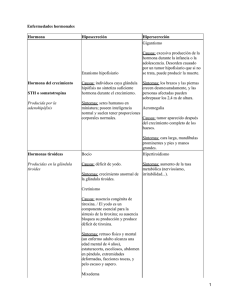
INDICE 1.− INTRODUCCIÓN 2.− LAS HORMONAS 2.1.− HORMONAS PROTEICAS
Anuncio

INDICE 1.− INTRODUCCIÓN 2.− LAS HORMONAS 2.1.− HORMONAS PROTEICAS 2.2.− HORMONAS ESTEROIDEAS 2.3.− GIBERELINAS 3.− GLÁNDULAS ENDOCRINAS DEL HOMBRE 3.1.− TIROIDES 3.2.− PARATIROIDES 3.3.−PANCREAS ENDOCRINO 3.4.− GLÁNDULAS ADRENALES 3.5.− HIPÓFISIS 3.6.− GÓNADAS 3.7.− GLÁNDULA PINEAL 4.− ORGANOS DIANA 5.− ANORMALIDADES ENDOCRINAS 5.1.− MIXEDEMA 5.2.− BOCIO EXOFTÁLMICO 5.3.− TETANIO 5.4.− DIABETES 5.5.− ENFERMEDAD DE ADDISSON 5.6.− DIABETES INSÍPIDA 5.7.− HIPO e HIPERTIROIDISMO 6.− DIFERENTES FUNCIONES DE LAS HORMONAS 6.1.− TRANSMISORES DE INFORMACIÓN 1 6.2.− CONTROLES DE LÍMITES 6.3.− CONTROLES DE RETROALIMENTACIÓN RECÍPROCOS O NEGATIVOS 7.− COMPLEJOS SISTEMAS DE CONTROL 7.1.− REGULACIÓN DE LOS CICLOS ESTRUAL Y MENSTRUAL 7.2.− REGULACIÓN DEL DESARROLLO Y FUNCIÓN DE LAS GLÁNDULAS MAMARIAS 7.3.− REGULACIÓN HORMONAL DE LAS INTENSIDADES METABÓLICAS 8.− MECANISMOS DE ACCIÓN DE LAS HORMONAS 9.− FEROMONAS INTRODUCCIÓN Las actividades de las diversas partes del cuerpo de los animales superiores están integradas por dos dispositivos de coordinación, los sistemas nervioso y endocrino. Las rápidas respuestas de músculos y glándulas, contadas en milisegundos, están bajo el control del sistema nervioso. Las glándulas del sistema endocrino segregan hormonas, que son transportadas por la sangre hasta todo punto en que su presencia sea necesaria para ejercer sus funciones, regular las actividades de las células. Las respuestas provocadas por las hormonas son, en general, algo más lentas (medidas en minutos, horas o incluso semanas) pero su duración es también mayor comparado con la respuesta del sistema nervioso. Las adaptaciones a largo plazo del metabolismo, el crecimiento o la reproducción son, entre otras, las funciones típicas del sistema endocrino. Las glándulas endocrinas se distinguen por la particularidad de que secretan sus productos y los vierten en la corriente sanguínea, de modo que carecen en general de tubo excretor que lleve el producto activo al exterior o a uno de los órganos internos: por este motivo, se ha llamado también a estas glándulas de secreción interna. Unas, como tiroides, paratiroides, hipófisis y suprarrenales, secretan exclusivamente hormonas y no tienen conducto excretor; otras, como el páncreas, ovarios y testículos, poseen ambas secreciones, externa, a través de conductos, e interna, transportada por la corriente sanguínea. El páncreas, en verdad una glándula funcionalmente doble, aunque reunida en un mismo órgano, elabora enzimas digestivas a la vez que hormonas. En ciertos animales inferiores, las dos porciones constituyentes del páncreas están anatómicamente separadas. El conocimiento práctico de la endocrinología (ejemplificado en la castración de hombres y animales) ha existido durante miles de años, pero la endocrinología moderna se supone haber nacido en 1849. Berthold, como consecuencia de experimentos de transplante de testículos en las aves, postuló que estas glándulas sexuales masculinas secretan alguna substancia, acarreada por la sangre, esencial en la discriminación de los caracteres sexuales secundarios. La primera tentativa de tratamiento endocrino se hizo en 1889, por Brown−séquard, fisiólogo francés, quién se inyectó extractos testiculares, de los que pretendió haber extraído resultados rejuvenecedores. No se pudo comprobar si sus extractos pudieron tener otro efecto que el psicológico, pero lo positivo es que sus afirmaciones estimularon muchos trabajos de investigación. Para determinar si una glándula sospechosa de producir una hormona es en realidad una glándula endocrina, un investigador comienza generalmente extirpando quirúrgicamente la glándula y observando el efecto producido en el animal. Luego, sustituye la glándula por otra trasplantada de otro animal y determina si los cambios inducidos extirpando la glándula pueden invertirse sustituyéndola. Podría tratar después de alimentar luego a un animal al que se haya extraído la glándula con glándulas desecadas para determinar si la sustancia activa, la hormona, puede ser sustituida de este modo. Finalmente, obtiene un extracto de la glándula con una 2 variedad de disolventes para determinar las características de solubilidad del material activo y obtener así un indicio sobre su naturaleza química. Haciendo un extracto de la glándula, o quizá un extracto de la sangre u orina, y purificándolo mediante procesos químicos y físicos, obtiene finalmente el compuesto puro y puede extraer su estructura química exacta. Las hormonas son substancias extraordinariamente eficaces y en cantidades muy pequeñas producen un marcado efecto en la estructura o la función de una u otra parte del cuerpo: solo pequeñas cantidades de una hormona son secretadas en un momento dado por la glándula endocrina y la cantidad que circula por el cuerpo es muy pequeña. A causa de esto, el aislamiento de una hormona pura puede ser una tarea verdaderamente difícil. por ejemplo, para obtener unos pocos miligramos de estradiol puro, una hormona sexual femenina, ¡se necesitó un extracto de más de dos toneladas de ovarios de cerda!. Cuando la hormona secretada por una glándula dada ha sido extraída y se ha determinado su estructura química, puede prepararse entonces sintéticamente para ser usada en tratamiento de enfermedades resultantes de una deficiencia de dicha hormona. LAS HORMONAS El término "hormona" es un término practico, no un término químico; designa una característica funcional de la sustancia, en vez de cierta clase particular de compuestos químicos. En la definición clásica de E.H. Starling: "Una hormona es una substancia secretada por células de una parte del cuerpo que pasa a otra parte, donde actúa en muy pequeña concentración regulando