CÉLULAS GERMINALES PRIMORDIALES Y DETERMINACIÓN
Anuncio

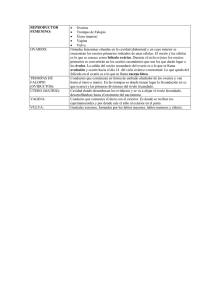
Biología molecular de LA CÉLULA – 4a. Edición LAS VENTAJAS DEL SEXO CÉLULAS GERMINALES PRIMORDIALES Y DETERMINACIÓN DEL SEXO EN LOS MAMÍFEROS Las estrategias de la reproducción sexual pueden variar muchísimo de unos organismos a otros. En esta apartado nos centramos principalmente en las estrategias utilizadas por los mamíferos. En todos los embriones de los vertebrados, durante el desarrollo tiene lugar una selección temprana de determinadas células como progenituras de los gametos. Estas células germinales primordiales emigran para desarrollar las gónadas, que serán los ovarios en las hembras y los testículos en los machos. Después de un período de proliferación mitótica, las células germinales primordiales experimentan la meiosis y se diferencian en gametos maduros -oocitos o espermatozoides-. Más adelante, la fusión del oocito y del espermatozoide tras el apareamiento conduce al inicio de la embriogénesis. Con la posterior producción de nuevas células germinales primordiales en el embrión, comienza otra vez el ciclo. En este apartado, consideraremos cómo aparece una célula germinal primordial en los mamíferos, cómo se determina el sexo en ellos y cómo establece la determinación sexual que las células germinales primordiales se diferencien en espermatozoides o en oocitos. Las células germinales primordiales emigran a las gónadas en desarrollo En la mayoría de animales, incluidos muchos vertebrados, el oocito no fecundado es asimétrico, con regiones del citoplasma que contienen diferentes dotaciones de mRXA y de proteínas (tratado en el Capítulo 21). Cuando el oocito es fecundado y se divide repetidamente produciendo las células del embrión temprano, las células que heredan moléculas específicas de una determinada región del citoplasma del oocito se transforman en células germinales primordiales. Por el contrario, en los mamíferos, el oocito es muy simétrico y todas las células resultantes de las primeras divisiones del oocito fecundado son totipotentes -es decir, son capaces de producir cualquier tipo de células del organismo, incluidas las germinales-. En el embrión temprano de los mamíferos, un pequeño grupo de células es inducido a convertirse en células germinales primordiales a consecuencia de las señales que reciben de sus células vecinas. En el ratón, por ejemplo, una semana después de la fecundación, unas 50 células del tejido situado fuera del embrión propiamente dicho son inducidas por sus vecinas a transformarse en células germinales primordiales. Unos días después, estas células proliferan y son arrastradas hacia el interior del embrión con la invaginación del intestino posterior. Luego emigran por el intestino hacia su destino final, que son las gónadas en desarrollo (Fig. 20-16). Mientras las células germinales primordiales emigran a través del embrión, son señaladas para sobrevivir, proliferar y emigrar por varias proteínas extracelulares producidas por las células somáticas adyacentes. Después de que las células germinales primordiales entren en la gónada en desarrollo del ratón, que en este estadio se llama cresta genital, continúan proliferando 2 o 3 días más. En este momento, escogen y siguen una vía de desarrollo que las llevará a transformarse en oocitos o en espermatozoides, dependiendo no de su propio sexo Ediciones Omega 1 Biología molecular de LA CÉLULA – 4a. Edición LAS VENTAJAS DEL SEXO cromosómico sino de si la cresta genital ha empezado a diferenciarse en un ovario o en un testículo. Los cromosomas sexuales de las células somáticas de la cresta genital determinan el tipo de gónada que se va a formar. Un solo gen del cromosoma Y desempeña un papel especialmente importante en esta decisión. Figura 20-16 Migración de las células germinales primordiales en los mamíferos. (A) Esquema que muestra las etapas finales de la migración a través del intestino posterior hasta las dos crestas genitales, cada una de las cuales desarrollará una gónada -un ovario o un testículo-. (B) Micrografía en la que se observan las células germinales primordiales de un embrión temprano de ratón. Las células germinales primordiales están teñidas con un anticuerpo monoclonal (verde) que las marca específicamente en este estado de la embriogénesis. Las otras células del embrión están teñidas con una lectina que se une al ácido siálico, que se encuentra en la superficie de todas las células. (C) Esquema correspondiente a la micrografía anterior. (B, por cortesía de Robert Anderson y Chris Wylie.) El gen Sry del cromosoma Y puede determinar Ediciones Omega 2 Biología molecular de LA CÉLULA – 4a. Edición LAS VENTAJAS DEL SEXO que un embrión hembra se transforme en macho Aristóteles creía que la temperatura del macho durante el contacto sexual determinaba el sexo de la descendencia: a temperatura más alta, mayor era la probabilidad de producir machos. Actualmente sabemos que el sexo de un mamífero viene determinado por sus cromosomas sexuales, más que por el medio ambiente donde vive (aunque en algunos animales como los cocodrilos y muchos peces sucede lo contrario). Las hembras de los mamíferos tienen dos cromosomas X en todas sus células somáticas, mientras que los machos tienen un cromosoma X y un cromosoma Y. El cromosoma Y es el factor determinante. Los individuos que tienen un cromosoma Y se desarrollan como machos