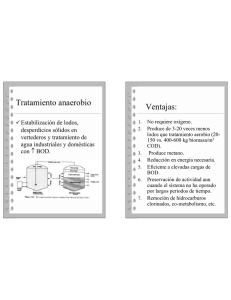
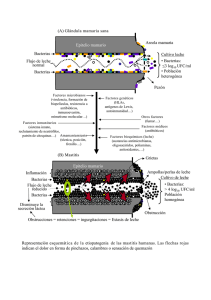
Centre de Recursos Docents de la Facultat de Medicina UAB
Anuncio

LAS BACTERIAS 1. Estructura de la célula bacteriana 2. Antígenos bacterianos 3. Formas de resistencia: esporas 4. Morfología y visualización de las bacterias 5. Metabolismo bacteriano 6. División bacteriana 7. Medios de cultivo 8. Taxonomía bacteriana 9. Identificación de bacterias 10. Bacterias de interés en medicina 1. Estructura de la célula bacteriana Las bacterias son células procariotas, de un tamaño entre 0’2 y 5 µm, envueltas por una membrana citoplasmática y una pared externa de estructura característica; poseen el citoplasma repleto de ribosomas y el material genético está libre sin membrana nuclear. Algunas son móviles por poseer flagelos (Figura 1). Citoplasma. El citoplasma de las células procariotas, a diferencia del de las eucariotas, carece de orgánulos membranosos internos (mitocondrias, retículo endoplasmático, aparato de Golgi) y de los microtúbulos que forman el citoesqueleto de las células eucariotas. Está constituido por una masa densa, amorfa, con aspecto granular por estar repleto de abundantes ribosomas. Los ribosomas de las células procariotas son más pequeños que los de las eucariotas, aunque de estructura semejante a los de éstas, en ellos tiene lugar la síntesis proteica. Genoma bacteriano. El genoma bacteriano está formado por una sola molécula de DNA bicaternario de forma circular, cerrada, y gran tamaño (“cromosoma”). En Escherichia coli mide 1-2 mm lo que supone unas mil veces la longitud de la bacteria. Este gran tamaño y la ausencia de histonas1 hace que el DNA se encuentre aparentemente apelotonado formando un ovillo en el interior del citoplasma. Esta molécula de DNA no se encuentra separada del citoplasma por una membrana nuclear sino que está directamente inmersa en el mismo, por lo que no se puede hablar con propiedad de núcleo bacteriano, refiriéndose a este acumulo de material genético como nucleoide. La ausencia de membrana nuclear permite una trascripción muy rápida de la información genética. Las bacterias pueden albergar material genético adicional al cromosoma en forma de pequeñas moléculas circulares de DNA bicaternario libre en el citoplasma, denominadas plásmidos, que constituyen material genético extracromosómico autónomo. Los plásmidos, como los fagos lisogénicos2 pueden codificar diversos 1 Aunque existen otras proteínas asociadas al DNA bacteriano con función de soporte 2 Ver lección de virus. Los bacteriófagos, aunque constituyen un fragmento de material genético exógeno, en estado lisogénico tienden a integrarse en el cromosoma bacteriano. También los plásmidos pueden circunstancialmente integrarse en el cromosoma bacteriano 1 productos de interés, como enzimas inactivantes de antibióticos, bacteriocinas3, exotoxinas, adhesinas y otros factores de patogenicidad, así como enzimas con actividades metabólicas. Los plásmidos se replican de forma sincrónica con el cromosoma celular y como él se transmiten de forma estable a las células hijas (transmisión vertical); este tipo de transmisión también se da en otros episomas como los fagos lisogénicos. Algunos plásmidos, conocidos como plásmidos transferibles, pueden transmitirse a otras células bacterianas carentes de ellos mediante un mecanismo de transferencia horizontal denominado conjugación, que comporta su replicación previa, y el paso de una copia a través de una fimbria a otra bacteria, por lo que ambas células, donadora y receptora, poseen una copia del mismo. Membrana celular. Posee las características generales de las membranas biológicas. Está formada por una doble capa de fosfolípidos en la que se encuentran incluidas proteínas de gran importancia funcional relacionadas con la permeabilidad y el metabolismo (Figuras 2 y 3). La membrana celular aísla al citoplasma del medio externo, en ella se localizan secuencialmente diversos enzimas implicados en el metabolismo energético y biosintético. También participa en el anclaje y segregación del “cromosoma” bacteriano durante la división celular mediante invaginaciones (mesosomas). Constituye el soporte para los flagelos y también aporta la energía necesaria para su movimiento. Pared bacteriana. La pared bacteriana es una estructura rígida, que se encuentra situada por fuera de la membrana citoplasmática constituyendo un exoesqueleto, cuya composición química fundamental es una molécula, conocida como peptidoglicano, que sólo se encuentra en el mundo bacteriano. El peptidoglicano es un largo polímero, filamentoso, constituido por dos aminoazúcares (N-acetil-glucosamina y N-acetil-murámico) alternados. Estos filamentos poliméricos se enlazan entre sí por pequeños péptidos. En efecto, del Nacetil-murámico se originan unos tetrapéptidos los cuales se unen entre sí por otros puentes peptídicos. 3 Las bacteriocinas son proteínas con actividad antibacteriana para otras bacterias diferentes a las que las producen, que les permite competir en territorios donde existe una flora mixta 2 Podría imaginarse la membrana celular como el frágil vidrio de una garrafa protegido por su envoltura de mimbre, que equivaldría a la pared. Los largos “mimbres” formados por los polímeros de los azúcares estarían unidos entre sí por pequeños péptidos para no desarmarse. Según las características generales de la pared, las bacterias pueden dividirse en dos grandes grupos, que se diferencian fácilmente entre sí al observarlas al microscopio óptico después de haber sido teñidas por el método de Gram. En la tinción de Gram se utiliza como colorante el violeta de genciana que tiñe las células bacterianas de color violeta oscuro. Después de esta primera tinción, se decolora la preparación con alcohol y acetona. Unas bacterias, las grampositivas, resisten la decoloración y persisten teñidas de violeta intenso, mientras que otras bacterias, las gramnegativas, se decoloran. Para poder observar estas bacterias decoloradas, se utiliza un colorante como la safranina que las tiñe de color rosa pálido, contrastando claramente con el violeta. Este comportamiento distinto depende de diferencias en la estructura de la pared (Figura 4). Pared de las bacterias grampositivas. En este grupo de bacterias el peptidoglicano es muy grueso, constituyendo una ancha malla tridimensional a través de la cual emergen radialmente al exterior estructuras fibrilares polisacáridas, entre otras los ácidos teicoicos. También existen proteínas fibrilares que emergen desde el peptidoglicano a la superficie4. Algunas de estas estructuras constituyen los antígenos de superficie de las bacterias grampositivas (Figuras 5 y 6). Pared de las bacterias gramnegativas. El peptidoglicano forma una malla fina, existiendo por fuera del mismo y unida a él por lipoproteínas, una membrana externa, que no se encuentra en las bacterias grampositivas. Entre la membrana externa y la membrana citoplasmática se delimita un espacio virtual, denominado periplásmico (Figuras 5 y 7). La membrana externa tiene estructura clásica en doble capa fosfolipídica y contiene en gran abundancia proteínas algunas de las cuales regulan la permeabilidad (porinas) y lipopolisacáridos (LPS). El lipopolisacárido es una molécula alargada que posee dos fragmentos, uno interno de naturaleza lipídica, anclado en la membrana, que se conoce como lípido A y otro que sobresale al exterior, formado a su vez por dos partes: una medial denominada 4 En algunos grupos bacterianos emergen lípidos 3 “core” y otra distal o externa formada por una secuencia de 2 a 3 azúcares que se repiten y constituyen el antígeno de superficie más importante de las bacterias gramnegativas denominado antígeno O (Figura 8). Algunas bacterias, poseen una pared rica en ácidos grasos de cadena larga (ácidos micólicos y ceras) junto a proteínas y polisacáridos. Estos ácidos grasos les confieren propiedades particulares; entre ellas una mayor resistencia a agentes químicos, como los ácidos y álcalis5. Se tiñen mal por el método de Gram y lenta pero intensamente con colorantes como la fucsina (rojo), pero una vez teñidas por este colorante a diferencia del resto de las bacterias, resisten la decoloración por ácidos y alcoholes denominándose bacterias alcohol-ácido resistentes. Esta característica tintorial se evidencia mediante la tinción de Ziehl-Neelsen en las que las bacterias alcohol-ácido resistentes se tiñen de rojo y las que pierden la coloración al ser tratadas por alcohol y ácido se tiñen con azul de metileno para contrastarlas con las anteriores. Sólo las micobacterias, nocardias y rodococos entre las bacterias de interés en medicina, y las esporas bacterianas, son alcohol-ácido resistentes, aunque existen otros grupos como las corinebacterias que no son alcohol-ácido resistentes, pero también poseen una pared rica en ácidos grasos. Ciertas bacterias con estructura de gramnegativas como las espiroquetas, poseen una pared caracterizada por presentar flagelos en el espacio periplásmico que les confiere una morfología espirilar característica y colaboran a su movilidad. Por último, las arqueobacterias y los micoplasmas carecen de pared bacteriana. Estas bacterias sin pared son de forma variable. El peptidoglicano constituye un exoesqueleto rígido. Su función principal es evitar la ruptura osmótica de la membrana ya que en condiciones normales la presión intracelular en las células procariotas es más elevada que la extracelular. Secundariamente esta pared rígida confiere la forma característica de cada bacteria. Según la morfología pueden distinguirse cuatro tipos de bacterias: cocos, de forma esférica; bacilos, de forma alargada; espiroquetas, de forma espiral y bacterias sin pared de forma irregular. 5 Aspecto muy importante, en relación a su capacidad de persistencia en el medio ambiente y a las técnicas para su cultivo (véase más adelante) 4 La inoculación a animales de experimentación de lipopolisacárido da lugar a una compleja y grave reacción tóxica que puede abocar al shock6, por lo que se le ha denominado endotoxina. Esta toxicidad, a la que también es susceptible el hombre, depende probablemente del lípido A. La cadena distal polisacárida del antígeno O protege a la bacteria frente a diversos sistemas bactericidas existentes en los tejidos, en particular el complemento. Se halla especialmente desarrollada en las bacterias gramnegativas que carecen de cápsula. La membrana externa constituye una barrera que condiciona el transporte de la mayoría de las sustancias químicas. La permeabilidad en las bacterias gramnegativas se efectúa a este nivel mediante porinas y proteínas de transporte específico. Este fenómeno de permeabilidad tiene importancia en microbiología clínica ya que es responsable de importantes diferencias en la sensibilidad a los antibióticos. Así, las bacterias gramnegativas, son resistentes a una serie de antibióticos hidrofílicos, a los que son sensibles las bacterias grampositivas, ya que no pueden atravesar sus membranas externas en función de su tamaño, configuración espacial de la molécula o carga neta. Por otra parte, en el espacio periplásmico se hallan libres diversas enzimas que aún siendo externas al protoplasma celular no difunden libremente al medio por lo que su eficacia resulta incrementada por concentración en este espacio. Las betalactamasas, enzimas que inactivan los antibióticos betalactámicos (penicilina y otros), se localizan a este nivel. Exopolímeros bacterianos: Cápsula y glicocalix. Son estructuras fibrilares poliméricas localizadas externamente a la pared, generalmente de naturaleza polisacárida que pueden ser productos de síntesis celular exportados (cápsula) o