el crecimiento o actividad de las células". Las hormonas generalmente son transportadas en la sangre desde su lugar de producción hasta el lugar donde actúan, pero los neurohumores pueden pasar por un axón y son transferidas prostaglandinas en el líquido seminal: las hormonas desempeñan un papel integrador en los vertebrados, los insectos, los crustáceos y las plantas. otros animales pueden tener sistemas endocrinos también y, de hecho, se han aislado hormonas sexuales en muchos animales inferiores. Las hormonas son, junto a las enzimas y las vitaminas, biocatalizadores o principios inmediatos en baja proporción. Estos, facilitan en general la materialización de los distintos procesos biológicos sobre los que intervienen, a pesar de que se requieren en escasa cuantía. En concreto, las hormonas son biocatalizadores endógenos, puesto que se requieren en pequeñas cantidades que pueden ser secretadas con facilidad por el propio organismo. Las hormonas no son específicas, sino que cada especie tiene sus propias hormonas concretas e incluso cada individuo dentro de su propia especie puede poseer diferentes hormonas que la de otro. Otra característica de las hormonas es que funcionan en sistema de retroalimentación negativa: el nivel más simple de retroalimentación se produce cuando ésta se produce entre las células diana y una determinada glándula endocrina. En la mayoría de los casos la retroalimentación se produce con la intervención de la hipófisis o del hipotálamo. Algunas hormonas pueden ser liberadas por las neuronas, funcionando entonces como neurotransmisores, pero la diferencia estriba en que mientras la neurotransmisión es más rápida y se produce en el mismo lugar (sinapsis), la neurosecrección hormonal es más lenta y produce sus efectos en una zona más o menos distante del lugar de secreción. Las hormonas pueden ser secretadas por glándulas endocrinas o exocrinas. Las glándulas exocrinas liberan su producto en el mismo sitio donde se han producido, y lo hacen a través de conductos específicos, y no a través del torrente sanguíneo, por lo que se las suele denominar glándulas "de conducto". Las principales glándulas exocrinas son el páncreas, como glándula mixta, las glándulas sudoríparas, salivares o sebáceas. Las glándulas endocrinas se localizan en nuestro cerebro y en algunas otras partes de nuestro organismo. Algunos órganos no específicamente endocrinos segregan sus propias hormonas, como la mucosa del duodeno, que produce tres hormonas (secretina, colecistoquinina y enterogastrona) y la mucosa pilórica del estómago, que produce gastrina. 3 La variedad de estructuras químicas que exhiben actividad hormonal es notable. Algunas hormonas son aminoácidos o derivados de aminoácidos (tiroxina, adrenalina, melatonina, ácido indolacético); otras son purinas (citocina), derivados de ácidos grasos (prostaglandinas, hormona juvenil), péptidos de cadena corta (oxitocina, hormona antidiurética), péptidos de cadena larga (ACTH, glucagón, insulina), proteínas (gonadotropinas, hormona del crecimiento), esteroides (hormonas sexuales, hormonas adrenocorticales, ecdisona) o giberelinas (hormonas del crecimiento de las plantas). No parece probable que todos estos diversos compuestos afecten a la función celular por el mismo mecanismo o por mecanismos similares. Ciertamente, es evidente que muchas hormonas tienen mecanismos de acción independientes mediante los cuales regulan las actividades celulares. Derivados proteicos. La adrenalina y la noradrenalina son aminas derivadas por descarboxilación de la tirosina. Son sintetizadas y almacenadas en las células cromafin de la médula adrenal y liberadas cuando se estimulan las células por impulsos nerviosos transmitidos por el sistema nervioso simpático. La noradrenalina es también un neurotransmisor producido por las puntas de los axones de nervios adrenérgicos y transmite el impulso a neuronas adyacentes Son, en última instancia, junto a la dopamina, el grupo constituyente de las catecolaminas. Estas dos hormonas reciben su nombre por el sitio dónde son producidas (las glándulas adrenales). La epinefrina también puede producirse en las neuronas, aunque esta producción tiene menos importancia que la de la norepinefrina. La función de este neurotransmisor radica en el control de la vigilia y el estado de alerta; también se ha demostrado su efecto en la memoria y el aprendizaje. Además de hormona y neurotransmisor, la noradrenalina tiene la función de neuromodulador, aumentando o disminuyendo la acción de otras neuronas adrenérgicas sobre las que actúa. La tiroxina, otro derivado de la tirosina, es sintetizada en la glándula tiroides. La tiroides tiene una bomba de yoduro notablemente eficaz que puede acumular yoduro del torrente sanguíneo y concentrarlo muchas veces. Los residuos de tirosina son yodados formando diyodotirosina y dos de estos se acoplan formando tiroxina. Se produce yodación de la tirosina y unión de las tirosinas yodadas cuando los aminoácidos son parte de una gran molécula de proteína, la tiroglobulina. La tiroxina contiene cuatro átomos de yodo y el compuesto relacionado, triyodotironina, que es también hormonalmente activo, tiene tres átomos de yodo. Una hormona secretada por el lóbulo anterior de la hipófisis, la tirotropina (TSH), refuerza todas las reacciones que intervienen en la producción de tiroxina −la absorción de yodo, la adición de yodo a la tirosina y la liberación de tirosina y la liberación de tiroxina a partir de la tiroglobulina. La tiroxina, en general, aumenta el metabolismo basal. El ácido indolacético, la hormona primaria del crecimiento o auxina de las plantas, es sintetizado a partir del aminoácido triptófano por transaminación y descarboxilación. En cualquier momento, solo hay cantidades muy pequeñas de auxina en las plantas pues es