siendo indiferente el número de cromosomas X que presenten, mientras que los individuos que no tienen ningún cromosoma Y se desarrollan como hembras, incluso aunque sólo tengan un cromosoma X. El espermatozoide que fecunda el oocito determina el sexo del zigoto resultante: el oocito tiene un solo cromosoma X; en cambio, el espermatozoide puede tener un cromosoma X o un cromosoma Y. El cromosoma Y determina el sexo del individuo, induciendo a las células somáticas de la cresta genital a diferenciarse en un testículo en lugar de un ovario. El gen crucial del cromosoma Y que tiene esta función determinante de testículo se denomina Sry. Cuando este gen se introduce en el genoma de un zigoto XX de ratón, el embrión transgénico resultante se desarrolla como un macho, aunque falten todos los demás genes del cromosoma Y (Fig. 20-17). Este ratón, sin embargo, no puede producir espermatozoides, porque la presencia de dos cromosomas X impide su formación. Sry se expresa solamente en un subgrupo de las células somáticas de la gónada en desarrollo y determina la diferenciación de estas células en células de Sertoli, que constituyen el principal tipo de células de soporte que hay en el testículo. Las células de Sertoli dirigen el desarrollo sexual de una línea masculina influyendo en otras células de la cresta genital, por lo menos de cuatro maneras: 1. Estimulando las células germinales primordiales recién llegadas para que se transformen en espermatozoides. 2. Mediante la secreción de la hormona anti Mülleriana, que bloquea el desarrollo del tracto genital femenino, produciéndose la regresión del conducto de Müller (que daría lugar al oviducto, al útero y a la parte superior de la vagina). 3. Estimulando la migración de unas determinadas células somáticas que se encuentran junto a la gónada en desarrollo hacia su interior, formando un tejido conjuntivo imprescindible para la producción normal de los espermatozoides. Figura 20-17 Reprogramación inducida por Sry en un embrión hembra de ratón para que se desarrolle como ratón macho. El gen Sry, inyectado al núcleo de un zigoto hembra XX, da lugar a un embrión transgénico que se desarrolla como un macho. Los genitales externos del ratón no se distinguen de los de un ratón macho normal XY. (De P. Koopman et al., Nature 351:117-121, 1991. © Macmillan Ediciones Omega 3 Biología molecular de LA CÉLULA – 4a. Edición LAS VENTAJAS DEL SEXO Magazines Ltd.) 4. Induciendo a otras células somáticas de la gónada en desarrollo a que se transformen en células de Leydig, secretoras de la hormona sexual masculina testosterona; dicha hormona es la responsable de la aparición de todos los caracteres sexuales secundarios de los machos. Entre ellos se incluyen las estructuras del tracto reproductor masculino, como la próstata y las vesículas seminales que se forman a partir de otro conducto, denominado sistema conductor de Wolffian. Este sistema degenera durante el desarrollo del sistema reproductor femenino porque para sobrevivir y desarrollarse requiere testosterona. La testosterona también masculiniza el desarrollo temprano del cerebro y por consiguiente el comportamiento: por ejemplo, ratas hembras recién nacidas tratadas con testosterona muestran, más tarde, un comportamiento sexual masculino. El gen Sry codifica una proteína reguladora de genes (Sry) que activa la transcripción de otras proteínas similares necesarias para el desarrollo de las células de Sertoli, como la proteína Sox9, relacionada con la proteína Sry. En ausencia de Sry o de Sox9, la cresta genital desarrolla un ovario. Las células de sostén se transforman en células foliculares en lugar de en células de Sertoli. Otras lo hacen en células de la teca en lugar de en células de Leydig y, al llegar a la pubertad, segregan estrógeno, la hormona sexual femenina, en lugar de testosterona. Las células germinales primordiales se diferencian en oocitos en vez de espermatozoides (Figs. 20-18 y 20-28), y el animal de desarrolla como una hembra. Figura 20-18 Influencia de Sry en el desarrollo gonadal. Las células de la línea germinal están coloreadas en rojo y las células somáticas en verde y azul. El cambio de color claro a oscuro indica que las células han madurado o se han diferenciado. El gen Sry actúa en una subpoblación de células somáticas durante el desarrollo gonadal induciendo que se diferencien en células de Sertoli o en células foliculares. Las células de Sertoli inducen a las células germinales primordiales a iniciar la espermatogénesis. Estas células también segregan la hormona anti Mülleriana que determina la regresión del conducto de Müller e induce a otras células somáticas a diferenciarse en células de Leydig, que son las células secretoras de testosterona (v. Fig. 20-28). En ausencia de Sry, las células germinales primordiales se implican en el desarrollo de los oocitos y las células somáticas se transforman o bien en células foliculares Ediciones Omega 4 Biología molecular de LA CÉLULA – 4a. Edición LAS VENTAJAS DEL SEXO que actúan como sostén del oocito durante su desarrollo o bien en células de la teca secretoras de estrógenos. Mientras que las células de Leydig empiezan la secreción de testosterona en el feto, las células de la teca no empiezan fe secreción de estrógenos hasta la pubertad. Si se eliminan las crestas genitales antes de que hayan empezado a diferenciarse en testículos u ovarios, un mamífero se desarrolla como