productos sintetizados extracelularmente por exoenzimas bacterianas a partir de sustratos extracelulares (glicocalix). La cápsula posee una notable capacidad antigénica y su actividad antifagocitaria la convierte en un importante factor de virulencia para las bacterias que la poseen (Figura 9). El glicocalix envuelve a las células bacterianas y facilita su adhesividad a diversas estructuras, como en Streptococcus mutans –agente causal de la caries dental– al 6 Hipotensión por vasodilatación, seguida de mala perfusión tisular y fallo multiorgánico 5 esmalte de los dientes. Retiene humedad y nutrientes y puede dificultar el acceso de los antibióticos y los anticuerpos a los antígenos de superficie. Muchas bacterias no poseen estos exopolímeros que son estructuras facultativas y por tanto no son imprescindibles para la vida bacteriana. Flagelos. Son estructuras fibrilares de naturaleza proteica que constituyen los órganos de locomoción de las bacterias. Están formados por la agregación de múltiples subunidades de una proteína globular denominada flagelina. La agregación de estas subunidades da lugar a un largo cilindro hueco (Figuras 10 y 11). Su naturaleza proteica confiere al flagelo un carácter muy inmunogénico (antígenos H). Pili y Fimbrias. Son estructuras externas filamentosas, tubulares, huecas formadas de modo semejante a los flagelos por agregación de múltiples subunidades proteicas. Poseen capacidad antigénica. Algunas fimbrias juegan un papel importante en la patogénia de las infecciones bacterianas ya que permiten a las bacterias que las poseen adherirse a las superficies mucosas, otras poseen funciones semejantes a la cápsula y otras participan en los procesos de transferencia de plásmidos por conjugación entre las células (pili sexuales). 2. Antígenos bacterianos Las estructuras con capacidad antigénica en las bacterias son muy numerosas. Sin embargo las que posen mayor interés en medicina, por las razones que se señalarán más adelante, son de localización periférica como la cápsula (K), el lipopolisacárido (O) y las proteínas de la membrana externa en las bacterias gramnegativas, y los ácidos teicoicos y otros exopolímeros de la pared en las grampositivas. Los flagelos (H), las fimbrias y las proteínas de la membrana citoplasmática también poseen carácter antigénico. Los anticuerpos protectores van dirigidos generalmente a los antígenos capsulares. Existe evidencia de este hecho ya que se han utilizado purificados como vacunas eficaces en el caso de neumococo, hemófilos y del agente causal de la fiebre tifoidea (antígeno capsular Vi de Salmonella typhi). 6 Se conoce menos la importancia protectora de los anticuerpos dirigidos frente a los antígenos O, H y proteínas de membrana externa y el papel que juegan estas estructuras en la patogenicidad de algunas bacterias. Todos estos antígenos se han utilizado, dependiendo de los microorganismos, para la subdivisión de las especies bacterianas en serogrupos o serotipos. Así el antígeno capsular permite subdividir al neumococo, al meningococo y a los hemófilos en diversos tipos. Los antígenos O y H permiten subdividir las especies de enterobacterias en serotipos y los antígenos de membrana externa permiten subdividir los grupos de meningococo en diversos subtipos. 3. Formas de resistencia: Esporas Algunas bacterias grampositivas , en situaciones ambientales adversas, son capaces de generar formas de resistencia, denominadas esporas7 mediante un proceso complejo de esporulación. Las esporas constituyen una de las escasas formas de diferenciación celular aparecidas en el mundo bacteriano8. Una espora bacteriana posee una forma esférica u oval y contiene una copia completa del material genético de la bacteria, estructuras para la síntesis proteica, parte de los sistemas enzimáticos para la obtención de energía y sustancias de reserva energética, rodeados por varias envolturas concéntricas, de diferente composición química, algunas de las cuales son muy resistentes a los agentes físicos y químicos. Al encontrar condiciones favorables las esporas revierten al estado morfológico y metabólico normal –vegetativo– a través de un proceso de germinación. Las esporas, que pueden ser circulares u ovales, según su localización en la bacteria antes de liberarse pueden clasificarse en centrales, subterminales y terminales. Estos datos poseen valor para la identificación de las bacterias esporuladas. Las esporas únicamente se producen en algunas bacterias grampositivas y entre ellas tan sólo existen dos géneros, Bacillus y Clostridium, de interés en medicina. 7 Para destruir las esporas se requiere calor húmedo a 121°C durante 20 minutos, calor seco a 180°C durante 2 horas o antisépticos enérgicos 8 Otras son las variaciones morfológicas que se producen en el ciclo vital de las clamidias, coxiela y las estructuras formadas por las mixobacterias 7 4. Morfología y visualización de las bacterias La mayoría de las bacterias poseen un tamaño variable entre 0.2 y 5 µm pudiéndose observar al microscopio óptico en fresco o teñidas mediante la tinción de Gram u otras como la de Ziehl-Neelsen. Aunque presentan escasa variabilidad morfológica, pueden adscribirse a varios grupos según su forma. Las redondeadas se denominan cocos y las alargadas bacilos. Los bacilos con forma de coma, se denominan vibriones. Algunos bacilos poseen forma espirilar denominándose espirilos, que no hay que confundir con las espiroquetas que presentan una morfología aparentemente semejante, pero poseen flagelos en el espacio periplásmico. Las bacterias sin pared, como los micoplasmas, poseen forma irregular. Cada bacteria perteneciente a uno de estos tipos morfológicos (cocos y bacilos) puede subdividirse según la tinción de Gram en grampositivas o gramnegativas y según la de Ziehl en alcohol-ácido resistentes o no. La morfología, la coloración por la tinción de Gram y la de Ziehl-Neelsen, así como la presencia o ausencia de esporas, que pueden visualizarse como una estructura morfológicamente definida, constituyen los caracteres más elementales para la clasificación de las bacterias. El examen microscópico es muy utilizado para detectar e identificar presuntivamente las bacterias en las diversas muestras clínicas que se remiten al laboratorio para estudio. Sin embargo, algunas bacterias como los micoplasmas, las espiroquetas, clamidias y rickettsias, no se visualizan mediante las técnicas microscópicas convencionales, por requerir tinciones especiales o por su pequeño tamaño. 5. Metabolismo bacteriano El metabolismo es el conjunto de reacciones bioquímicas catabólicas que producen energía para el movimiento, el transporte de sustancias en la célula y para la síntesis de los compuestos celulares estructurales a través de las reacciones anabólicas. Aunque las bacterias poseen todas las potencialidades metabólicas, autótrofas y heterótrofas conocidas, sólo poseen interés en medicina las quimiorgano- 8 heterótrofas, que utilizan compuestos orgánicos para la obtención de energía, como las existentes en los tejidos y secreciones del cuerpo humano. Las bacterias quimiorganotrofas obtienen energía de los hidratos de carbono y otros sustratos orgánicos que se incorporan a las rutas metabólicas fermentativas. La energía se acumula en forma de ATP, que se produce a partir del ADP (ADP+P=ATP), por dos mecanismos, el de fosfosilización a nivel de sustrato o la fuerza motriz de protones (por gradiente electroquímico membrana). El primero tiene lugar en las fermentaciones y el segundo en la cadena respiratoria. La fermentación es un proceso anaerobio en el que, a través de diversas vías metabólicas –siendo la de Embden-Meyerhof una de las más universales– se produce ácido pirúvico a partir de la glucosa y se obtiene una ganancia neta de 2 ATP y 2 NADPH, fuentes de energía y de poder reductor, respectivamente (Figura 12). El ácido pirúvico (piruvato) puede ser posteriormente degradado, por fermentación, a varios catabolitos como ácido láctico, etanol, butilenglicol, ácido butírico, ácido propiónico y otros (Figura 13). Del ácido pirúvico, formado por fermentación puede aún extraerse gran cantidad de energía mediante un proceso de respiración aerobia. A partir del pirúvico se forma acetil coenzima A que se incorpora al ciclo de Krebs. A él están acopladas las cadenas respiratorias que permiten la síntesis de ATP. En las cadenas respiratorias el aceptor final de electrones es el oxígeno (Figura 14). Muchas bacterias heterótrofas pueden obtener energía tanto por vía fermentativa (anaerobia) como por la respiración (aerobia) por lo que pueden cultivarse y crecer tanto en ausencia como en presencia de oxígeno, denominándose aerobias y anaerobias facultativas. Algunas, denominadas aerobias estrictas, sólo pueden crecer en presencia de oxígeno porqué las vías fermentativas no producen suficiente energía. Otras sólo pueden obtener energía por vía fermentativa anaerobia y además la presencia de oxígeno es letal para ellas. Estas bacterias denominadas anaerobias estrictas deben cultivarse en una atmósfera carente de oxígeno. Por último, algunas bacterias denominadas microaerófilas activan su metabolismo a 9 concentraciones bajas de oxígeno, pero no crecen a la concentración atmosférica y lo hacen muy lentamente en anaerobiosis estricta. Las subunidades moleculares necesarias para la biosíntesis de macromoléculas, como los aminoácidos, azúcares, lípidos, bases púricas, pirimidínicas, así como ciertas vitaminas y otros factores esenciales, pueden ser tomadas del medio y utilizadas como tales para la síntesis. Sin embargo, la mayoría de estas subunidades pueden ser sintetizadas a partir de catabolitos intermediarios formados en los procesos catabólicos de moléculas sencillas y universales como los glúcidos y otras. La glucosa 6-fosfato, el fosfoenol piruvato, el oxalacetato, y el α-cetoglutarato, constituyen “metabolitos focales” a partir de los cuales, formando diversos intermediarios, pueden dar lugar a la mayoría de las moléculas estructurales o funcionales complejas de las células bacterianas, de modo que la combustión de los azúcares además de aportar energía va formando catabolitos químicos que son precursores de subunidades básicas estructurales. Ello muestra como existe una profunda interdependencia entre los procesos catabólicos bioenergéticos y los biosintéticos. Aspectos aplicados del estudio de metabolismo bacteriano. Aunque el estudio del metabolismo bacteriano posee gran interés desde numerosos puntos de vista (teórico9, farmacológico, industrial) desde el punto de vista de la microbiología médica aplicada posee valor taxonómico ya que las bacterias, tras su clasificación inicial según la morfología, Gram, Ziehl, movilidad y esporulación, se diferencian en grupos según el tipo respiratorio: aerobias, anaerobias, facultativas o microaerófilas y posteriormente pueden clasificarse a nivel de familia, género o especie según posean o no diversas capacidades metabólicas. El conocimiento del metabolismo bacteriano permite además comprender el fundamento de los métodos de cultivo, el modo de acción de algunos antibióticos que actúan interfiriendo rutas metabólicas específicas y otros aspectos de la biología relacionados con la enfermedad humana, como la localización intracelular obligada de algunas bacterias (clamidias y rickettsias) por insuficiencia de sus vías metabólicas para permitirles la vida libre. 