convertida enzimáticamente en le compuesto inactivo indolformaldehido. Una segunda hormona del crecimiento de las plantas, llamada citocinina, es un derivado de la adenina. Tiene una cadena isoprenoide de cinco carbonos ligada al grupo amino en el carbono 6 del anillo purina. La zeatina, citocinina hallada en semillas de maíz jóvenes, es purina 6. Una purina muy similar a esta es un constituyente del ARN de transferencia de serina y el ARN de transferencia de tirosina. Esta es una purina 6 que difiere de la zeatina sólo en que carece de un grupo hidroxilo en uno de los grupos metilo de la cadena lateral. Las gonadotropinas, la tirotropina y la hormona del crecimiento, secretadas por la hipófisis anterior, son verdaderas proteínas con un peso molecular de 25.000 o más. Las hormonas del crecimiento de diferentes mamíferos difieren notablemente en peso molecular. El hombre y otros primates tienen hormonas de crecimiento con pesos moleculares de 25.000 o más, mientras que en ovejas y bueyes tienen un peso molecular de 45.000 o más. La hormona del crecimiento en estos animales no estimulará el crecimiento en humanos, ya que −como antes se dijo− las hormonas son únicas para cada especie animal e incluso dentro de la misma especie, las hormonas sean diferentes e incompatibles; sin embargo, hay excepciones, y las hormonas de crecimiento humanas y vacunas si estimularán el crecimiento en ratas. 4 La hormona del crecimiento humana consta de 188 aminoácidos en una sola cadena. Hasta 10% de peso seco de la hipófisis humana puede ser hormona del crecimiento. Es curioso que un hombre de 80 años tiene tanta hormona de crecimiento en su hipófisis como un niño en pleno crecimiento. La hormona del crecimiento es rápidamente degradada tras haber sido secretada en el plasma; su periodo de semidesintegración biológica es de 25 minutos. La tirotropina o TSH es una glucoproteina básica con peso molecular de 25000. Contiene varias clases de carbohidratos: N−acetil galactosamina, manosa y fucosa. Es segregada por la hipófisis anterior que estimula el tiroides y la producción de tiroxina. Las gonadotropinas, o sea, hormona folículoestimulante, FSH, y la hormona luteinizante, LH, secretadas por el lóbulo anterior de la hipófisis también, son glucoproteinas. La FSH, con peso molecular de 31.000, contiene 8% de carbohidrato, incluyendo ácido siálico. La FSH pierde su actividad biológica −que es estimular la formación del folículo de Graff del ovario y de los túbulos seminíferos del testículo− cuando el ácido siálico es desdoblado por la enzima neuraminidasa o cuando los otros componentes del carbohidrato son eliminados por tratamiento con amilasa. La LH es una proteína ligeramente menor, con un peso molecular de 26.000. Las LH de diferentes especies tienen contenido de carbohidrato que varía desde el 4,5% en la LH humana hasta 11% en la de oveja. La LH regula la producción y liberación de estrógenos y progesterona por el ovario y testosterona por el testículo. Una tercera gonadotropina, la prolactina u hormona luteotrópica, es una proteína compuesta de una sola cadena de péptidos de 205 aminoácidos con peso molecular de 25.000. Esta hormona mantiene la secreción de estrógenos y progesterona por el ovario; estimula la producción de leche y, en cuanto a la conducta, influye en el "instinto materno". Las tres hormonas hipofisiarias de glucoproteina −TSH, FSH y LH− se componen de dos subunidades denominadas ð y ð. Las subunidades ð de las tres son muy similares, mientras que las ð son distintas y a ellas se debe la especificidad biológica de la hormona. Las hormonas pueden separarse en subunidades, que tienen poca o ninguna actividad biológica, y luego combinarse de nuevo para dar actividad total. Una puede combinar una subunidad ð de TSH con una subunidad ð de FSH y dar una proteína con actividad FSH. Hormonas esteroideas. Las prostaglandinas, derivados de ácidos grasos insaturados de 20 carbonos, tienen un anillo de cinco carbonos en medio de la cadena y grupos hidroxilo o cetona en ciertos carbonos de la cadena. Son secretados por la vesícula seminal y transportados con el semen al conducto reproductor femenino, donde regulan la actividad de los músculos uterinos. Son sintetizados a partir de ácidos grasos polinsaturados como el arachidónico, un ácido graso de 20 carbonos con cuatro enlaces dobles. Las prostaglandinas evidentemente afectan a los músculos lisos en otros tejidos distintos del útero y son secretadas por tejidos distintos de la vesícula seminal. Se les ha atribuido el control de una multitud de hechos. Péptidos de cadena corta. La óxitocina y la vasopresina u hormona antidiurética son péptidos cortos, compuestos de nueve aminoácidos, cinco de los cuales son idénticos en las dos hormonas. Las dos substancias poseen propiedades fisiológicas muy diferentes, aunque solo difieren en dos aminoácidos. Ambas son sintetizadas en las células neurosecretoras de los núcleos supraóptico y supraventricular del hipotálamo y luego pasan por los axones de estas células al lóbulo posterior de la hipófisis, dónde son almacenadas y posteriormente liberadas para que cumplan su función. La oxitocina estimula las contracciones de los músculos uterinos y la secrección de leche. La vasopresina u hormona antidiurética estimula la contracción de los músculos lisos y tiene una acción −como su nombre indica− antidiurética sobre los túbulos del riñón. La hormona estimulante de los melanocitos (MSH), un péptido que contiene 13 aminoácidos, es secretado por el lóbulo intermedio de la hipófisis. En ranas y otros anfibios, la MSH parece desempeñar una función en el oscurecimiento de la piel, haciendo que se expandan los melanocitos de la piel. La MSH en el hombre no está claro, pero la hipófisis humana produce MSH y la piel responde lentamente a ella. 5 Peptidos de cadena