una hembra independientemente de su sexo cromosómico. Parece ser que en los mamíferos la vía de desarrollo sexual "por defecto" es el desarrollo sexual femenino. OOCITOS Los oocitos son las células animales más extraordinarias que existen, al menos en un aspecto: una vez activados pueden dar lugar a un nuevo individuo completo en tan sólo cuestión de días o semanas. Ninguna otra célula de un animal superior tiene esta capacidad. Normalmente, la activación es la consecuencia de la fecundación -la fusión de un espermatozoide con un oocito-. Sin embargo, en algunos organismos el espermatozoide no es estrictamente necesario, de forma que un oocito puede ser activado artificialmente mediante diferentes tratamientos químicos o físicos no específicos. Efectivamente, algunos organismos, incluidos algunos vertebrados como determinadas especies de lagartija, normalmente se reproducen a partir de oocitos que se han activado en ausencia de espermatozoides, es decir de forma partenogénica. A pesar de que el oocito da lugar a todos los de un organismo, no es más que una célula altamente destinada exclusivamente a generar un nuevo tipos celulares especializada, individuo. Figura 20-19 Tamaño real de tres óvulos diferentes. El diámetro de de 0,1 mm. un óvulo humano es El citoplasma de un oocito puede llegar a reprogramar el núcleo de una célula somática, haciéndolo capaz de dirigir el desarrollo de un nuevo individuo. Ésta es la forma como se ha producido la famosa oveja Dolly. Se destruyó el núcleo de un oocito no fecundado de una oveja y se reemplazó por el núcleo de una célula somática adulta. Se activó el oocito mediante un shock eléctrico y el embrión resultante se implantó en el útero de una madre "de alquiler". La oveja resultante tenía el genoma del dador de la célula somática. Por lo tanto era un clónico de esta oveja dadora. En este apartado vamos a considerar brevemente algunas de las características especiales del oocito y después estudiaremos cómo se desarrolla hasta el momento que es apto para la fecundación. Ediciones Omega 5 Biología molecular de LA CÉLULA – 4a. Edición LAS VENTAJAS DEL SEXO Figura 20-20 Tamaños relativos de varios óvulos. Se compara con el tamaño de una célula somática característica. Un oocito es una célula altamente especializada con desarrollo independiente que contiene una gran cantidad de reservas nutritivas y una compleja cubierta Los oocitos de la mayoría de animales son células gigantes, con una gran cantidad de reservas de todos los nutrientes necesarios para el desarrollo inicial del embrión hasta que alcanza el estadio en el cual el nuevo individuo es capaz de alimentarse por sí mismo. Antes de alcanzar este punto, la célula gigante se divide en muchas células "más pequeñas, sin que se produzca crecimiento. El embrión de los mamíferos es una excepción. En este caso, el crecimiento del embrión se puede iniciar antes, ya que toma los nutrientes de la madre a través de la placenta. Por esta razón, los oocitos de mamífero, a pesar de ser células voluminosas, no necesitan serlo tanto como, por ejemplo, los de una rana o los de un ave. En general, los oocitos son esféricos u ovoides, con un diámetro aproximado de 0,1 mm los humanos y los de los erizos de mar (que son comestibles), de 1 mm a 2 mm los de anfibios y peces, y de varios centímetros los de aves y reptiles (Fig. 20-19). Por el contrario, normalmente las células somáticas tienen un diámetro de tan sólo entre 10 y 20 μm (Fig. 20-20). Figura 20-21 La cubierta vitelina. (A) Micrografía electrónica de barrido de un oocito de hámster, mostrando la cubierta vitelina. En (B) la zona (a la que están adheridos muchos espermatozoides) se ha separado para poner de manifiesto la membrana plasmática subyacente que presenta numerosos microvilli. La cubierta vitelina ha sido sintetizada exclusivamente por el oocito. (De D.M. Phillips J. Ultrastruct Res. 72:1-12, 1980.) El citoplasma de los oocitos contiene reservas de nutrientes en forma de vítelo, el cual es rico en lípidos, proteínas y polisacáridos y generalmente se encuentra formando unas estructuras específicas que se denominan plaquetas vitelinas. En algunas especies las plaquetas vitelinas presentan una membrana a su alrededor, pero en otras no. En los oocitos que se desarrollan fuera del cuerpo de la madre y dan lugar a organismos más o menos grandes, el vitelo puede constituir hasta el 95% del volumen de la célula. En los oocitos de los mamíferos, cuyos embriones son alimentados durante bastante tiempo por sus madres, hay muy poco vitelo si es que lo hay. La cubierta es otra de las características de los oocitos. Se trata de una especialización de la matriz extracelular que está formada, en gran parte, por glucoproteínas, Ediciones Omega 6 Biología molecular de LA CÉLULA – 4a. Edición LAS VENTAJAS DEL SEXO unas secretadas por el oocito y otras por las células acompañantes. En muchas especies, la cubierta principal es una capa adyacente a la membrana plasmática del oocito denominada cubierta vitelina (Fig. 20-21). Esta capa protege al oocito frente a agresiones mecánicas y en muchos casos también actúa como una barrera específica de especie para los espermatozoides, de forma que solamente admite los espermatozoides de la misma especie del oocito o de especies muy próximas. Muchos oocitos contienen vesículas secretoras especializadas situadas inmediatamente por debajo de la membrana plasmática