9 El metabolismo de las bacterias heterótrofas no es esencialmente diferente del de las células de otros organismos como hongos, protozoos y animales 10 6. División bacteriana La división bacteriana se hace por un proceso relativamente sencillo en relación a las células eucariotas que se denomina fisión binaria. La duplicación del cromosoma bacteriano constituye el paso previo a la división celular. La molécula de DNA se abre y cada una de las hebras se fija a la membrana, probablemente en un mesosoma, replicándose. Los dos cromosomas resultantes se separan por crecimiento de la membrana de la zona intermedia entre los mesosomas que hacen las funciones de un aparato mitótico primitivo. Posteriormente se sintetiza un septo bacteriano de doble pared que dividirá a las dos células hijas. Ver vídeo: http://www.youtube.com/watch?v=DY9DNWcqxI4&feature=player_detailpage Nutrición bacteriana. En condiciones naturales las bacterias crecen en lugares en los que encuentran los elementos necesarios para su multiplicación, que en las bacterias quimiorganoheterótrofas, como las patógenas para el hombre, son substratos orgánicos así como agua e iones minerales. Si bien numerosas bacterias de vida libre son capaces de crecer con substratos simples como única fuente de carbono y energía, muchas de las bacterias adaptadas a la vida parasitaria precisan de la presencia de diversas moléculas orgánicas que no son capaces de sintetizar, por carecer de los sistemas enzimáticos necesarios. A estas moléculas se las denomina factores de crecimiento y esas bacterias se denominan auxótrofas para esta determinada sustancia. Si se incluyen en un tubo de ensayo todos los elementos necesarios para la multiplicación in vitro de una bacteria, como el agua, sales minerales, azúcares, proteínas y factores de crecimiento, si son necesarios, se obtiene un medio de cultivo (Figura 15). 7. Medios de cultivo Los medios de cultivo, al permitir la multiplicación bacteriana in vitro facilitan el aislamiento de las bacterias a partir de sus hábitats naturales y permiten su propagación en el laboratorio. 11 Los medios de cultivo pueden ser líquidos, cuando los componentes nutritivos se hallan disueltos en solución acuosa, o sólidos, cuando esta solución de nutrientes se gelifica al añadirle un polisacárido gelificante no degradable por la actividad metabólica de las bacterias como el agar-agar. El agar-agar se disuelve en el medio de cultivo líquido por ebullición y al enfriarse por debajo de los 45ºC solidifica como una gelatina endurecida dando al medio la consistencia de la carne de membrillo. Los medios líquidos se dispensan en matraces o tubos de ensayo y los sólidos se reparten en placas circulares provistas de tapa, denominadas placas de Petri, o en tubos de ensayo. La ventaja de los medios sólidos dispuestos en placas de Petri consiste en que en ellos pueden aislarse de modo individualizado las distintas bacterias de una mezcla contenidas en un material clínico (orinas, pus, etc.). En efecto, si se inocula (siembra) un medio líquido con una mezcla de bacterias (o una muestra clínica que contenga diversos tipos de bacterias) éstas se multiplican en el medio líquido entremezclado, en suspensión. Si por el contrario mediante un asa10 se disemina por la superficie de un medio sólido el material que contiene las bacterias (siembra por agotamiento), cada célula bacteriana se multiplica en el lugar donde ha quedado depositada, dando lugar, al cabo de unas horas o días, a millones de células que forman un conglomerado celular, visible macroscópicamente, denominado colonia bacteriana (Figura 15). Ver vídeo: http://www.youtube.com/watch?v=_1KP9zOtjXk&feature=player_detailpage Como algunas células bacterianas han quedado depositadas en la superficie del medio aisladas de las demás, a partir de ellas se forman colonias aisladas constituidas por un solo tipo de bacteria. Las bacterias de estas colonias pueden transferirse, mediante el asa a otros medios de cultivo, líquidos o sólidos, en los que crecen en cultivo puro. A partir de los cultivos puros se efectúa la identificación, se evalúa la sensibilidad a los antibióticos o pude realizarse cualquier otro estudio biológico. 10 El asa está formada por un mango semejante a un lápiz con un filamento recto y fina de metal que acaba en un bucle que permite tocar y sembrar el material biológico 12 Condiciones fisicoquímicas de los cultivos bacterianos. Para que se produzca la multiplicación, las bacterias deben sembrarse en medios que posean además de los nutrientes necesarios, el pH y la osmolaridad adecuados. Estos medios de cultivo deben incubarse en estufas a la temperatura óptima de crecimiento que, para la mayoría de las bacterias de interés médico, se sitúa entre 35º y 37ºC (bacterias mesófilas)11. Asimismo la atmósfera necesaria durante la incubación depende del tipo respiratorio de las bacterias, que como se ha señalado pueden ser aerobias, anaerobias, facultativas o microaerófilas. Dinámica del crecimiento bacteriano. Cuando se siembra una bacteria en un medio líquido originalmente transparente, después de unas horas de incubación éste se enturbia como consecuencia del crecimiento bacteriano. En la superficie de los medios sólidos, se desarrollan colonias como ya se ha indicado. La cinética de la multiplicación bacteriana se establece experimentalmente midiendo variaciones de la masa bacteriana en función del tiempo. Éste cálculo conduce a la obtención de una curva de crecimiento (Figura 16). El tiempo de generación es el tiempo necesario para que tenga lugar una división bacteriana, doblándose por tanto la masa bacteriana existente: De él depende el mayor o menor tiempo necesario para que se visualice una colonia sobre el medio sólido o se observe el enturbiamiento de un medio líquido. El tiempo de generación depende de la presencia de substratos energéticos, de la temperatura de incubación y otros factores externos, pero es fundamentalmente una característica propia de cada especie bacteriana, variando de una a otra. La mayoría de las bacterias de interés médico tiene un tiempo de generación de alrededor de 20 minutos detectándose macroscópicamente el crecimiento en los medios de cultivo (turbidez o colonias) tras 18 o 24 horas de incubación. Algunas bacterias poseen tiempos de generación más prolongados como Mycobacterium tuberculosis, causante de la tuberculosis, que posee un tiempo de generación superior a 24 horas, requiriendo semanas para formar colonias visibles macroscópicamente en los medios sólidos. 11 Bacterias psicrófilas: crecen entre 4-20°C; bacter ias mesófilas: crecen entre 25-35°C y bacterias termófilas: crecen entre 40-45°C 13 Utilización de los medios de cultivo. Según su función los medios de cultivo pueden dividirse en medios de aislamiento, medios de propagación y medios de identificación. Los medios de aislamiento se utilizan para aislar las bacterias a partir de los materiales o productos donde naturalmente se hallan (tierra, aguas, superficies cutaneomucosas y tejidos u órganos infectados). Los medios de aislamiento pueden clasificarse 1) usuales, 2) enriquecidos, 3) selectivos, 4) diferenciales y 5) de enriquecimiento. Para su descripción es preciso comprender el concepto de medio usual al que se ha hecho referencia anteriormente. Los medios usuales son medios de formulación sencilla que incluyen agua, proteínas obtenidas de un extracto o infusión de carne, glucosa y sales minerales, particularmente cloruro sódico y oligoelementos. Las bacterias que crecen en él son las que no poseen requerimientos de factores esenciales y se denominan no exigentes. Las bacterias exigentes son las que requieren diversos factores esenciales para su crecimiento y se cultivan en medios enriquecidos que aportan estos factores. Los medios enriquecidos se preparan añadiendo a un medio usual sangre, suero, extractos de levadura, que aportan esos factores o incorporándolos purificados, como el factor V (nicotin-adenin-dincleótido, NAD), el factor X (hemina), la cisteina u otros. Los medios enriquecidos más utilizados en bacteriología son el agar usual con sangre (agar sangre) y el agar-chocolate (agar usual enriquecido con los factores X y V). Cuando se pretende aislar todas las bacterias de un producto se siembra por agotamiento un medio sólido usual o preferentemente enriquecido. Sin embargo, es habitual que de un producto que contenga una mezcla de diversas especies bacterianas se desee aislar tan solo una especie en particular, que está en cantidades muy escasas con relación al resto de bacterias acompañantes, como una 14 salmonela de las heces o una legionela del esputo12. Para conseguir este objetivo pueden utilizarse medios selectivos o de enriquecimiento. Los medios selectivos son medios sólidos, usuales o enriquecidos, a los que se ha añadido alguna sustancia que inhibe el crecimiento de las bacterias, excepto las de la especie que se desea aislar. Diversos productos consiguen este efecto, siendo los más utilizados algunos colorantes con actividad antibacteriana selectiva, las sales biliares y los antibióticos entre otros. Los medios de enriquecimiento son medios líquidos que se utilizan para incrementar la cantidad de células bacterianas de una especie existente en una mezcla. Su actividad depende, como en los selectivos, de la incorporación de sustancias con actividad inhibitoria para determinadas bacterias. Durante la incubación se produce la multiplicación en el medio líquido de las bacterias que se desean aislar y se inhibe la de las demás bacterias. Tras la incubación, cuando la especie buscada ya está en una proporción muy superior a la de las bacterias que le acompañaban inicialmente en la muestra clínica, se efectúa una resiembra en un medio selectivo sólido. Los medios diferenciales son medios sólidos que incorporan a su formulación un substrato cuya utilización o no por las diferentes especies bacterianas da lugar a colonias de aspecto diferente; especies cuyas colonias en medios usuales aparecerían como idénticos. Los substratos más utilizados son azúcares (lactosa, sacarosa, etc.) que se añaden al medio junto a un colorante que es indicador de pH (rojo de fenol, azul de bromotimol, etc.). Las bacterias que no utilizan el substrato dan lugar a colonias con el color original del medio al no modificar el pH. Las que lo utilizan acidifican el medio con sus catabolitos y dan lugar a colonias de otro color al virar el indicador de pH. Los medios cromogénicos son medios diferenciales que incorporan substratos químicos sintéticos, que unidos a sustancias naturales son incoloros. Algunas bacterias, mediante enzimas específicas pueden hidrolizar estos enlaces y liberar compuestos de color intenso (Figura 17). La mayoría de medios selectivos se formulan a la vez como diferenciales, funcionan por tanto como medios selectivos-diferenciales. 12 Las heces y el esputo contienen decenas o centenas de diferentes especies bacterianas 15 8. Taxonomía bacteriana Las bacterias se clasifican en grupos taxonómicos, siendo la familia, el género y la especie los más operativos desde el punto de vista de la microbiología médica. La unidad taxonómica básica es la especie bacteriana. En la actualidad el criterio utilizado para establecer que dos bacterias pertenecen a una misma especie es el de la homología genética que existe entre ellas (taxonomía genética). Si se supone que la secuencia de bases del DNA de una bacteria es AGTCCGCTAT, se considerará con una homología del 100% las que poseen la misma secuencia AGTCCGCTAT. Si se supone que otra bacteria posee una secuencia ATTCCTAGTT, en la que sólo coinciden con la primera la mitad de las bases su homología con ella es del 50%. Se consideran como pertenecientes a la misma especia aquellas bacterias que poseen una homología genética superior al 80%. La técnica que permite determinar la homología genética entre dos bacterias es la de hibridación. Si se estudian los caracteres metabólicos de las bacterias, en la mayoría de los casos puede efectuarse una correlación entre éstos y el grupo taxonómico definido por hibridación. Las especies semejantes entre sí se agrupan en géneros y estos en familias. La familia y el género se denominan mediante un nombre científico latino (vg: familia Pseudomonadacae, género Pseudomonas). La especie se denomina mediante dos nombres latinos, el primero de los cuales es el del género y el segundo el propio de la especie (vg: Pseudomonas aeruginosa). A veces se utilizan como sinónimos nombres coloquiales vernáculos (vg: Pseudomonas aeruginosa, sinónimo bacilo piociánico). 