larga. Insulina, glucagón, ACTH y calcitonina son péptidos de cadena larga, con 30 aminoácidos en cada cadena. La insulina, secretada por las células ð del páncreas, aumenta la utilización de la glucosa por el músculo y otros tejidos; reduce la concentración de azúcar en sangre y aumenta los depósitos del glucógeno y el metabolismo de la glucosa. Consta de dos cadenas de péptidos unidos por enlaces de disulfuros. Una cadena contiene 21 aminoácidos y la otra 30. La insulina es producida en el páncreas como una sola cadena de péptidos compuesta de 84 aminoácidos. Esta, denominada proinsulina, experimenta plegamiento, formándose tres enlaces de disulfuro, y una cadena de péptidos que contiene 33 aminoácidos es eliminada del centro mediante hidrólisis, dejando dos cadenas de péptidos unidas por los puentes de disulfuro. El glucagón, secretado por las células ð de los islotes del páncreas, estimula la conversión de glucógeno hepático en glucosa de la sangre. Es un péptido de cadena larga que contiene 29 aminoácidos. La secretina, producida por la mucosa del duodeno, estimula la secreción de jugo pancreático. Es un péptido que contiene 27 aminoácidos, 16 de los cuales son identificados en sucesión con los del glucagón. La hormona adrenocorticotrópica (ACTH), secretada por el óvulo anterior de la hipótesis, estimula el crecimiento de la corteza adrenal y la producción por esta de esteroides corticoadrenales (es decir, de hormonas). Tiene 39 aminoácidos en una sola cadena de péptidos, 10 de los cuales tienen una posición similar a una parte de la cadena de péptidos en la MSH. La ACTH es sintetizada rápidamente, se almacena poco en la hipófisis y es eliminada pronto en el plasma. Su periodo de semidesintegración biológica, tiempo requerido para que la mitad de una cantidad dada producida sea eliminada del plasma, es de 20 minutos o menos. Un péptido aun más largo, la parathormona, es secretado por las glándulas paratiroides e interviene en la regulación del metabolismo del calcio y el fósforo. La parathormona es una cadena de péptidos que contiene 84 aminoácidos. La calcitonina, secretada por los cuerpos ultimobranquiales situados en la tiroides, tiene una acción antagonista a la de la parathormona. La corteza adrenal. los testículos y el ovario secretan esteroides, sintetizados del colesterol y compuestos de 21, 19 ó 18 carbonos dispuestos en cuatro anillos unidos. ES un hecho curioso que la hormona sexual femenina primaria, el estradiol, con 18 carbonos, es sintetizada a partir de la hormona sexual masculina de 19 carbonos testosterona. Esta, a su vez, es sintetizada partiendo del segundo tipo de hormonas sexual femenina, la progesterona, que tiene 21 carbonos. La progesterona es también un precursor en la corteza adrenal de glucocorticoides y mineralocorticoides. Glucocorticoides como el cortisol estimulan la conversión de proteínas en carbohidratos, y mineralocorticoides como la aldosterona regulan el metabolismo del sodio y del potasio. La corteza adrenal de hombres y mujeres produce dehidroepiandrosterona y adrenosterona, esteroides de 19 carbonos con ligera actividad de hormona sexual masculina, y una pequeña cantidad de la potente hormona testosterosna. Giberelinas. El grupo final de compuestos con actividad hormonal comprende substancias con efectos específicos de promoción del crecimiento en las plantas. Investigando una enfermedad de las plantas de arroz que causaba excesivo alargamiento del retoño, Kurosawa halló que la substancia activa era secretada por unos hongos infeccioso llamado Gibberella fujikuroi y llamó a la substancia giberelina. Se han encontrado nueve o más clases de giberelinas como constituyentes de plantas verdes. Como los esteroides, se derivan de las unidades isoprenoides por intermediarios con dieterpeno de 20 carbonos. GLANDULAS ENDOCRINAS DEL HOMBRE TIROIDES Son un par de glándulas situadas en la zona anterior del cuello, unidas por un estrecho itsmo de tejido situado delante de la traquea, inmediatamente debajo de la laringe. Las tiroides constan de grupos de células epiteliales cuboides dispuestas en folículos, esferas huecas de una célula de espesor. La cavidad de la esfera contiene un coloide gelatinoso, secretado por las células epiteliales.(ver abajo) 6 Las tiroides se desarrollan como excrecencias del piso de la faringes, pero cuando el embrión humano es de 6 mm de longitud, se pierde esta conexión. El efecto fundamental de la tiroxina es acelerar los procesos oxidativos liberadores de energía en todos los tejidos corporales. Si se da una cantidad complementaria, el organismo utiliza más oxígeno, produce desechos en más cantidad y desprende más calor que en condiciones normales. Si se añade tiroxina a una suspensión de mitocondrias se altera la permeabilidad de la membrana mitocondrial, se produce tumefacción de las mitocondrias, y quizá en esta forma se desconecte la fosforilación oxidativa del transporte de electrones. No se ha dilucidado si la tiroxina ejerce este efecto sobre las mitocondrias en el organismo indemne. Si la cantidad de tiroxina es insuficiente, el metabolismo basal desciende hasta 600 o 900 calorías diarias, lo que equivale al 50−30% de la cantidad en condiciones de salud. Si se toman porciones de tejidos separados en un animal en estado de insuficiencia tiroidea, al ser cultivados in vitro, dan también lugar a un bajo metabolismo. Por sus efectos metabólicos, la tiroxina influye extraordinariamente en el crecimiento y formación caracterológica. La extirpación del tiroides en un animal joven provoca disminución del crecimiento, retraso del desarrollo mental y demora o disminución del desarrollo de los genitales. PARATIROIDES Son cuatro masas de tejido del tamaño de un guisante pequeño, adheridas a la substancia de la glándula tiroides. Las células de la paratiroides