en la región más periférica o córtex del citoplasma. Cuando un espermatozoide activa un oocito, se produce la exocitosis de dichos granulos corticales; el contenido liberado de los gránulos modifica la cubierta de tal manera que impide que se fusione otro espermatozoide (v. más adelante). Los gránulos corticales están distribuidos regularmente por todo el córtex del oocito, pero en algunos organismos existen otros componentes citoplasmáticos que presentan una sorprendente distribución asimétrica. Estos componentes permiten establecer, más tarde, la polaridad del embrión, como veremos en el Capítulo 21. Los oocitos se desarrollan por etapas Un oocito es un óvulo en desarrollo. La diferenciación de un oocito en un óvulo maduro supone una serie de cambios que se producen coordinados con las etapas de la meiosis, mediante la cual las células germinales experimentan dos divisiones extraordinariamente especializadas. Los oocitos han desarrollado mecanismos concretos para detener la meiosis: se mantienen en la profase I durante un período largo de tiempo mientras crecen en tamaño y en muchos casos se paran de nuevo en la metafase II, esperando la fecundación (aunque también pueden detenerse en otros momentos distintos, dependiendo de las especies de que se trate). Figura 20-22 Las etapas de la oogénesis. Las oogenias se forman a partir de las células germinales primordiales, que ai principio de la embriogénesis migran hacia la gónada en desarrollo. Tras un cierto número de divisiones mitóticas, las oogenias empiezan la división meiótica I, recibiendo entonces el nombre de oocitos primarios. En los mamíferos, los oocitos primarios se forman muy temprano Ediciones Omega 7 Biología molecular de LA CÉLULA – 4a. Edición LAS VENTAJAS DEL SEXO (entre los 3 y los 8 meses de gestación en el embrión humano) y permanecen en la profase de la división meiótica I hasta que la hembra es madura sexualmente. En este momento, periódicamente un pequeño número de oocitos maduran por influencia hormorai. completando la división meiótica I para convertirse en oocitos secundarios, los cuales finalmente experimentan la divisiéir meiótica II y se transforman en óvulos maduros. El momento en que el oocito es liberado del ovario y es fecundado varía de una especie a otra. En la mayoría de tos vertebrados la maduración de los oocitos está detenida en la metafase de la meiosis II y el oocito secundario sólo completa la meiosis II si es fecundado. Finalmente, los corpúsculos polares degeneran. En la mayoría de animales, el oocito en desarrollo está rodeado por células accesorias (células foliculares) especializadas que ayudan 2 aislarlo y a nutrirlo (no se muestran). Los detalles del desarrollo del oocito (oogénesis) varían según la especie. Sin embargo, las etapas principales son muy parecidas entre sí, como se muestra en la Figura 20-22. Las células germinales primordiales migran hacia la gónada en formación convirtiéndose en oogonias, que proliferan mediante sucesivas mitosis hasta diferenciarse en oocitos primarios. En este estadio (antes en aves y mamíferos), empieza la primera división meiótica: el DNA se replica de manera que cada cromosoma está formado por dos cromátidas hermanas, los cromosomas homólogos se aparean a lo largo de su eje longitudinal y se producen los entrecruzamientos entre cromátidas no hermanas de cada par de cromosomas. La célula se para en la profase de la división meiótica I (en un estadio equivalente a la fase G2 del ciclo mitótico, como hemos señalado antes) durante un período de tiempo que varía desde algunos días hasta varios años, según la especie. Durante este largo período (o en algunos casos, al llegar a la madurez sexual), los oocitos primarios sintetizan la cubierta vitelina y los gránulos corticales. En los grandes oocitos de animales no mamíferos, también se acumulan ribosomas, vitelo, glucógeno, lípidos y el mRNA que luego dirigirá la síntesis de las proteínas necesarias para el crecimiento embrionario temprano y la puesta en marcha del programa de desarrollo. En muchos oocitos, la gran actividad biosintética se refleja en la estructura de los cromosomas, que se descondensan y forman bucles laterales, tomando un aspecto "plumulado", que indica que están en síntesis activa de RNA (v. Figs. 4-36 y 4-37). Figura 4-36 Cromosomas plumulados. (A) Imagen de microscopía óptica de cromosomas plumulados de un oocito de anfibio. En una fase inicial de la diferenciación, cada cromosoma se replica al empezar la meiosis y los cromosomas homólogos replicados se aparean formando esta estructura muy extendida que contiene un total de cuatro moléculas de DNA replicadas o cromátidas. El estado de cromosomas plumulados puede persistir durante meses o años mientras el oocito elabora el aprovisionamiento de materiales que serán utilizados para su desarrollo en un nuevo individuo. (B) Imagen de microscopía de fluorescencia donde aparece una porción de un cromosoma plumulado de anfibio. Las regiones del cromosoma que se expresan activamente están marcadas en verde utilizando anticuerpos que reconocen una proteína que procesa el RNA durante una de las etapas de la expresión génica (v. Capítulo 6). Se supone que los granulos redondos corresponden a complejos de la maquinaria de maduración del RNA, los cuales trataremos en el Capítulo 6. (A, por cortesía de: Joseph G. Gall; B por cortesía de Joseph G. Gall y Cristine Murphy.) Ediciones Omega 