9. Identificación de las bacterias La identificación de una bacteria como perteneciente a una determinada especie se establece en la práctica basándose en caracteres estructurales, metabólicos y antigénicos (fenotípicos). Ello es posible, a pesar de que los grupos taxonómicos se establecen por criterios genéticos, porque cada grupo definido por técnicas genéticas presenta caracteres fenotípicos y propios del grupo. 16 Los caracteres más universalmente estudiados son la morfología, la reacción al Gram y al Ziehl, la presencia de esporas y flagelos (movilidad), el tipo respiratorio, su exigencia en factores esenciales de crecimiento y la presencia de catalasa y citocromooxidasa que permiten diferenciar las bacterias en grandes grupos. Otros caracteres estudiados son su actividad metabólica sobre diversos substratos como azúcares, aminoácidos, lípidos, etc. Para estudiar estas propiedades metabólicas se utilizan medios de identificación que incorporan los substratos a estudiar. La metabolización de los substratos por la bacteria se detecta mediante indicadores de pH cuando comporta un cambio importante del pH del medio o mediante la detección de un catabolito específico producido en el proceso metabólico, mediante reactivos que sufren variaciones de color (como por ejemplo la detección del indol, producido por metabolización del triptófano, mediante el reactivo de Kovacs de color amarillo que vira a rojo). En la actualidad existen en el mercado sistemas de identificación que incorporan los diversos medios de identificación a micropocillos dispuestos en soportes de plástico. Estos sistemas han facilitado, mediante la realización sencilla de numerosas pruebas metabólicas, la identificación de las bacterias. Los antígenos que poseen valor en la clasificación bacteriana infraespécifica pueden detectarse, la mayoría de las ocasiones, por pruebas sencillas de aglutinación en porta mediante antisueros específicos. Aparte del serotipado existen otros métodos para la detección de grupos bacterianos diferentes dentro de una misma especie (clasificación infraespecífica), como son el fagotipado (lisotipado), el antibiotipado y otros basados en técnicas de biología molecular13. 10. Bacterias de interés en medicina Las bacterias de interés en medicina pueden clasificarse, según un criterio patogénico, en dos grupos: las que forman parte de la flora normal del hombre en la piel y las mucosas digestiva, respiratoria y urogenital, como simbiontes inocuos y las patógenas que poseen capacidad para producir enfermedad. Esta diferencia 13 Polimorfismo de fragmentos de restricción del ADN, ribotipado, diferencias en isoenzimas, y otros (ver más adelante) 17 práctica, en realidad no es tajante ni absoluta, ya que la capacidad patógena de un microorganismo depende tanto de él como de la capacidad de defensa del hospedador, lo que se refleja en dos hechos: 1) la existencia de portadores sanos de bacterias patógenas y 2) que bacterias de la flora normal pueden causar en determinadas circunstancias infecciones oportunistas graves. Las infecciones bacterianas se caracterizan por ser de curso agudo, con fiebre, leucocitosis, signos biológicos de reacción de fase aguda (VSG, proteína C reactiva y fibrinógeno elevados), y dar signos focales dependientes de los órganos afectados por la infección. Algunas bacterias como las brucelas (fiebre de Malta) o el bacilo de Koch (tuberculosis), después de la primoinfección, tienen capacidad para persistir en los macrófagos durante años en estado latente, pudiendo dar lugar a reactivaciones. Se dispone de numerosos medicamentos antibacterianos útiles, aunque su uso, en ocasiones indiscrimado, ha favorecido la selección de cepas resistentes. Sólo existen vacunas eficaces frente a algunas bacterias capsuladas y toxigénicas. A continuación se presentan sucintamente algunas bacterias de interés médico. La mayoría de estas bacterias se visualizan fácilmente al microscopio óptico y crecen bien en diversos medios de cultivo a temperatura óptima de 35 a 37ºC, en caso contrario se señala en el texto. Cocos grampositivos Los cocos grampositivos incluyen tres géneros de gran interés, Staphylococcus, Streptococcus y Enterococcus. Género Staphylococcus. Los estafilococos son cocos grampositivos que se agrupan en racimos. La especie Staphylococcus aureus [Sin: estafilococo dorado] es la que posee mayor capacidad patógena por producir numerosas enzimas exocelulares involucradas en la patogenicidad. Una de ellas, la coagulasa, permite diferenciar esta especie del resto de especies de estafilococos de interés en medicina, como S. epidermidis y otros, denominados colectivamente “estafilococos coagulasa negativa”. Los estafilococos se hallan fundamentalmente en la piel y mucosa nasal. S. aureus causa por mecanismo invasor, penetrando a las glándulas sebáceas y folículos 18 pilosos o a través de heridas, infecciones cutáneas, desde las que puede difundir por vía hemática, ocasionando infecciones generalizadas. Algunas cepas de S. aureus producen distintas exotoxinas que dan lugar a enteritis y otros procesos patológicos14. Los estafilococos coagulasa negativa son los agentes causales más frecuentes de infecciones oportunistas asociadas a cuerpos extraños como catéteres endovasculares, prótesis valvulares cardíacas y prótesis articulares, entre otras. Género Streptococcus. Los estreptococos son cocos grampositivos que se disponen en cadenas o parejas. Al cultivarlos en agar-sangre pueden dar lugar a diversos tipos de hemolisis de la sangre alrededor de las colonias por liberación de hemolisinas, lo que permite diferenciar 1) los estreptococos que poducen hemolisis total de la sangre y que se denominan β-hemolíticos, 2) los que producen hemolisis parcial, denominados α-hemolíticos o “viridans” y 3) los no hemolíticos. Entre los β-hemolíticos, que se diferencian en serogrupos15 por la estructura antigénica del carbohidrato C de su pared, se incluye Streptococcus pyogenes (Grupo A), que se halla en la faringe. Puede producir infecciones graves en la propia faringe y en la piel. Algunas cepas lisogénicas producen una exotoxina eritrogénica, que hace que la infección local invasora –faringitis o piodermitis– se acompañe de un exantema cutáneo característico causado por la exotoxina dando lugar a un cuadro clínico denominado escarlatina. Por otra parte en algunas personas, después de la infección invasora, pueden aparecer lesiones a distancia como consecuencia del desencadenamiento de una reacción inmunopatológia que da lugar a un cuadro clínico sistemático, la fiebre reumática, que se acompaña de inflamación aséptica local en las articulaciones (artritis) y en el endocardio (endocarditis). También se puede producir lesión en los glomérulos renales por mecanismo inmunopatológico (glomorulonefritis). Obsérvese pues, que esta especie es capaz de producir infección por los tres mecanismos patogénicos conocidos: invasor, toxigénico e inmune. 14 El shock tóxico y el síndrome de la piel escaldada 15 Grupos A, B, C, D, E, F, G, H, K, L, M, N y O de Lancefield. Se señalan en cursiva las que pueden encontrarse en el hombre 19 S. agalactiae es otro estreptococo beta-hemolítico (Grupo B) que se halla en el tubo digestivo y vagina pudiendo causar, además de infecciones urinarias, infecciones sistémicas perinatales (sepsis y meningitis) cuando el recién nacido se contamina a su paso por la vagina. No se conoce con precisión la vía de penetración de este microorganismo a los órganos profundos del recién nacido desde la colonización cutánea e intestinal (deglución) del que tiene lugar a su paso por la vagina. Los estreptococos del grupo viridans, que comprende varias especies de taxonomía mal definida, forman parte de la flora normal de la orofaringe. Streptococcus mutans, se considera directamente involucrado como agente causal de la caries dental, y el resto de especies, excepcionalmente pueden causar infecciones oportunistas (Tabla 1). Una especie particular de estreptococo α-hemolítico, el neumococo (Streptococcus pneumoniae), se halla en la faringe y posee gran capacidad para producir infección pulmonar severa –neumonía– seguida con frecuencia de bacteriemia que puede dar lugar a meningitis de pronóstico muy grave. Posee una cápsula polisacárida antigénica que ha permitido dividir la especie en más de 80 serotipos y que está involucrada en la patogenicidad por ser un factor de resistencia a la fagocitosis. Se ha preparado una vacuna polivalente con los antígenos capsulares de los 23 serotipos más frecuentes y otra conjugada heptavalente16. La conjuntiva y las mucosas de los senos paranasales y del oído medio están en comunicación con la faringe a través de sus conductos naturales y las infecciones bacterianas en esas localizaciones (conjuntivitis, sinusitis y otitis) están causadas por patógenos que residen en la rino y orofaringe: Streptococcus pneumoniae, Haemophilus influenzae, Streptococcus pyogenes y Staphylococcus aureus. 16 La vacuna 23-valente incluye los siguientes serotipos (nomenclatura danesa): 1, 2, 3, 4, 5, 6B, 7F, 8, 9N, 9V, 10A, 11A, 12F, 14, 15B, 17F, 18C, 19F, 19A, 20, 22F, 23F, 33F. Aunque la prevalencia de los serotipos patógenos varía según el país, se han señalado en cursiva los serotipos más prevalentes en nuestro medio. También son frecuentes los serotipos 6A, 7A y 15F, no incluidos en la vacuna. Recomendada en niños de más de dos años y adultos. La nueva vacuna conjugada de polisacáridos y proteínas heptavalente es eficaz en niños de corta edad. 20 El género Enterococcus, engloba un conjunto de especies (E. faecalis y otras17), semejantes a estreptococos, de hábitat intestinal. Los enterococos poseen creciente interés en el campo de la infección oportunista; ya que por su elevada resistencia a los antimicrobianos son seleccionados fácilmente por los antibióticos de amplio espectro. Cocos gramnegativos Género Neisseria. Los cocos gramnegativos de mayor interés en medicina pertenecen al género Neisseria. Morfológicamente se presentan formando parejas con aspecto de granos de café enfrentados por su cara plana. Son aerobios estrictos. El género posee varias especies comensales ubicadas en la orofaringe y dos especies patógenas N. meningitidis (meningococo), y N. gonorrhoeae (gonococo) cuyo único hábitat natural es el hombre. El meningococo se halla en la faringe, se transmite por vía aérea, y bajo determinadas circunstancias mal parecidas, puede producir bacteriemia para localizarse en las meninges causando se asocia a shock séptico. La meningitis meningococica es más frecuente en la infancia y suele presentarse en brotes epidémicos. Basándose en la estructura antigénica de los polisacáridos de la cápsula se han diferenciado 13 serogrupos, los más frecuentes de los cuales son el A, B, C, W-135 y el Y. Se han ensayado vacunas que han resultado eficaces frente a los serogrupos A y C pero el hombre responde mal a la inoculación del antígeno del grupo B que es el más frecuente en nuestro medio18. El gonococo causa infecciones genitales: de la uretra (uretritis), del