están dispuestas en una masa compacta y no en folículos como las de la tiroides. Al igual que las tiroides, las paratiroides se originan de excrecencias de la faringe siendo restos evolutivos de la tercera y cuarta bolsas branquiales. PANCREAS ENDOCRINO Dispersas entre las células acinares del páncreas que secretan las enzimas digestivas hay un millón o más de islas de tejido endocrino llamadas islas de Langerhan. Estas contienen tres tipos de células que pueden distingirse fácilmente en los cortes histológicos. Las células ð secretan insulina, las células ð secretan glucagón, y las gamma que segregan somatostatina. GLANDULAS ADRENALES Los pares de glándulas adrenales situados en el extremo superior de cada riñón son combinaciones de dos glándulas totalmente independientes: la médula adrenal secreta adrenalina y noradrenalina, y la corteza adrenal secreta los esteroides corticoadrenales. Los dos pares tienen diferente estructura celular y distinto origen embrionario. En algunos vertebrados inferiores las células correspondientes a la corteza y la médula adrenal están claramente diferenciados y distanciados. La médula adrenal rojo−obscuro pardo se deriva embriológicamente del mismo sitio que le sistema nervioso y sus células, por ello, semejan células nerviosas modificadas. La médula secreta grandes cantidades de adrenalina cuando el hombre o el animal está asustado o irritado. La adrenalina promueve varias respuestas, útiles para hacer frente a urgencias −se eleve la presión arterial, la frecuencia cardiaca aumenta, se eleva el contenido de glucosa en sangre, se contrae el brazo y se libera una reserva almacenada de sangre, se reduce el tiempo de coagulación de la sangre, se dilatan las pupilas y se contraen los músculos que ponen los pelos erectos, proporcionando una piel protectora más gruesa a los animales y pone la "piel de gallina" a los humanos. La corteza adrenal exterior amarillo rosado está compuesta por tres capas de células y secreta varias hormonas fisiológicamente importantes. Glucocorticoides como cortisol estimulan la conversión de aminoácidos en glucosa, mineralocorticoides como aldosterona regulan el metabolismo del sodio y del potasio, favorecen la reabsorción de sodio por los túbulos renales y los sexocorticoides. HIPÓFISIS 7 Está situada en una pequeña depresión de la base del craneo, inmediatamente debajo del hipotálamo, es una glándula del doble de tamaño que las glándulas adrenales. Es la glándula endocrina más importante del organismo. Su lóbulo anterior se forma en el embrión como una excrecencia del cielo de la boca. El lóbulo posterior crece hacia abajo, desde el piso del cerebro. Las dos partes se unen y el lóbulo anterior crece parcialmente alrededor del posterior. El lóbulo anterior pierde su conexión con la boca, pero el lóbulo posterior mantiene su conexión con el hipotálamo. Ciertas células del hipotálamo (el hipotálamo no es una glándula endocrina propiamente dicho, mas regula toda la actividad hormonal) secretan factores liberadores que producen secreción de la hipófisis anterior. La primera en ser caracterizada químicamente fue el factor liberador de tirotropina (TRF), un tripéptido, glutámico−histidina−prolina. El lóbulo anterior contiene por lo menos cinco tipos de células que difieren en su forma, tamaño, propiedades tintoriales y clase de gránulos presentes en el citoplasma. Parece probable que cada tipo produzca y secrete una clase diferente de hormona. La célula que secreta la hormona del crecimiento fue identificada como una célula redondeada cuyo citoplasma contenía numerosos gránulos acidofílicos densos y redondos. Una vez aislados, se comprobó su elevada concentración de hormona del crecimiento. Las células que secretan prolactina tiñen fuertemente con carmín y contienen gránulos mayores y más ovoides que los gránulos de las células que secretan hormona del crecimiento. La acción hipofisiaria se regula a través de la epífisis, que regula a su vez los centros liberadores del hipotálamo . En general, podemos decir que la hipófisis tiene dominio y control de la mayoría del resto de glándulas endocrinas, puesto que secreta hormonas destinadas a la influencia de los islotes de Langerhans, la corteza adrenal y las gónadas. además de segregar, como ya hemos visto, hormona de crecimiento, y regular el metabolismo del agua y las sales. GONADAS Las gónadas estimulan el desarrollo y mantenimiento de las características sexuales primarias y secundarias, la conducta sexual y el embarazo. Todo ello, con la ayuda de la hipófisis y otros centros reguladores (epífisis e hipotálamo). Son los ovarios, en la mujer, y los testículos, en el hombre, los principales centros de regulación sexual o gónadas, aunque se entiende por gónada la macroestructura de desarrollo sexual en el ser humano. Entre los túbulos seminíferos que producen los espermatozoides se encuentran las células intersticiales que producen y secretan las hormonas sexuales masculinas (andrógenos) como testosterona. Si los testículos permanecen en la cavidad abdominal, en lugar de descender al saco escrotal, los túbulos seminíferos degeneran y el hombre es estéril, pero sus células intersticiales son normales y secretan una cantidad normal de testosterona. Las células formadoras de espermatozoides son especialmente susceptibles al calor, y el saco escrotal, que tiene una temperatura inferior a la cavidad abdominal, proporciona un medio ambiente en que las células pueden desarrollarse. Los andrógenos estimulan el desarrollo de los caracteres sexuales secundarios masculinos −la barba, el crecimiento y distribución del vello corporal, la voz grave, el aumento de fuerza y tamaño en los músculos esqueléticos, y el desarrollo de las glándulas sexuales accesorias, la próstata y las vesículas seminales. La testosterona desempeña un