8 Biología molecular de LA CÉLULA – 4a. Edición LAS VENTAJAS DEL SEXO Figura 4-37 Modelo de estructura de un cromosoma plumulado. El conjunto de cromosomas plumulados en muchos anfibios contiene un total de aproximadamente 10.000 bucles de cromatina diferertes, a pesar de que la mayor parte del DNA de cada cromosoma está muy coodensado en los cromómeros. Cada bucte corresponde a una determinada secuencia de DNA. Cada célula contiene cuatro copias de cada bucle, dado que cada une de los cromosomas consiste en dos cromátidas hermanas muy juntas, como se muestra en la parte superior del esquema. Esta estructura en cuatro hebras es muy característica de esta fase del desarrolo del oocito, la fase de diploteno de la meiosis; véase Figura 20-12. La fase siguiente de la oogénesis se denomina maduración de los oocitos. Generalmente no se produce hasta la madurez sexual, cuando el oocito es estimulado por hormonas. Por influencia hormonal, la célula reanuda la meiosis I. Los cromosomas vuelven a condensarse, sé rompe la envoltura nuclear (estos cambios marcan el inicio de la maduración) y en la anafase I los cromosomas homólogos replicados se segregan en dos núcleos hijos, cada uno de los cuales contiene la mitad del número inicial de cromosomas. Al finalizar la división I, el citoplasma se divide asimétricamente en dos, dando lugar a dos células de tamaño muy diferente: una es un pequeño corpúsculo polar y la otra es un gran oocito secundario, el precursor del óvulo. En esta etapa, cada cromosoma está formado por dos cromátidas hermanas. Estas cromátidas no se separan hasta la división meiótica II, al distribuirse en dos células, como se ha descrito anteriormente. Después de esta separación cromosómica final en la anafase II, el citoplasma del gran oocito secundario se vuelve a dividir asimétricamente produciendo el óvulo y un segundo corpúsculo polar, ambos con una dotación haploide de cromosomas (v. Fig. 20-22). A causa de estas dos divisiones asimétricas de su citoplasma, los óvulos mantienen su gran tamaño a pesar de experimentar dos divisiones meióticas. Los dos corpúsculos polares son pequeños y finalmente degeneran. En la mayoría de los vertebrados la oogénesis de desarrolla hasta la metafase de la meiosis II, momento en el que se detiene hasta la fecundación. En la oocitación el oocito secundario se desprende del ovario y experimenta una rápida maduración que lo transforma en un oocito apto para la fecundación. Si se produce la fecundación, el oocito es estimulado para que finalice la meiosis. Ediciones Omega 9 Biología molecular de LA CÉLULA – 4a. Edición LAS VENTAJAS DEL SEXO Los oocitos alcanzan su gran tamaño mediante mecanismos especiales de crecimiento Normalmente, una célula somática de un diámetro de entre 10 y 20 um necesita unas 24 horas para duplicar su masa como preparación para la división. A esta velocidad de biosíntesis, esta misma célula necesitaría mucho más tiempo para tener la masa un millón de veces superior de un oocito de insecto de 100 μm de diámetro. Sin embargo algunos insectos viven sólo unos días y lo consiguen. Es evidente que los oocitos necesitan mecanismos especiales para alcanzar su gran tamaño. Una estrategia sencilla para crecer rápidamente consiste en disponer de copias extra de genes. Para ello el oocito retrasa el final de la primera división meiótica, es decir, crece mientras su dotación cromosómica diploide está duplicada. De esta forma dispone de doble cantidad de DNA para la síntesis de RNA en comparación con una célula somática en fase G1 del ciclo celular. Los oocitos de algunas especies también alcanzan grandes tamaños acumulando DNA extra: producen muchas copias extras de determinados genes. En el Capítulo 6 hemos visto que las células somáticas de la mayoría de organismos necesitan entre 100 y 500 copias de los genes que codifican el RNA ribosómico para producir suficiente cantidad de ribosomas para la síntesis proteica. Los oocitos necesitan un número mayor de ribosomas para atender a la síntesis proteica en los inicios de la embriogénesis, por lo que en los oocitos de muchos animales los genes del RNA ribosómico están amplificados; por ejemplo, los oocitos de algunos anfibios contienen entre 1 y 2 millones de copias de estos genes. Los oocitos también dependen para su crecimiento de la actividad sintetizadora de otras células. El vitelo, por ejemplo, se sintetiza habitualmente fuera del ovario y luego el oocito lo incorpora. En aves, anfibios e insectos, las proteínas del vitelo se sintetizan en el hígado y son vertidas a la sangre. En los ovarios, los oocitos importan las proteínas vitelínicas de la matriz extracelular por un mecanismo de endocitosis mediada por receptor (v. Fig. 13-41). Figura 13-41 Formación de vesículas revestidas de clatrina a partir de la membrana plasmática. Estas micrografías electrónicas ilustran la secuencia probable de acontecimientos que se producen durante la formación de una vesícula revestida de clatrina a partir de una depresión revestida de clatrina. Las depresiones y las vesículas revestidas que aparecen en las fotografías son mucho mayores que las que se presentan en las células de tamaño normal. Intervienen en la captación de lipoproteínas en un gran oocito de gallina, formando la yema del huevo. En la superficie extracelular de la membrana plasmática -que se corresponde con la superficie interior de la vesícula- puede verse una capa