cervix uterino (cervicitis) y del recto (proctitis) de transmisión sexual19. Tras la infección clínica 17 Las especies más frecuentes aisladas en clínica son: E. faecalis (80-90%), E. faecium (5-10%), E. avium, E. raffinosus y E. gallinarum, que se aíslan raramente. El aislamiento de otras especies es excepcional. Estos microorganismos causan infecciones urinarias, colecistitis, infecciones intraabdominales y pélvicas, sepsis y con menor frecuencia endocarditis bacteriana, así como infecciones en otras localizaciones 18 Ello ha conducido a ensayar vacunas utilizando como antígenos no el polisacárido, sino las proteínas de membrana externa 21 algunas personas pueden persistir como portadores asintomáticos del microorganismo en la uretra, vagina y recto y transmitir la enfermedad. En el género Neisseria existe un gran número de especies que son simbiontes inocuos en la orofaringe, entre las que cabe destacar N. catarrhalis por su potencial patógeno recientemente descubierto (infecciones respiratorias). Este microorganismo ha sido transferido a otro género bajo la denominación de Moraxella catarrhalis. Bacilos grampositivos Entre los bacilos grampositivos aerobios estrictos o facultativos, los géneros de mayor interés en medicina son Corynebacterium, Listeria, Bacillus y Lactobacillus20. Corynebacterium diphtheriae causa la difteria que se caracteriza por una faringitis muy exudativa que puede llegar a obstruir la vía respiratoria, con grave sintomatología general por producción de una potente exotoxina que al ser absorbida afecta al miocardio y nervios periféricos. Existen portadores sanos del microorganismo, que se transmite por vía aérea. La toxina está codificada por un bacteriófago lisogénico (fago β). El asilamiento de C. diphtheriae de la faringe no es fácil, requiere medios especiales y un cierto nivel de experiencia. Aunque la disposición de una vacuna antidiftéria eficaz ha erradicado en nuestro medio la enfermedad y disminuido el número de portadores, la vacunación inadecuada de la población podría hacerla reaparecer. Otras especies de este género, denominadas colectivamente “difteroides” o “difteromorfos21”, forman parte de la flora normal de la piel y mucosas y pueden causar, como oportunistas, infecciones sobre catéteres, sondas , prótesis y otros cuerpos extraños, una patología semejante a la de los estafilococos coagulasanegativa. 19 Las enfermedades cuya transmisión específica es por contacto sexual también se denominan venéreas 20 Excepcionalmente Erysipelothrix rhusiopathiae, bacilo grampositivo no esporulado que se encuentra en los peces puede infectar la piel humana a través de microheridas causando el erisipelatoide, infección cutánea eritematoviolácea que constituye una zoonosis profesional (pescateros, cocineros, etc.) 21 Corynebacterium jeikeium, C. urealyticum, C. pseudodiphthericum, C. pseudotuberculosis, C. xeroris, C. striatum, y el grupo no nominado G-2. C. minutissimum, causa una infección cutánea característica: la eritrasma 22 El género Listeria posee una especie, Listeria monocytogenes, psicrófila (crece a 4ºC), que se halla ampliamente distribuida en la naturaleza en particular en el tubo digestivo de numerosos animales. Puede contaminar al hombre de modo directo o a través de los alimentos. Una vez que ha colonizado el tubo digestivo humano puede persistir asintomáticamente o producir una patología en todo semejante a la del estreptococo ß hemolítico del grupo B (contaminación del neonato, sepsis con meningitis y meningitis primaria en el inmunodeprimido). El género Bacillus se distingue por su morfología característica, producir esporas y crecer en aerobiosis22. Dos especies poseen potencial patógeno para el hombre. Bacillus anthracis causa el carbunco en los animales y el hombre. La puerta de entrada de la infección es cutánea, dando lugar a una lesión local característica –la pústula maligna– desde donde puede producir sepsis. La inhalación de esporas causa una neumonía grave. El carbunco es una enfermedad zoonótica y profesional (pastores, matarifes, curtidores, etc). Bacillus cereus contamina con sus esporas, abundantes en el suelo, los alimentos y al germinar y multiplicarse en ellos libera toxinas que al ser ingeridas causan gastroenteritis. Lactobacillus es un género algunas de cuyas especies constituyen parte de la flora normal de la vagina. Bacilos gramnegativos Familia Enterobacteriaceae. Incluye más de 20 géneros y 100 especies de bacilos gramnegativos, aerobios y anaerobios facultativos que crecen bien en medios usuales. La mayoría tiene su hábitat natural en el medio ambiente (agua, tierra). Algunas especies se han adaptado al tubo digestivo de numerosos animales incluyendo el hombre, como Escherichia coli, Klebsiella pneumoniae o Proteus mirabilis, constituyendo parte de la flora normal. Estas bacterias, con E. coli a la cabeza, pueden causar infección oportunista grave (urinaria, abdominal, respiratoria y sepsis) y son junto a algunas bacterias anaerobias que desde su mismo hábitat producen infecciones semejantes, a Staphylococcus epidermidis, que causa infecciones asociadas a cuerpos extraños, los agentes causales más frecuentes de infección oportunista. 22 Carácter diferencial con el otro género esporulado de interés en patología humana (Clostridium), que es anaerobio estricto y con el que morfológicamente es indistinguible 23 Esta familia, incluye, además varios géneros patógenos para el hombre como Shigella, Salmonella, Yersinia y algunas cepas de la especie Escherichia coli que poseen factores de enteropatogenicidad comportándose como patógenos primarios (Tabla 2). Esta familia de microorganismos es una de las más estudiadas en bacteriología, en particular E. coli, que es la bacteria aerobia y anaerobia facultativa más abundante en el tubo digestivo del hombre, y que por su facilidad de cultivo a sido tomada como modelo de estudio de las células procariotas, siendo probablemente el ser vivo mejor conocido desde el punto de vista biológico (estructura , fisiología y genética). Una característica de interés de algunos miembros de esta familia es que distintos grupos dentro de una misma especie poseen potencialidades patogénicas muy diversas y ello no sólo en E. coli sino también en Salmonella y Yersinia, como se ha descrito en la Tabla 2. En otra familia de bacilos gramnegativos, Pseudomonadaceae, el género Pseudomonas y en particular P. aeruginosa posee características microbiológicas y de hábitat muy semejantes a las enterobacterias telúricas. Ocasionalmente pueden colonizar el tubo digestivo, la orofaringe o la piel del hombre, y causar infecciones oportunistas semejantes a las causadas por enterobacterias comensales23. Las pseudomonas forman parte de un amplio grupo de bacilos gramnegativos aerobios que se hallan en abundancia en las plantas y el medio ambiente denominados colectivamente bacilos gramnegativos no fermentadores24. Aparte de P. aeruginosa se encuentran en este grupo los géneros Burkholderia (B. cepacia, B. pseudomallei), Stenotrophomonas (S. maltofhilia). Bacterias de morfología espirilar. El género Vibrio está formado por bacilos gramnegativos con morfología curvada y su hábitat natural lo constituyen los ecosistemas acuáticos (marinos y de agua dulce). La especie Vibrio cholerae se 23 Aparte de P. aeruginosa existen otras especies del género Pseudomonas que también causan infecciones oportunistas aunque con menor frecuencia, como P. fluorescens y P. putida, P. stutzeri, etc. Existen dos especies, B. mallei y B. pseudomallei patógenas primarias para los animales y ocasionalmente el hombre, causantes del muermo y la meliodosis respectivamente; estas especies patógenas no se hallan en nuestro país (S. E. asiático, O. Medio, África, Australia). 24 Entre estos géneros de bacilos gramnegativos no fermentadores telúricos, que pueden causar infecciones oportunistas se hallan Acinetobacter, (A. baumanii), Alcaligenes y Flavobacterium, entre otros. 24 divide por su antígeno 0 (somático) en más de 80 serogrupos, pero sólo las cepas del serogrupo 1 (01) producen el cólera mediante liberación de una potente exotoxina tras colonizar la mucosa del intestino25. El cólera, que posee un gran potencial epidémico (pandemias), es una gastroenteritis toxigénica muy grave por la intensa deshidratación que causa, y en ausencia de tratamiento presenta una elevada tasa de mortalidad. Otros vibriones pueden también producir patología al hombre26. Género Campylobacter. Campylobacter jejuni, posee una morfología espirilar muy característica, es microaerófilo, se halla en el tubo digestivo de diversos animales, en particular a aves y causa enteritis al hombre27. Es de destacar que C. jejuni y las salmonelas gastroenteríticas son con mucho las bacterias más frecuentemente causantes de enteritis en nuestro medio. Helicobacter pylori, con características morfológicas y de cultivo semejantes a las campilobacterias tiene como hábitat natural la mucosa gástrica del hombre y se ha relacionado con las gastritis crónica, la úlcera duodenal y el cáncer de estómago y otros procesos patológicos. Bacterias hemófilas. Los hemófilos (género Haemophilus) son bacilos gramnegativos pleomórficos, que requieren para su crecimiento los factores X y V que se hallan en la sangre –de ahí su nombre– por lo que se cultivan en agar chocolate. La especie que causa infección con mayor frecuencia es H . influenzae. En términos nemotécnicos puede decirse que su hábitat natural, (orofaringe), y su capacidad patógena (neumonía, sepsis, meningitis, así como otitis, conjuntivitis y sinusitis) es semejante a la del neumococo. Además ambas bacterias poseen cápsulas como 25 Recientemente se han descrito cepas de otro serogrupo aparte el 01 (0139) productoras de toxina colérica 26 Los otros serotipos de V. cholerae y V. parahaemolyticus, pueden infectar al hombre por consumo de marisco, causando gastroenteritis. Las especies V. vulnificus, V. alginolyticus y otros pueden causar infecciones de heridas y otitis en bañistas 27 Otras especies de Campylobacter que causan enteritis al hombre con menor frecuencia son C. coli y C. lari. C. fetus puede colonizar el tubo digestivo y causar sepsis primaria oportunista; pero no causa gastroenteritis 25 factor de virulencia ya que les permite eludir la fagocitosis28. Existe una vacuna preparada con el antígeno capsular del serotipo b, que es el más prevalente en las infecciones humanas. Entre las demás especies del género (tabla 3) destaca H. ducreyi que produce el chancroide (úlcera genital con adenomegalia satélite) enfermedad de transmisión sexual, poco frecuente en nuestro medio. Bordetella pertussis es un cocobacilo gramnegativo aerobio estricto de morfología semejante a Haemophilus. Cuando infecta el árbol traqueobronquial causa una enfermedad respiratoria local acompañada de síntomas generales severos (tosferina). La patogenia de la enfermedad es compleja. La disposición de una vacuna eficaz, ha hecho que esta enfermedad sea poco frecuente en nuestro medio. Gardnerella vaginalis, morfologicamente semejante a los hemofilos, se asocia a una patología vaginal muy frecuente denominada vaginosis; que se caracteriza por: 1) desaparición de la flora normal de lactobacilos de la vagina, con aparición de Gardnerella y anaerobios 2) flujo moderado o abundante con olor fétido característico y 3) molestias vaginales imprecisas. A pesar de la presencia regular de este microorganismo en la vagina de las pacientes con este cuadro clínico no se conoce con precisión su papel patógeno en el proceso. Brucelas. El género Brucella está formado por bacilos gramnegativos pequeños, aerobios estrictos, que crecen bien en medios usuales, pero lo hacen lentamente. El género incluye varias especies siendo B. melitensis la más frecuente en nuestro medio. Esta especie infecta a los animales, en particular el ganado caprino y ovino produciéndoles diversos procesos patológicos (aborto epidémico). Pueden alcanzar al hombre en general a través de productos lácteos, como la leche y el queso, causando una grave enfermedad generalizada acompañada de septicemia persistente conocida como “fiebre de Malta29”. 