papel en la determinación de la conducta sexual masculina y el impulso sexual. Los testículos y los ovarios se desarrollan a partir del mesodermo, del puente genital situado en el lado ventral de los riñones mesofrénicos. Las células de los ovarios que producen y secretan las hormonas sexuales esteroides son las células que revisten la cavidad del folículo y las células del cuerpo amarillo, formadas de estas células después de la 8 ovulación. Los ovarios secretan dos tipos de hormonas sexuales femeninas, estrógenos como el estradiol y progesterona. El estradiol regula los cambios corporales que se producen en la mujer en la pubertad, como es el ensanchamiento de la pelvis, desarrollo de los senos, crecimiento del tuero y la vagina, crecimiento del vello púbico y los genitales externos, cambio en la calidad de la voz y comienzo de ciclo menstrual. La progesterona es necesaria para la terminación de cada ciclo menstrual, para la implantación del óvulo fecundado en útero y para el desarrollo de los senos durante el embarazo. GLANDULA PINEAL También llamada epífisis. Es una pequeña estructura redonda situada en la superficie superior del tálamo tal vez pueda no considerarse como una glándula endocrina propiamente dicha y, por supuesto, no tiene la importancia de las otras mencionadas hasta aquí. Se sitúa entre los hemisferios cerebrales y deriva embriológicamente del cerebro. Secreta melatonina, y en el proceso de secreción intervienen la transferasa hidroxi indol O−metilo y la serotonina. La luz que incide en la retina incrementa la síntesis de melatonina por la glándula pineal. Un pequeño nervio, el conducto óptico accesorio inferior, pasa desde el nervio óptico a través del procencéfalo medio, y se une al sistema nervioso simpático. La secreción pineal inhibe las funciones ováricas directamente o por mediación de un efecto en la hipófisis. Las niñas ciegas de nacimiento llegan a la pubertad antes que las normales por la ausencia de ese efecto inhibidor sobre los ovarios. OTROS ORGANOS ENDOCRINOS menos importantes o considerados ya en este trabajo, son el Timo, estimulante de la actividad inmunológica en los tejidos linfáticos o la placenta. La placenta, además de ser un órgano de sostén y nutrición del feto en desarrollo, también es un órgano endocrino, que secreta esteroides estradiol y progesterona, y dos hormonas proteínicas. La gonadotropina coriónica (HCG) es similar a la hormona luteizante en muchos aspectos, secretada por la hipófisis, y el lactógeno placentario posee ciertas propiedades similares a la hormona del crecimiento y otras similares a la prolactina. Durante la gestación se producen grandes cantidades de gonadotropina coriónica y pasan a la orina de la madre (lo que da lugar a la clásica prueba del embarazo mediante la orina). La placenta produce gran cantidad de esteroides, especialmente en el último o últimos dos meses de la gestación. La placenta de otros mamíferos, como la coneja, producen otro tipo de esteroides, como la relaxina, que produce también el ovario, y que es capaz de relajar los ligamentos de la pelvis inmediatamente antes del parto, para facilitar el paso de la cría a través del canal. Por último, mencionar que el revestimiento del conducto digestivo produce hormonas que estimulan o inhiben la secrección de jugos gástricos (gastrina, secretina, colecistocinina y enterogastrona) ORGANOS BLANCO Todas las hormonas secretadas por las glándulas endocrinas en el hombre y otros vertebrados (con posible excepción de las prostaglandinas) son secretadas en el torrente sanguíneo y transportadas por este a todas partes del cuerpo. Algunas hormonas, como la tiroxina o la hormona del crecimiento afectan a las condiciones metabólicas de todas las células del cuerpo; cada célula responde a la presencia de la hormona y presenta un estado metabólico alterado cuando se la priva de ella. Pero la mayor parte de las hormonas solo afectan a ciertas células del cuerpo, a pesar de que el torrente sanguíneo las lleva por todo el organismo. Las células que responden a una hormona dada se llaman "órganos blanco" de dicha hormona. Solo el páncreas responde a la secretina que circula por la sangre; la glándula tiroides es el órgano blanco de la tirotropina segregada por la hipófisis; el ovario o el testículo es el órgano blanco de las gonadotropinas FSH y LH de la hipófisis. Algunas hormonas, como el estradiol, producen notables efectos en sus órganos blanco primarios, el útero, la vagina y las glándulas mamarias, menores efectos en otros caracteres, la voz, la distribución del vello por el cuerpo, el crecimiento de los huesos, etc... y efectos menores sobre otros tejidos. 9 Está claro ahora que la capacidad de un tejido para responder al estradiol se relaciona con la presencia en ese tejido de una proteína que específicamente toma estradiol y se asocia con él. El útero, la vagina, la hipófisis y el hipotálamo contienen esta proteína y pueden acumular estradiol procedente de la sangre. Se supone que esta proteína desempeña un papel en el transporte de estradiol desde el exterior de la célula hasta el núcleo, donde produce su efecto. Hay pruebas de receptores proteínicos comparables en el oviducto, que son específicos para la progesterona, y receptores en la próstata específicos para testosterona. La especifidad de los efectos de las hormonas peptídicas y proteínicas es también una propiedad de las células que responden a ellas, aunque el mecanismo que interviene puede ser diferente del de una molécula receptora. Se sabe que todas las células tienen exactamente toda la información necesaria para desarrollar cualquier función, así, por ejemplo, las células que sintetizan el alimento en el estómago gracias a ciertas enzimas, tienen gracias al ADN la información necesaria que las capacitaría para realizar la respiración