez, 1997. Como resultado de la necesidad de producir compuestos orgánicos con fosfato de la alta energía (generalmente bajo la forma de CoA-ésteres) los organismos fermentantes utilizan NADH y otros cofactores para producir una gran variedad de subproductos metabólicos reducidos, a menudo incluyendo hidrógeno. Estos compuestos orgánicos reducidos son generalmente ácidos orgánicos cortos y alcoholes derivados del piruvato, el producto final de la glicólisis. Ejemplos incluyen el etanol, acetato, lactato y el butirato (Figura Nº 1.16). Los organismos fermentantes son muy importantes industrialmente y se utilizan parar elaborar muchos tipos de productos alimenticios. Los productos finales 52 metabólicos producidos por cada especie bacteriana específica son los responsables del gusto y características de cada alimento. Figura Nº 1.16. Estrategias microbianas de descarboxilación del piruvato. 1, piruvato descarboxilasa. Funcional en levaduras y algunas bacterias, como Zymononas mobilis cuando fermentan la glucosa. 2, piruvato deshidrogenasa. Característico del desarrollo aerobio de, entre otras, las bacterias entéricas. 3, piruvato-formiato-liasa. Lo sintetizan las bacterias entéricas en condiciones anaeróbicas. 4, piruvato-ferredoxina-oxidorreductasa. Característico de los clostridios. 5, parte del complejo de la α-acetolactato sintasa de algunas bacterias entéricas (el α-acetolactato se genera por condensación del acetaldehído activo con piruvato). Un sistema equivalente es utilizado por las bacterias lácticas para producir diacetilo (en este caso, el acetaldehído activo condensa con acetil-CoA) Fuente: (Pares I.F. y A. Juárez, 1997). No todos los organismos fermentantes usan fosforilación a nivel de substrato. En su lugar, algunos organismos son capaces de acoplar directamente la oxidación de compuestos orgánicos de poca energía con la formación de un gradiente 53 electroquímico para mover un protón (o sodio) y así realizar la síntesis de ATP. Ejemplos de estas formas inusuales de fermentación incluyen la fermentación del succinato por el Propionigenium modestum y la fermentación del oxalato por Oxalobacter formigenes. Estas reacciones son de rendimiento energético extremadamente bajo. Los seres humanos y otros animales también utilizan la fermentación para consumir el exceso de NADH produciendo lactato, aunque ésta no es la forma principal de metabolismo como en los microorganismos fermentantes. 54