28 En H. influenzae se han descrito diferentes inmunotipos de cápsula (a, b, c…), siendo el b el que produce con mayor frecuencia infecciones sistémicas en el hombre. Bajo determinadas circunstancias, cepas no capsuladas pueden también producir infecciones invasivas 29 Otras especies que producen al hombre un cuadro clínico semejante son B. abortus (ganado bovino) B. suis (ganado porcino) y más raramente B. canis (perros) 26 Las brucelas poseen interés conceptual por varias razones; en primer lugar por constituir un ejemplo claro de zoonosis (enfermedades de los animales trasmisibles al hombre) y en segundo lugar porque su capacidad biológica para persistir en los macrófagos tras ser fagocitadas –característica que comparten con las micobacterias– las faculta para producir infecciones recidivantes, a diferencia de las bacterias estudiadas hasta ahora cuyas infecciones poseen un carácter agudo. Legionela. El género Legionella está formado por bacilos o cocobacilos con estructura de bacterias gramnegativas. Se tiñen mal por el método de Gram. Son aerobios estrictos y requieren para su cultivo medios específicos en los que crecen lentamente (↑ 3 días) . Su hábitat natural lo constituyen los ambientes acuáticos. Algunas especies de entre las más de 22 existentes causan infección humana, la más frecuente es L. pneumophila30 (14 serogrupos) que al alcanzar al hombre por inhalación de vapor de agua contaminada produce una neumonía grave, o un síndrome febril más benigno, denominado fiebre de Pontiac, en particular en personas ancianas o con un déficit en las defensas antiinfecciosas. Se han descrito epidemias en instituciones causadas por el vapor de agua (aerosoles) de las duchas o calefacción, y en barrios de ciudades por vapor de agua contaminado eliminado a partir de industrias. Bacterias alcohol ácido-resistentes Un conjunto de bacterias cuya pared presenta la estructura básica de las bacterias grampositivas, poseen en ésta gran cantidad de ácidos grasos, entre ellos ácidos micólicos, que les confieren el carácter de la alcohol-ácido resistencia, lo que puede constatarse mediante la tinción de Ziehl-Neelsen. El género Mycobacterium es el más importante de este grupo31. Género Mycobacterium. Las micobacterias son bacilos que se tiñen difícilmente mediante la tinción de Gram, pero se observan como alcohol-ácido resistentes mediante la tinción de Ziehl. 30 Seguida por L. micdadei, L.bozemanii, L. dumoffii, L. longbeachae y otras Los géneros Nocardia y Rhodococcus también están constituidos por bacterias débilmente alcoholácido resistentes. Sus especies y en particular N. asteroides y R. equi pueden causar infecciones oportunistas pulmonares 31 27 Se hallan ampliamente distribuidas por la naturaleza constituyendo, como sucede en otras familias bacterianas, un espectro que va desde especies saprófitas telúricas no patógenas, hasta especies estrictamente adaptadas al hombre con gran capacidad patógena. Son aerobias estrictas y crecen en medios orgánicos complejos con yema de huevo y glicerol, siendo sus exigencias nutritivas variables y el tiempo de generación también variable, diferenciándose a este respecto en dos grupos, crecedoras lentas y crecedoras rápidas (> o < 5 días). Algunas especies producen regularmente pigmentos carotenoides (escotocromógenas), otras los producen tras inducción por la luz (fotocromógenas) y otras no los producen (no cromógenas). Estos caracteres –tiempo de crecimiento y pigmentación– se han utilizado para la clasificación inicial de las numerosas especies definidas en la actualidad (Tabla 4). M. tuberculosis (bacilo de Koch) es la especie más importante por su capacidad patógena causando la tuberculosis humana. Se caracteriza por su crecimiento lento y no producir pigmento. Su huésped natural es el hombre. Se transmite por vía aérea y produce principalmente infección pulmonar desde donde puede difundir a otros órganos por vía hematógena32 . Otras especies afines a M. tuberculosis, son M. africanum y M. bovis; esta última produce enfermedades en los bóvidos y puede transmitirse al hombre, y una micobacteria (M. bovis), modificada en el laboratorio por cultivos repetidos en medios glicerinados con lo que se consiguió la atenuación de su capacidad patógena, se denomina Bacilo de Calmette y Guerin (BCG) y es utilizada como vacuna. Otras especies de interés en patología son M. kansasii, M. scrofulaceum, M. avium – M. intracellulare y M. xenopi, entre los crecedores lentos y M. fortuitum y M. chelonae entre los rápidos. Mycobacterium leprae, causa la lepra. No ha podido ser cultivada en medios artificiales, pero sí propagada por inoculación en el cojinete plantar al armadillo. Su 32 Riñón, genitales, meninges 28 huésped natural es el hombre, aunque también se han documentado infecciones naturales en los armadillos. Bacterias anaerobias Las bacterias anaerobias incluyen un amplio conjunto de géneros y especies (cocos y bacilos grampositivos y gramnegativos) que crecen únicamente en ausencia de oxígeno (anaerobiosis) y requieren para su cultivo medios enriquecidos con sangre. Poseen una morfología relativamente característica, por lo que un observador experimentado puede sospechar su presencia al observarlas por el método de Gram. Se hallan formando parte de la flora autóctona de la orofaringe (surcos gingivodentales y criptas amigdalares) y del tubo digestivo, particularmente en el colon, constituyendo el grupo más abundante de la flora normal del intestino. Las bacterias anaerobias de mayor interés en medicina son las pertenecientes a los géneros Peptostreptococcus, que son cocos grampositivos, Bacteriodes, Prevotella, Porphyromonas y Fusobacterium, que son bacilos gramnegativos, y Eubacterium, Bifidobacterium, Propionibacterium33 y Actinomyces que son bacilos grampositivos no espurulados y Clostridium que es un género de bacilos grampositivos esporulados. Los géneros señalados, excepto Clostridium y Actinomyces que incluyen especies patógenas primarias, causan infecciones oportunistas. El género Clostridium está formado por bacilos grampositivos esporulados34 En él junto a especies comensales, existen otras de elevado poder patógeno por producción de exotoxinas como C. difficile que causa enteritis, C. perfringens y otras que causan gangrena gaseosa, C. tetani que causa el tétanos y C. botulinum que produce el botulismo (Tabla 5). 33 En el lenguaje coloquial suele referirse colecticamente como “bacteriodes” a los géneros Bacteriodes, Prevotella, Porphyromonas y Fusobacterium y como “corinebacterias anaerobias” a Eubacterium, Bifidobacterium y Propionibacterium 34 Recuérdese que Bacillus (aerobio) y Clostridium (anaerobio), son los únicos géneros bacterianos de interés médico que forman esporas 29 La vacuna antitetánica, elaborada con toxina tetánica inactiva (toxoide), es de extraordinaria eficacia. Las bacterias anaerobias no sólo requieren la anaerobiosis para su multiplicación, sino que el oxigeno es letal para ellas, por lo tanto las muestras clínicas en las que se sospecha su presencia (exudado, pus, esfacelos, etc) deben tomarse y trasladarse al laboratorio mediante técnicas adecuadas para evitar su contacto con el oxígeno atmosférico. Por descontado que la incubación de los medios de cultivo en que se siembran debe hacerse en anaerobiosis. Bacterias sin pared Género Mycoplasma. Los micoplasmas constituyen un grupo de bacterias carentes de pared, por lo que son pleomórficos, flexibles, frágiles y susceptibles a la lisis osmótica. Su tamaño es de 0.2 – 0.3 µm. por lo que algunas células son submicroscópicas. Poseen un genoma relativamente pequeño. En la naturaleza se hallan fundamentalmente asociados a las mucosas de los animales y a las plantas y pueden causar diversas enfermedades. La especie de mayor interés en medicina en M. pneumoniae que causa infección pulmonar (neumonía “atípica”)35. Los micoplasmas pueden manipularse y estudiarse por técnicas bacteriológicas convencionales, pero debido a que se tiñen mal con la tinción de Gram, requieren medios de cultivo complejos y crecen muy lentamente; para su detección en microbiología clínica se han adoptado técnicas alternativas al examen directo y el cultivo fundamentalmente pruebas serológicas y genéticas. Espiroquetas Las espiroquetas constituyen un grupo heterogéneo de bacterias definidas por su morfología espiral, presencia de fibras con estructura flagelar en el espacio periplásmico y movilidad característica. 35 El concepto de neumonía atípica se aplica a las neumonías que presentan una clínica diferente de la neumonía neumocócica, considerada como “típica”. Están causadas Mycoplasma, Chlamidia, Chlamidophila, Coxiella y virus respiratorios. Las legionelas, presentan un cuadro clínico más diferenciado y grave 30 La familia Spirochetaceae incluye tres géneros de interés en patología humana, Treponema, Borrelia y Leptospira. La mayoría de las espiroquetosis evolucionan por estadios con manifestaciones clínicas, separados por períodos de latencia asintomáticos de duración variable. Es difícil definir la importancia relativa del mecanismo invasor y el inmune en la patogenia de los estadios clínicamente expresivos. El género Treponema incluye especies patógenas para el hombre como T. pallidum causante de una enfermedad venérea, la sífilis, otras causantes de enfermedades no venéreas y varias especies no patógenas para el hombre (Tabla 6). Treponema pallidum, causante de la sífilis, es el patógeno humano más importante de este género. Tiene al hombre como único huésped natural. No se visualiza por el Gram, debido a su pequeño diámetro (0,15 x 10 µm) pero puede visualizarse por microscopia de campo oscuro y mediante tinciones argénticas. No crece en medios artificiales y para su propagación debe inocularse a animales de experimentación (testículo de conejo). La sífilis es una enfermedad de transmisión sexual, caracterizada porque la lesión inicial primaria en el lugar de inoculación, generalmente las mucosas genitales, es una úlcera indolora que cura espontáneamente. La enfermedad evoluciona, progresiva y asintomáticamente al estadio secundario y posteriormente con lentitud al terciario, afectando a numerosos órganos, como la piel, el hígado, el endocardio y el SNC y pudiendo transmitirse al feto por vía placentaria (sífilis congénita). El diagnóstico de la sífilis es fundamentalmente serológico. En el género Borrelia, B. recurrentis, causa la fiebre recurrente36, transmitida por piojos su reservorio es el hombre. Tras el período de inoculación (7 días) se produce una espiroquetemia con afectación del estado general, que suele curar después de una semana para reaparecer 2 a 4 días después. La enfermedad es endémica en algunas áreas, como África (Etiopia y Sudán), Asia y América del Sur. Otras especies como B. duttonii y B. hispanica que causan un cuadro clínico semejante, 36 El carácter recurrente de la enfermedad se debe a que cuando la respuesta inmune ha controlado la infección emergen en bacterias con variantes antigénicas que reactivan el proceso infeccioso al evadir la respuesta inmune inicial 31 tienen su reservorio natural en diversos roedores y alcanzan al hombre vehiculados por garrapatas del género Ornithodorus (Tabla 8). Las borrelias