electrodensa que corresponde a las lipoproteínas asociadas a su receptores. (Por cortesía de M.M. Perry y A.B. Gilbert, de J. Cell Sci. 39:257-272, 1979. ©The Company of Biologists.) Ediciones Omega 10 Biología molecular de LA CÉLULA – 4a. Edición LAS VENTAJAS DEL SEXO Figura 20-23 Células nodriza y células foliculares asociadas a un oocito de Drosophíla. Las células nodriza y el oocito proceden de una misma oogenia, que da lugar a un oocito y a 15 células nodriza (de las cuales en esta figura se observan solamente 7). Estas células permanecen unidas por puentes citoplasmáticos, resultantes de una división celular incompleta. Finalmente, las células nodriza descargan el contenido de su citoplasma en el oocito y se autodestruyen. Las células foliculares se desarrollan independientemente (a partir de las células del mesodermo). Los nutrientes también pueden proceder de las células accesorias del ovario. Las hay de dos tipos. En algunos invertebrados, determinadas células de la progenie de las oogonias se transforman en células nodriza y no en oocitos. Estas células generalmente están conectadas al oocito por puentes citoplasmáticos a través de los cuales pueden pasar las macromoléculas directamente al oocito (Fig. 20-23). En los oocitos de los insectos, sintetizan muchos de los productos -ribosomas, mRNA, proteínas y otras moléculas- que los oocitos de los vertebrados tienen que sintetizar ellos mismos. Otras células accesorias del ovario que colaboran en la nutrición de los oocitos en desarrollo son las células foliculares, presentes en invertebrados y vertebrados. Figura 20-24 Micrografía electrónica de oocitos primarios en desarrollo en el ovario de una hembra de conejo. (A) Estadio inicial de un oocito primario en desarrollo. Todavía no se han formado ni la cubierta vitelina ni los gránulos corticales y el oocito está rodeado por una monocapa de células foliculares planas. (B) Oocito primario más maduro, observado a un aumento seis veces menor, debido a que es mucho más grande que el oocito de (A). Este oocito presenta una delgada cubierta vitelina y está rodeado por varias capas de células foliculares y una lámina basal que lo aislan de las otras células del ovario. El oocito primario, junto con las células foliculares que lo rodean, forma un folículo primario. Las células foliculares están conectadas entre sí y con el oocito mediante uniones de Ediciones Omega 11 Biología molecular de LA CÉLULA – 4a. Edición LAS VENTAJAS DEL SEXO tipo gap. (De The Cellular Basis of Mammalian Reproduction [J. Van Blerkom y R Motta eds.]. Baltimore-Munich: Urban & Schwarzenberg, 1979.) Se disponen como una capa de células epiteliales alrededor del oocito (Fig. 20-24 y v. Fig. 20-23), al que están conectadas mediante uniones de tipo gap, que permiten el intercambio de moléculas pequeñas pero no de macromoléculas. Estas células no pueden proporcionar macromoléculas al oocito a través de estas uniones, pero pueden colaborar suministrando los precursores, de menor tamaño, a partir de los cuales se sintetizan. Además, a menudo las células foliculares secretan macromoléculas que contribuyen a la formación de las cubiertas del oocito o son incorporadas por el oocito en crecimiento mediante endocitosis o actúan como receptores de la superficie celular del oocito controlando el patrón espacial y las asimetrías axiales de los oocitos (v. Capítulo 21). ESPERMATOZOIDES En la mayoría de especies existen dos tipos de gametos muy distintos: el oocito, una de las células más voluminosas del organismo; y el espermatozoide, probablemente más pequeña. Ambos están optimizados en sentidos diferentes para la transmisión de los genes que transportan. El oocito es una célula inmóvil que asegura la supervivencia de los genes maternos mediante una gran cantidad de materia prima para el crecimiento y el desarrollo, y una eficaz cubierta protectora. El espermatozoide está preparado para la propagación de los genes paternos utilizando las reservas maternas: es una célula muy móvil e hidrodinámica, lo que le proporciona velocidad y eficacia para la fecundación. La competencia entre los espermatozoides es intensa y la mayoría fracasan en su misión: de los miles de millones que son liberados en el período fértil de la vida de un hombre, sólo unos cuantos fecundarán un oocito. Los espermatozoides están extraordinariamente especializados en transmitir su DNA a un oocito En general, los espermatozoides son células "despojadas", con un potente flagelo que les da impulso para desplazarse en un medio acuoso, pero sin organillos citoplasmáticos como ríbosomas, retículo endoplasmático, o complejo de Golgi, orgánulos, que no necesitan para transmitir el DNA al oocito. Sin embargo, contienen muchas mitocondrias colocadas estratégicamente donde pueden proporcionar, con la mayor eficacia, la energía que necesita el flagelo para su movimiento. Un espermatozoide está formado normalmente por dos regiones, morfológica y funcionalmente distintas, limitadas por una membrana plasmática única. Estas regiones son la cola, que impulsa el espermatozoide hacia el oocito y le ayuda a atravesar la cubierta oocitaria, y la cabeza, donde se encuentra el núcleo haploide (Fig. 20-25). El DNA del núcleo está fuertemente empaquetado, de modo que su volumen queda reducido al mínimo facilitando su transporte y no es posible la transcripción. En muchas especies los cromosomas de los espermatozoides