celular que se lleva a cabo en los pulmones o el proceso diurético de los riñones. Simplemente, estas células tienen activadas funciones para las que han sido programadas, sin embargo, con otro tipo de información genética, se podría cambiar la información de las células para que activasen otra o varias funciones diferentes de para las que fueron diseñadas (lo que causaría, lógicamente, un cáncer). Esto explica el porqué unas células recogen ciertas hormonas, y otras no, o por que unas células secretan ciertas hormonas y otras no: todo está en el código genético, en el ADN. El caso de la feminización testicular A propósito del tema, un fenómeno curioso que se procede por irregularidad en el ADN, es el de la feminización testicular, una afección humana hereditaria. Los individuos con esta afección son varones genéticos XY, y secretan niveles masculinos normales de testosterona, aunque fenotípicamente mujeres, con desarrollo típico de pecho femenino y genitales externos femeninos. La vagina es una bolsa ciega corta y el resto de los órganos genitales internos son rudimentarios o están completamente ausentes. Las gónadas son testículos no descendidos típicos. Después de la pubertad, estos individuos adquieren proporciones, hábitos y voz femeninos, y son generalmente muy atractivos. Las células de estos individuos son incapaces de responder a andrógenos (o eso era hace algunos años, cuando esta información se publicó. N. del A.) dado que la información genética de esas células va modificada desde el origen, y probablemente si se lograra una respuesta, la célula mutaría produciendo cáncer. Así, el desarrollo de estos individuos es en parte bajo el estímulo de los estrógenos secretados por los testículos y la glándula adrenal. ANORMALIDADES ENDOCRINAS Las funciones de las glándulas endocrinas son de un interés mayor que el teórico para el hombre, pues la falta o el exceso de actividad de esas glándulas puede producir marcados efectos sobre el cuerpo y, por ende, sobre la conducta, que es lo que en última instancia interesa a la psicobiología. Curiosamente, el hombre se dio cuenta de la existencia de las glándulas endocrinas cuando estudiaba las enfermedades funcionales −para diferenciarlas de las infecciosas o por deficiencia− que producían. MIXEDEMA Una deficiencia en la cantidad de tiroxina secretada por las tiroides en un adulto produce mixedema, caracterizada por un bajo índice metabólico y reducida producción de calor. La temperatura corporal puede descender varios grados por debajo de la normalidad, por lo que el enfermo sentirá un frío constante. Su pulso es lento y se muestra física y mentalmente letárgico. Sin embargo, su apetito suele permanecer normal, y como el alimento consumido no es usado al ritmo normal, hay tendencia a la obesidad. La piel se vuelve cérea y entumecida, debido a la deposición de líquido mucoso en los tejidos subcutáneos, y generalmente se cae el pelo. El mixedema responde bien a la administración de tiroxina o glándula tiroides desecada. Como la tiroxina no es digerida apreciablemente por los jugos digestivos, puede darse por la boca. 10 El mixedema es causado por un exceso de actividad o degeneración de la glándula tiroides. Otro tipo de hipotiroidismo resulta cuando la dieta no contiene suficiente yodo para la síntesis de la tiroxina. La glándula tiende a compensar la insuficiencia aumentando de tamaño. El agrandamiento correspondiente, conocido como bocio simple, puede ser una pequeña hinchazón o una masa grande y desfigurante, de varios kilos de peso. Los síntomas que acompañan al bocio se parecen a los del mixadema, pero son más benignos. Este tipo de bocio aparece en lugares donde el suelo carece de yodo, o regiones apartadas del mar donde no pueden obtenerse pescados y marisco. Sin embargo, en la actualidad, esta enfermedad queda relegada a países tercermundistas o regiones de interior pobres en yodo, si, pero también económicamente pobres. El hipotiroidismo presente desde el nacimiento se conoce como cretinismo. Los niños que padecen esta enfermedad son enanos de poca inteligencia que nunca maduran sexualmente. Si el tratamiento con tiroxina es temprano, puede lograrse la normalización sin ningún problema. BOCIO EXOFTÁLMICO El hipertiroidismo es resultado de la hiperactividad de una glándula de tamaño normal o de un aumento de la glándula misma. En ambos casos, el índice metabólico basal aumenta hasta casi el doble de lo normal. La producción de calor excesivamente rápida hace que el hipertiroideo se sienta incómodamente caliente y transpire profusamente. Como el alimento que ingiere es usado rápidamente, tiende a perder peso aun con una dieta de alto contenido calórico. Alta presión arterial, tensión nerviosa e irritabilidad, debilidad muscular y temblores son sintomáticos de la afección. Pero probablemente el síntoma más característico es la profusión del globo del ojo, llamado exoftalmos, que da al paciente una expresión de mirada fija. El aumento de la glándula como consecuencia de la enfermedades se conoce con el nombre de bocio exoftálmico, para distingirlo del bocio simple. Síntomas idénticos pueden darse por la ingesta de tiroxina en las personas normales. El hipertiroidismo puede ser tratado por extirpación quirúrjica de parte de la glándula tiroides o destruyendo las células con radio o laser. El hipertiroidismo puede ser tratado también con éxito administrando tiouracilo, que inhibe la síntesis de la tiroxina, o inyectando yodo radiactivo. TETANIA La secrección de las glándulas paratiroides, parathormona, es esencial para la vida porque regula la concentración de calcio y fosfato en la sangre y en los tejidos. Cuando se extirpan las glándulas paratiroides, el animal sufre temblores musculares, calambres y convulsiones como respuesta a los estímulos, que en un animal