poseen un tamaño de 2-4 x 10-30 µm y pueden visualizarse en extensiones de sangre teñidas por el método de Giemsa. B. burgdorferi transmitida por garrapatas desde su reservorio animal (roedores) al hombre, causa la enfermedad de Lyme. Esta se caracteriza en una fase precoz por una lesión cutánea, el eritema crónico migrante cerca del lugar de inoculación por la garrapata y sintomatología “gripal”. Las manifestaciones tardías, que aparecen semanas o meses más tarde, son recurrentes y se caracterizan por producir artralgias, artritis, meningitis y miocarditis que pueden responder a un proceso preponderantemente inmunopatológico dada la escasez de bacterias observadas en esas lesiones. Aunque B. burgdorferi se tiñe por el método de Giemsa y puede crecer en medios artificiales su diagnóstico se hace habitualmente por serología. En el género Leptospira, L. interrogans no se observa por su finura con las tinciones bacteriológicas habituales (0,15 x 5-15 µm) crece en aerobiosis en medios enriquecidos. Causa diversas enfermedades en animales (ratas, perros, bóvidos) que las eliminan al medio (charcas) con la orina donde sobrevive varios días. Ocasionalmente se transmite al hombre desde el medio acuático a través de lesiones cutáneas o por ingesta causando bacteriemia seguida, tras un breve periodo asintomático, de afectación sistémica. La hepatitis, meningitis, afectación renal y vasculitis son las manifestaciones clínicas prominentes. Clamidias (géneros Chlamydia y Chlamidophila) Las clamidias son bacterias con estructura semejante a las gramnegativas. Efectúan síntesis proteica y biosíntesis, ya que poseen vías anabólicas diversas semejantes a otras bacterias heterótrofas, pero carecen de vías metabólicas para la obtención de ATP y la reoxidación del NADH, por lo que dependen de las células eucariotas, para obtener aporte energético, siendo por tanto parásitos intracelulares obligados. Presentan dimorfismo entre su estadio de multiplicación intracelular y la forma libre, lo que constituye uno de los escasos procesos de diferenciación celular del mundo bacteriano. 32 La partícula extracelular infecciosa presenta una estructura celular denominada cuerpo elemental de 0,3 µm, con un nucleoide denso. Es ingerida por la célula susceptible por endocitosis, situándose en una vacuola endocitaria. Esta partícula evolucionará a una estructura mayor, de 0,5 µm, denominada cuerpo reticulado que se divide para dar lugar a cuerpos elementales que son liberados por lisis celular. Se tiñen mal por el método de Gram, pero pueden colorearse por Giemsa u otras tinciones, mediante las cuales, además, puede observarse en las células la presencia de cuerpos de inclusión formados por el cuerpo reticulado. El examen microscópico convencional no se utiliza rutinariamente para su detección en tejidos y exudados. Se multiplican en cultivos celulares por lo que se estudian por técnicas semejantes a las empleadas en virología. El género Chlamydophila incluye dos especies de interés: C. psittaci que tienen su reservorio natural en diversas aves, C pneumoniae con reservorio humano, causantes de infecciones pulmonares (neumonía atípica). Chlamydia trachomatis, que produce diversos y heterogéneos procesos patológicos como el tracoma (conjuntivitis crónica folicular maligna), el linfogranuloma venéreo, uretritis, cervitis, conjuntivitis e infección pulmonar neonatal. Las clamidias poseen un antígeno común de grupo y antígenos específicos de especie o serotipo37. En C. trachomatis se han descrito 15 serotipos (A, B, Ba, C-K, L1-L3). Existe una relación específica entre cada serotipo y los diversos procesos patológicos señalados (Tabla 7); aunque no se conoce la causa de este hecho ni el porqué de la limitada distribución geográfica del tracoma y del linfogranuloma venéreo. Rickettsias Las rickettsias son bacilos gramnegativos aerobios, de pequeño tamaño, 0.3-2 µm, que se tiñen mal por el Gram y son parásitos intracelulares obligados, aunque no se conoce con exactitud las razones de esta dependencia; por todo ello fueron inicialmente confundidas con virus. 37 El antígeno común reside en el lipopolisacárido y los específicos en proteínas de la membrana externa 33 La familia Rickettsiaceae comprende cuatro géneros de interés en medicina: Rickettsia, Coxiella, Rochalimea y Ehrlichia. (Tabla 9). Todas las rickettsias presentan un reservorio natural animal y alcanzan accidentalmente al hombre para producir enfermedad a través de artrópodos vectores, excepto Coxiella burnetii que se transmite al hombre por inhalación o ingesta. Presenta una distribución geográfica localizada, o universal según la especie. Las especies del género Rickettsia, tras ser inoculadas al hombre mediante la picadura de un artrópodo producen una infección generalizada, afectando a numerosos órganos, localizándose en las células endoteliales de los pequeños vasos sanguíneos en los que se produce una vasculitis característica. Clínicamente se expresan por fiebre, afectación del estado general y exantema. R. prowazekii, cuyo reservorio natural es el hombre, es transmitida por piojos, causa el tifus epidémico38 enfermedad inexistente en la actualidad en nuestro país (África, Asia y Sudamérica). En los países de la cuenca mediterránea, incluyendo el nuestro, existe una ricketssiosis exantemática causada por R. conorii, transmitida por garrapatas, denominada fiebre botonosa mediterránea. Coxiella burnetii, de distribución universal, tiene como reservorio diferentes animales, bóvidos, cabras y ovejas en las que causa diversas enfermedades, se elimina con las excretas y la leche y puede, por inhalación o ingestión, causar infección sistémica denominada “fiebre Q” que se manifiesta como un síndrome febril sin focalidad. En otras ocasiones la infección causa neumonía atípica y menos frecuentemente hepatitis. En la práctica las rickettsias no se visualizan mediante técnicas de microscopía convencional y se cultivan en líneas celulares como los virus. Por estas razones y por su elevada contagiosidad en el laboratorio, su diagnóstico suele efectuarse por métodos serológicos y otros. 38 La recurrencia del tifus epidémico que aparece años después de la infección se conoce como enfermedad de Brill 34 Figura 1. Estructura de la célula bacteriana 35 Figura 2. Membrana citoplasmática Está formada por una doble capa de fosfolípidos en la que se encuentran embebidas proteínas que pueden desplazarse lateralmente. 36 Figura 3. Fosfoglicéridos Están constituidos por una molécula de glicerol cuyos dos primeros hidroxilos se encuentran esterificados por moléculas de ácidos grasos, mientras que el hidroxilo del tercer átomo de carbono está esterificado por el ácido fosfórico el cual esterifica a su vez a un alcohol o aminoácido. 37 Figura 4. Peptidoglicano Es un polímero de dos aminoazúcares (N-acetil-glucosamina y N-acetil-murámico) que se alternan. Estos polímeros se entrelazan dando lugar a una malla rígida: del N-acetil-murámico se originan unos tetrapeptidos que se unen entre sí mediante puentes peptídicos. 38 Figura 5. Diferencias en la pared entre las bacterias grampositivas y gramnegativas En las bacterias grampositivas el peptidoglicano es muy grueso. En las bacterias gramnegativas, el peptidoglicano es una malla fina, existiendo por fuera del mismo una membrana externa que no se encuentra en las bacterias grampositivas. 39 Figura 6. Estructura de la pared celular de las bacterias grampositivas LTA: Ácidos teicoicos; WTA: Proteínas fibrilares 40 Figura 7. Estructura de la pared celular de las bacterias gramnegativas 41 Figura 8. Liposacarido Está constituido por el lípido A inmerso en la membrana externa y una cadena polisacarida que sobresale hacia el exterior. El lípido A es un glicofosfolípido formado por un disacárido de N-acetil-glucosamina unido a grupos fosfato y a varias moléculas de ácidos grasos de cadena larga. En el polisacarido se distingue una parte interna formada por 5 a 7 azúcares comunes a diversos grupos bacterianos y una parte externa o cadena lateral formada por una secuencia de 2 a 3 azúcares que se repiten n veces. 42 Figura 9. Cápsula Estructura fibrilar polimérica, de naturaleza generalmente polisacárida que se sintetiza en el interior de la célula y se exporta al exterior. Por su actividad antifagocitaria constituye un importante factor de virulencia. Su visualización en el laboratorio de microbiología suele hacerse por tinciones negativas. 43 Figura 10. Flagelos Estructuras fibrilares de naturaleza proteica, formadas por la agregación de múltiples subunidades de una proteína globular denominada flagelina. Constituyen el órgano de locomoción de las bacterias. 44 Figura 11. Disposición de los flagelos en la superficie bacteriana Según esta, las bacterias flageladas pueden subdividirse en: monotricas (un solo flagelo), periticas (varios flagelos distribuidos por toda la superficie bacteriana) y lofotricas (varios flagelos formando un penacho). 45 Figura 12. Esquema de la fermentación de glucosa según Embden-Meyerhof Es la vía más universal. Lleva al ácido pirúvico que es el catabolito central de otras vías fermentativas (metabolismo anaerobio) y de la vía aerobia representada por el ciclo de Krebs. 46 Figura 13. Vías fermentativas del ácido pirúvico. Incluye, entre las más importantes: láctica, alcohólica, ácida mixta, butilenglicolica, butírica, propionica. En cualquiera de ellas, el aceptor de electrones liberados en los procesos oxidativos de los sustratos metabólicos es un compuesto orgánico. 47 Figura 14. Ciclo de Krebs Los microorganismos con metabolismo aerobio pueden llevar los catabolitos producidos en las fermentaciones a niveles energéticos aun más bajos obteniendo mayor energía. En este proceso, se produce una oxidación del sustrato con transferencia de los electrones al oxígeno a través de la cadena respiratoria. 48 Figura 15. Colonia bacteriana Cuando las bacterias contenidas en un inóculo se diseminan por la superficie del agar, se dividen en el lugar donde han quedado depositadas dando lugar al cabo de unas horas a una masa celular macroscópicamente visible denominada colonia bacteriana. 49 Figura 16. Curva de crecimiento La cinética del crecimiento bacteriano se establece experimentalmente midiendo las variaciones de la masa bacteriana en función del tiempo. Se observan cuatro fases: a) tiempo de latencia, b) crecimiento exponencial, c) estacionaria y d) muerte celular. 50 Figura 17. Medio cromogénico Las colonias de las bacterias adoptan diferente color, en función del sustrato del medio que pueden hidrolizar. 