carecen de las histonas que presentan las células somáticas y en su lugar están empaquetados con unas proteínas de elevada carga positiva llamadas protaminas. En la cabeza de los espermatozoides de muchos animales existe una vesícula secretora muy especializada en estrecha aposición con la parte anterior de la envoltura nuclear llamada vesícula acrosómica (v. Fig. 20-25). Esta vesícula contiene enzimas hidrolíticas que ayudan a atravesar la envoltura externa del oocito. Cuando un espermatozoide entra en contacto con un oocito, el contenido de la vesícula se libera por Ediciones Omega 12 Biología molecular de LA CÉLULA – 4a. Edición LAS VENTAJAS DEL SEXO exocitosis, la denominada reacción acrosómica; en algunos espermatozoides, esta reacción también deja al descubierto o libera proteínas que colaboran específicamente en la unión del espermatozoide con la cubierta del oocito. La cola móvil de un espermatozoide es un largo flagelo, cuyo axonema emerge de un corpúsculo basal situado justo por debajo del núcleo. Como se describe en el Capítulo 16, el axonema está formado por dos microtúbulos centrales rodeados por nueve dobletes de microtúbulos dispuestos ordenadamente. El flagelo de algunos espermatozoides (incluido el caso de los mamíferos) se diferencia de los otros flagelos en que el habitual patrón 9 + 2 del axonema está rodeado además por nueve fibras densas externas (Fig. 20-26). Se desconoce el papel que desempeñan estas fibras densas y no contráctiles en el movimiento del flagelo, que se produce por el deslizamiento entre los dobletes adyacentes de microtúbulos. El movimiento flagelar está impulsado por la dineína, proteína motora de los microtúbulos, que utiliza la energía de la hidrólisis del ATP para el deslizamiento de los microtúbulos, como se ha visto en el Capítulo 16. El ATP lo generan las mitocondrias extraordinariamente especializadas de la parte anterior de la cola del espermatozoide (denominada pieza, intermedia], que es el lugar donde se necesitan (v. Figs. 20-25 y 20-26). En la mayoría de mamíferos, los espermatozoides se producen de manera continuada En los mamíferos, existen diferencias importantes en la manera en que se producen los oocitos (oogénesis) y los espermatozoides (espermatogénesis). En las mujeres, las oogonias proliferan exclusivamente en el feto, entran en meiosis antes del nacimiento y se detienen, transformadas en oocitos, en la profase de la meiosis I, estado en el cual se pueden mantener durante más de 50 años. Los oocitos maduran a partir de este limitado stock y ovulan a intervalos, generalmente empezando en la pubertad. En cambio, en el hombre la meiosis y la espermatogénesis no empiezan hasta la pubertad y desde entonces se producen de manera continua Figura 20-25 Espermatozoide humano. en el epitelio que forma la pared de unos Esquema de la sección longitudinal largos y enrollados tubos del testículo, los túbulos seminíferos. Las células germinales las espermatogonias, se localizan alrededor lámina basal, donde proliferan del límite externo de estos túbulos junto a la continuamente por mitosis. Ediciones Omega 13 Biología molecular de LA CÉLULA – 4a. Edición LAS VENTAJAS DEL SEXO Figura 20-26 Esquema de la sección transversal de la pieza media de un espermatozoide de mamífero. El centro del flagelo está formado por un axonema rodeado por nueve fibras densas. Éste comprende dos microtúbulos sencillos rodeados por nueve dobletes de microtúbulos. Las mitocondrias (verde) están bien situadas para proporcionar el ATP necesario para el movimiento flagelar; su disposición en espiral (v. Fig. 20-25) se debe a la fusión de pequeñas mitocondrias durante la diferenciación de las espermátidas. Algunas de las células hijas paran la proliferación y se diferencian en espermatocitos primarios. Éstos entran en la profase meiótica, en la que se aparean los cromosomas homólogos y se producen los entrecruzamientos, y después continúan hasta terminar la división meiótica I, dando lugar a dos espermatocitos secundarios, cada uno de los cuales contiene 22 cromosomas autosómicos duplicados y un cromosoma X o Y duplicado. Los dos espermatocitos secundarios procedentes de un espermatocito primario entran en la segunda división meiótica, produciendo cuatro espermátidas, cada una de las cuales tiene una dotación cromosómica haploide. Las espermátidas experimentan una diferenciación morfológica que las transforma en espermatozoides (Fig. 20-27), los cuales son liberados a la luz del túbulo seminífero (Fig. 20-28). A continuación los espermatozoides pasan por el epidídimo, un tubo muy sinuoso que se encuentra pegado a los testículos, donde son almacenados y donde se produce su maduración final. Un rasgo interesante de la espermatogénesis es que las células germinales masculinas en desarrollo no completan la división citoplasmática (citocinesis) ni durante la mitosis ni durante la meiosis. Por consiguiente, grandes clones de células hijas diferenciadas descendientes de una espermatogenia madura permanecen conectados por puentes citoplasmáticos, formando un sincitio (Fig. 20-29). Los puentes citoplasmáticos se mantienen hasta el final de la diferenciación de los espermatozoides, cuando éstos son liberados a la luz del túbulo. Ello explica el hecho de que los espermatozoides maduros aparezcan sincrónicamente en cualquier área del túbulo seminífero. Sin embargo, ¿cuál es la función de esta organización