51 Tabla 1. Estreptococos viridans Bajo el nombre de estreptococos viridans se incluye un conjunto de estreptococos para los que no existe unanimidad en su clasificación, difíciles de identificar a nivel de especie mediante caracteres metabólicos convencionales, que son generalmente alfa-hemolíticos, sensibles a la vancomicina y productores de la enzima leucinaaminopeptidasa. Las distintas especies se han incluido en varios “grupos” en razón de la semejanza de sus caracteres metabólicos. En la práctica, por las razones señaladas, no se identifican a nivel de especie y se hace referencia a ellos como estreptococos “viridans”. Son comensales de la cavidad orofaríngea y algunos se encuentran en el tracto respiratorio superior (“S. milleri”) y tracto intestinal (“S. bovis” y “S. milleri”). Grupos Especie 1 Patología específica Grupo del S. mutans: S. mutans S. sobrinus S. rattus S. cricetus Caries Endocarditis Grupo del S. salivarius: S. salivarius S. vestibularis S. intestinalis S. thermophilus Bacteriemia en pacientes con cáncer Endocarditis Streptococcus bovis: S. bovis I S. bovis II Bacteriemia en pacientes con cáncer Endocarditis Grupo “Streptococcus milleri”: Endocarditis Grupo del S. sanguis: S. anginosus S. constellatus S. intermedius S. sanguis S. gordonii S. parasanguis S. crista Endocarditis Streptococcus mitis: S. mitis Endocarditis Otros estreptococos viridans: S. acidomininus S. uberis Endocarditis S. defectivus S. adjacens Endocarditis Estreptococos piridoxal dependientes 2 1 Los estreptococos viridans pueden causar caries dental, endocarditis bacteriana subaguda (endocarditis lenta) y también pueden aislarse como contaminantes en hemocultivos y cultivos de líquido cefalorraquídeo, aunque cada vez con más frecuencia causan infecciones en neonatos y pacientes inmunodeprimidos 2 Constituye un grupo de estreptococos auxotróficos para el piridoxal, que sólo crecen en presencia de esta sustancia o en anaerobiosis 52 Tabla 2. Especies patógenas de la familia Enterobacteriaceae Géneros patógenos Especies Patología Observaciones Escherichia E. coli Gastroenteritis Sólo algunas cepas de esta especie son enteropatógenas Shigella S. sonnei S. flexneri S. boydii S. dysenteriae Gastroenteritis Gastroenteritis Gastroenteritis Gastroenteritis Causan una enteritis invasora (diarrea con sangre: disenterica). El único reservorio natural es el hombre. S. dysenteriae no es autóctona serotipo Typhi serotipo Paratyphi A serotipo Paratyphi B serotipo Paratyphi C Sepsis Sepsis Sepsis Sepsis El único reservorio conocido de estos serotipos es el hombre. El único serotipo septicemiante que se aísla con frecuencia en nuestro país es Typhi resto de serotipos: Enteritidis Typhimurium Wirchow etc. hasta 2000 Gastroenteritis Estos serotipos se hallan ampliamente distribuidos en el tubo digestivo de diversos animales. Y. pestis Peste Y. pseudotuberculosis Adenitis mesentérica Gastroenteritis Gastroenteritis Salmonella Yersinia Y. enterocolitica El hombre al infectarse puede quedar como portador asintomático y ser a su vez fuente de infección La peste es una enfermedad que afecta a los ratones y puede alcanzar al hombre a través de las pulgas. Sólo algunos serotipos de esta especie son patógenos para el hombre (O3; O5; O27; O8; O9; entre los más frecuentes). 53 Tabla 3. Especies de Haemophilus aisladas en patología humana. La especie que con más frecuencia causa infección en el hombre es H. influenzae del serotipo b. Causa una patología semejante al neumococo (infección respiratoria y de estructuras pararespiratorias), sepsis y meningitis. Algunas cepas no capsuladas pueden ocasionar infección respiratoria y colonización vaginal, contaminando al neonato a su paso por el canal del parto y causando infección neonatal (sepsis con meningitis). Algunas especies tras producir bacteriemia transitoria pueden causar endocarditis en pacientes con alteraciones valvulares. Especie Frecuencia Distribución Patología H. influenzae frecuente Universal Infección respiratoria Sepsis Meningitis Países cálidos Conjuntivitis, fiebre purpúrica de Brasil Universal Endocarditis H. aegypticus 1 H. parainfluenzae poco frecuente H. aphrophilus poco frecuente Endocarditis H. paraphrophilus poco frecuente Endocarditis H. ducreyi Asia y África H. hemolyticus raro H. parahemolyticus raro Chancroide 1 H. aegypticus corresponde al biotipo III de H. influenzae y causa específicamente conjuntivitis y la fiebre purpúrica de Brasil 54 Tabla 4. Principales especies de Mycobacterium Solo M. tuberculosis tiene su reservorio animal en el hombre. El resto de especies se hallan en los animales (M. bovis) o son de hábitat telúrico. Los órganos lesionados que se señalan son los afectados con mayor frecuencia en personas previamente sanas. Las micobacterias pueden alcanzar otros órganos por vía hemática, particularmente en pacientes inmunodeprimidos. Especie Patología prevalente (localización) Crecedores lentos (>5 días) * No cromógenos M. tuberculosis complex (M. tuberculosis, M. bovis, M. africanum, BCG) M. avium – M. intracellulare M. ulcerans M. malmoense M. haemophilum Pulmón Pulmón Piel Pulmón Piel * Fotocromógenos M. kansasii M. marinum M. simiae Pulmón Piel Pulmón * Escotocromógenos M. scrofulaceum M. szulgai M. xenopi Ganglios linfáticos Pulmón Pulmón Crecedores rápidos (<5 días) M. fortuitum-chelonae complex Piel Cuerpos extraños Especies no cultivables M. leprae Piel, S.N.C. (lepra) 55 Tabla 5. Principales bacterias anaerobias Las bacterias anaerobias se encuentran como comensales en la piel, orofaringe, tracto intestinal bajo, vagina y uretra terminal. Las infecciones causadas por anaerobios suelen tener un origen endógeno y ser polimicrobianas. La identificación de estos microorganismos a nivel de especie no suele ser útil a no ser que tengan características particulares de patogenicidad o resistencia que les diferencien de las de su género. Especie Cocos grampositivos Gen. Peptostreptococcus P. magnus Consideraciones Posible resistencia a la clindamicina Cocos gramnegativos Gen. Veillonela Bacilos gramnegativos Gen. Bacteroides B. fragilis B. thetaiotaomicron, B. distasonis, B. ovatus, B. uniformis Gen. Porphyromonas Gen. Prevotella P. disiens P. bivia Gen. Fusobacterium F. necrophorum F. nucleatum F. mortiferum F. varium Bacilos grampositivos no esporulados Gen. Eubacterium Gen. Bifodobacterium Gen. Propionebacterium Gen. Actinomyces Gen. Mobiluncus Gen. Lactobacillus Gen. Ramibacterium Bacilos grampositivos esporulados Gen. Clostridium C. tetani C. botulinum C. septicum C. perfringens C. difficile C. ramosum C. clostridioforme, C. innocum Distribución Presentes en cavidad oral, tracto gastrointestinal y genitourinario Presente en cavidad oral Uno de los aislados con mayor frecuencia Mayor resistencia a los antibióticos que B. fragilis Asociadas a infección del tracto genital femenino Gran virulencia Es el género predominante en el tracto intestinal. El complejo B. fragilis interviene con gran frecuencia en las infecciones de la cavidad abdominal. Los bacteroides pigmentados están presentes en la cavidad oral y suelen asociarse a infecciones pulmonares Similar a Bacteroides Similar a Bacteroides Presentes en vagina De distribución similar a los Bacteroides, suelen aislarse como causa de infección respiratoria Resistencia a la penicilina y clindamicina Asociado a infección del tracto genital femenino Presentes en cavidad oral y en vagina Presentes en cavidad oral Posible poder patógeno en ciertas Presentes en los folículos y en las glándulas sebáceas de circunstancias la piel Suele requerir de tratamientos prolongados Asociados a vaginosis Presentes en vagina Presentes en vagina Causante del tétanos Causante del botulismo Se ha asociado con tumores malignos gastrointestinales Puede causar infecciones graves Presentes en el tubo digestivo del hombre y la mayoría de animales, sus esporas se encuentran en gran número en el suelo Causa de colitis pseudomembranosa asociada a terapéutica antibiótica Posible resistencia a la penicilina, clindamicina y cefalosporinas 56 Tabla 6. Treponematosis humanas El pián, la pinta y el bejel son infecciones cutáneas que se transmiten por contacto directo. El reservorio natural de sus agentes es el hombre. Especie Distribución Patología T. pallidum ss pallidum Mundial Sífilis T. pertenue Trópicos (África, América, Asia) Frambesia (Pián) T. carateum Trópicos (América) T. pallidum ss endemicum Distribución irregular en zonas cálidas y pobres África sahariana, Asia) Pinta Bejel 57 Tabla 7. Infecciones por Chlamydia y Chlamydophila Especie Chlamydophila psittaci Muchos serotipos mal sistematizados (Distribución universal) Chlamydophila pneumoniae Un serotipo (Distribución universal) Patología Su principal reservorio lo constituyen las aves domésticas y salvajes. La infección humana es una zoonosis contraída por la inhalación de secreciones respiratorias o polvo de las excretas de aves infectadas. El cuadro clínico es de neumonía atípica. Su reservorio es humano. Causa infección del tracto respiratorio alto (faringitis, sinusitis) y bajo (bronquitis, neumonía atípica). Su transmisión es aérea. Chlamydia trachomatis Serotipos A, B, Ba, C (Prevalente en regiones desérticas o semidesérticas de África, India, Pakistán, Indonesia y Australia) Causan el tracoma; grave conjuntivitis folicular crónica, que constituye una de las causas más importantes de ceguera evitable en las areas endémicas. Serotipos D, E, F, G, H, I, J, K (Distribución universal) Uretritis y cervicitis de transmisión sexual que pueden complicarse con epidedimitis en el varón, endometritis, salpingitis y pelviperitonitis (enfermedad inflamatoria pélvica) en la mujer, y proctitis en ambos sexos. Conjuntivitis de inclusión en adultos. El neonato puede contaminarse a su paso por el canal del parto. La mayoría presenta conjuntivitis de inclusión. En el 5-10% puede observarse una neumonía neonatal. Serotipos L1, L2, L3 (Prevalentes en África y Sudamérica) Dan lugar a linfogranuloma venéreo, de transmisión sexual. Se caracteriza por una lesión inicial ulcerada en los genitales, que se resuelve espontánea y rápidamente. Posteriormente aparece linfoadenopatía regional fistulizada que se sigue tardiamente de reacción fibrótica y alteración del drenaje linfático. 58 Tabla 8. Especies de Borrelia causantes de fiebre recurrente Las fiebres recurrentes están causadas por numerosas especies de Borrelia que tienen como reservorio al hombre o diversos roedores y son transmitidas por piojos y garrapatas. Su distribución geográfica es irregular. Especie Vector Reservorio Distribución Patología B. recurrentis Pediculus humanus humanus (piojo1) Humano Mundial Fiebre recurrente epidémica B. duttoni Ornithodoros moubata Humano África (C, E y S) Fiebre recurrente endémica de EAfrica B. hispanica Ornithodoros erraticus Roedores España, Portugal, Fiebre Marruecos, Argelia recurrente y Túnez hispano-africana B. burgdorferi Garrapatas del género Ixodes Roedores E-O Estados Unidos, Europa, Asia y Australia África, Asia, América y caúcaso (según hábitat del vector) Otras bacterias: Ornithodoros spp B. crocidurae, Roedores y otros pequeños mamíferos Enfermedad de Lyme Fiebres recurrente B. merionesi, B. microti B. dipodilli, B. persica, B. caucasica, B. latyschewii, B. hermsii, B. turicatae, B. parkeri, B. mazzottii, B. venezuelensis 1 El piojo se infecta al ingerir sangre de un enfermo, pero la infección no se transmite por vía transovárica 59 Tabla 9. Rickettsias y rickettsiosis R. sibirica (Asia Central) y R. australis (Australia) causan infecciones sistémicas con exantema transmitidos por garrapatas. R. akari (V.S. Rusia) es transmitida por ácaros. Organismos Vector Reservorio Distribución Patología R. prowazekii Piojo humano Humano África, Asia, S. América Tifus exantemático epidémico R. typhi Pulgas Roedores Mundial Tifus marino R. rickettsii Garrapata Roedores, perros N. y S. América Fiebre exantemática de las Montañas rocosas R. conorii Garrapata Roedores Cuenca mediterránea Fiebre botonosa mediterránea Rochalemaea quintae Piojo humano Humano Europa, Asia, África Fiebre de las trincheras Inoculación Ganado Mundial Fiebre Q 2 Erlichia, Anaplasma 1 y Neorikettsia Coxiella burnetii 2 1 Son un grupo de zoonosis transmitidas por garrapatas. Con excepción de N. sennetsu, especie confinada al área de Malasia-Japón, que produce un cuadro similar a la mononucleosis infecciosa, y de la que desconocemos su ciclo, las Erlichia spp y Anaplasma spp se mantienen en un ciclo garrapata, rumiante, roedor. Provocan un cuadro pseudogripal consistente en fiebre, malestar general, artromialgias y cefalea. Algunos pacientes desarrollan neumonía atípica y en el caso de E. chaffeensis alteraciones del S.N.C. (meningitis). 2 Microorganismos que causan enfermedad en nuestro medio 60