sincitial? Ediciones Omega 14 Biología molecular de LA CÉLULA – 4a. Edición LAS VENTAJAS DEL SEXO Al contrario que los oocitos, los espermatozoides experimentan la mayor parte de su diferenciación después de que el núcleo haya completado la meiosis, siendo por lo tanto células haploides. Sin embargo, la presencia de puentes citoplasmáticos entre ellos significa que cada espermatozoide haploide en desarrollo comparte un citoplasma común con sus vecinos. Así, puede disponer de todos los productos de un genoma diploide completo. Por ejemplo, el espermatozoide en desarrolIo que transporta el cromosoma. Y, puede disponer de las proteínas esenciales codificadas por los genes del cromosoma X. Así pues, el genoma diploide dirige la diferenciación de los espermatozoides de igual mañera que en la diferenciación de los oocitos. Figura 20-27 Etapas de la espermatogénesis. Las espermatogonias se forman a partir de las células germinales primordiales que migran al testículo en las primeras etapas de la embriogénesis. Cuando el animal alcanza la madurez sexual, las espermatogonias empiezan a proliferar rápidamente, generando una línea de células que mantienen la capacidad de dividirse continua e indefinidamente (como células madre de las espermatogonias) y otra línea celular (espermatogonias en maduración) que, después de un número limitado de mitosis, entra en la meiosis transformándose en espermatocitos primarios. Los espermatocitos primarios continúan la división meiótica I hasta convertirse en espermatocitos secundarios. Completada la división meiótica II, los espermatocitos secundarios producen las espermátidas haploides, que se diferencian en espermatozoides maduros. La espermatogénesis se diferencia de la oogénesis (v. Fig. 20-22) en varios aspectos. (I) A partir de la pubertad, continuamente se producen nuevas células que inician la meiosis. (2) Cada célula que empieza la meiosis da lugar a cuatro gametos maduros y no a uno solo. (3) Los espermatozoides maduros se forman mediante un elaborado proceso de diferenciación celular, que empieza cuando acaba la meiosis. (4) Se dan aproximadamente dos veces más divisiones en la producción de un espermatozoide que en la producción de un oocito. En el ratón, por ejemplo, se ha calculado que por término medio se producen unas 56 divisiones entre el zigoto y el espermatozoide maduro y alrededor de 27 entre el zigoto y el oocito maduro. Ediciones Omega 15 Biología molecular de LA CÉLULA – 4a. Edición LAS VENTAJAS DEL SEXO Figura 20-28 Esquema muy simplificado de la sección transversal de un túbulo seminífero de testículo de mamífero. (A) Todas las fases de la espermatogénesis que se muestran tienen lugar mientras los gametos en desarrollo están en íntima asociación con las células de Sertoli. Estas grandes células se extienden desde la lámina basal hasta el lumen del túbulo seminífero; son necesarias para la supervivencia de las células germinales y son análogas a las células foliculares del ovario (v. Fig. 20-18). La espermatogénesis también depende de la testosterona secretada por las células de Leydig situadas entre los túbulo seminíferos. (B) Las espermatogenias en división se encuentran apoyadas sobre la lámina basal. Algunas de ellas se autorrenuevan como células madre de las espermatogenias, mientras que otras se transforman en espermatogenias maduras. Después de un determinado número de mitosis, las espermatogenias maduras frenan las mitosis y entran en la meiosis transformándose en espermatocitos primarios. Posteriormente, los espermatozoides son liberados al lumen del túbulo. En el hombre, tienen que pasar 24 días para que un espermatocito complete la meiosis y se transforme en espermátida, y otras 5 semanas para que una espermátida se convierta en un espermatozoide. Los espermatozoides experimentan una maduración posterior y se hacen móviles en el epidídimo; sólo entonces son espermatozoides completamente maduros. Algunos de los genes que regulan la espermatogénesis se han conservado a lo largo de la evolución desde las moscas hasta los seres humanos. El gen DAZ, por ejemplo, que codifica una proteína de unión a RNA y que se localiza en el cromosoma Y, falta en muchos hombres estériles, la mayoría de los cuales no pueden producir espermatozoides. Dos genes de Drosophila homólogos a DAZ son esenciales para la espermatogénesis en esta especie. Las proteínas de unión a RNA son especialmente importantes en la espermatogénesis porque muchos de los genes expresados en la línea espermática están regulados a nivel de la traducción del RNA. Ediciones Omega 16 Biología molecular de LA CÉLULA – 4a. Edición LAS VENTAJAS DEL SEXO Figura 20-29 Puentes citoplasmáticos entre los espermatozoides en desarrollo y sus precursores. Generalmente, las células descendientes de una espermatogenia madura permanecen conectadas entre sí por puentes citoplasma-ticos durante su diferenciación hasta espermatozoides maduros. Para una mejor comprensión sólo se muestra la entrada en la meiosis de dos espermatogenias maduras unidas, que producen ocho espermátidas haploides también unidas. En realidad, el número de células conectadas que pasan por las dos divisiones meióticas y se diferencian sincrónicamente es mucho mayor del que se representa aquí. Es interesante señalar que, en el proceso de diferenciación, la mayor parte del citoplasma de las espermátidas se separa en forma de cuerpo residual. Ediciones Omega 17