con paratiroides solo produciría leves respuestas. Esta afección, llamada tetania, se debe a un aumento de la irritabilidad de los nervios y músculos causados por la reducción del contenido de calcio en sangre y líquidos tisulares. Si se inyecta una solución de sal de calcio a una vena de un animal que sufre convulsiones tetánicas, estas cesan al cabo de unos minutos, y pueden evitarse más contracciones con la inyección repetida de calcio o la ingesta vía bucal. La cantidad de fósforo en la sangre aumenta cuando disminuye el calcio y se reduce por la inyección de parathormona. La principal función, pues, de la hormona paratiroides, es regular el contenido de calcio, magnesio y fósforo de la sangre y los líquidos tisulares. La parathormona favorece la absorción de calcio de la luz del intestino, la liberación de calcio de los huesos y la reabsorción de calcio del filtrado glomerular de los túbulos renales. Inhibe también la resorción de fosfato en los túbulos renales y favorece así la excreción de fosfato en la orina. Se ve hiperfunción de las paratiroides en tumores o aumento de tamaño de las glándulas, y se caracteriza por un alto nivel de calcio en sangre. Puesto que el calcio procede, al menos en parte, de los huesos, el hiperparatiroidismo se caracteriza por huesos blandos que se doblan y fracturan con facilidad. Los músculos son menos irritables que normalmente, y pueden atrofiarse y ser dolorosos. Al elevarse el nivel de calcio en sangre, el mineral es depositado en lugares anormales como el riñón. La enfermedad puede tratarse quirúrjicamente o por rayos, para eliminar el tejido paratoideo sobrante. 11 En cuanto a la conducta, dado que el fósforo y el calcio juegan un papel vital en la neurotransmisión (sinapsis) es lógico pensar que estas se van a ver afectadas de algún modo por una cantidad anómala de estas sustancias en la sangre. DIABETES La insulina aumenta la velocidad con que toman la glucosa de la sangre ciertas células, especialmente células de músculos esqueléticos, y se convierte en glucosa−6−fosfato. Esto reduce la concentración de glucosa en sangre, aumenta el almacenamiento de glucógeno en los músculos y el metabolismo de glucosa a dióxido de carbono y agua. Una deficiencia de insulina reduce la utilización de glucosa, y las resultantes alteraciones en el metabolismo de los carbohidratos producen cambios en el metabolismo de las proteínas, las grasas y otras substancias. La hipofunción del páncreas en la diabetes causa dificultad en la utilización de glucosa, elevada concentración de glucosa en sangre, y excreción de grandes cantidades de glucosa en la orina, porque la concentración en la sangre supera el umbral renal. Se requiere más agua para excretar ese azúcar, aumenta el volumen de orina y el paciente se deshidrata y siente sed. Los tejidos, incapaces de obtener bastante glucosa de la sangre, convierten la proteína en carbohidrato. Gran parte de este también es excretado y hay una progresiva pérdida de peso, los depósitos de grasa son movilizados y se metabolizan los lípidos. El incremento de la oxidación de las grasas provoca una acumulación de ácidos grasos incompletamente oxidados. Estos cuerpos cetónicos son volátiles y tienen un sabor dulce, que da al aliento de los diabéticos su olor característico. Los cuerpos cetónicos son ácidos y deben ser excretados con la orina, causando acidosis (reducción de la reserva alcalina de los líquidos corporales). La diabetes no tratada es finalmente mortal por la acidosis, la toxicidad de los cuerpos cetónicos acumulados y la continua pérdida de peso. La inyección de insulina alivia todos los síntomas diabéticos, pero se debe hacer repetidamente para obtener resultados continuos. La insulina no cura la diabetes, porque el páncreas no empieza a secretar su hormona de nuevo, pero las inyecciones continuas impedirán la aparición de síntomas y capacitan al diabético para una vida normal. Hay una base hereditaria en la diabetes. Si en el tratamiento de diabetes se inyecta demasiada insulina, el nivel de azúcar en sangre desciende drásticamente y el paciente sufre un shock. El último tratamiento para la diabetes que está en investigación consiste en la administración al paciente de islotes de Langerhans los cuales, vía sanguinea, quedarán estancados en el hígado donde tendrán una funcionalidad perfecta. ENFERMEDAD DE ADDISSON La enfermedad de Addisson es resultante de la insuficiente secreción de la corteza adrenal. Es causada normalmente por una infección tubercular o sifilítica de la corteza que destruye sus células. Se caracteriza por una baja presión arterial, debilidad muscular, trastornos digestivos y aumento de excreción de sodio y cloruro en la orina, aumento de la concentración de potasio en los líquidos y células corpolas y un bronceado peculiar de la piel causado por la deposición inhabitual de melanina. La enfermedad de Addisson es tratada por la administración bucal o intravenosa regular de un esteroide adrenal natural, como cortisol, o de un esteroide sintético, como dexametasona o acetato 2a−metil9 ð−fluorocortisol.Este último es extraordinariamente potente en actividades mineralocorticales y glucorticoides. La corteza adrenal de hombres y mujeres produce dehidroepiandrosterona y adrenosterona, esteroides con ligera actividad hormonal sexual masculina. Además, la corteza produce pequeñas cantidades de testosterona. 12 DIABETES INSIPIDA La lesión del lóbulo posterior de la hipófisis o de sus conductos nerviosos puede producir una deficiencia hormonal que causa diabetes insípida. Esta enfermedad, caracterizada por imposibilidad de concentrar orina por el riñón, hace que el paciente excrete hasta treinta o cuarenta litros de orina diariamente, y por tanto padezca sed excesiva. La inyección de vasopresina no cura la enfermedad, de igual modo que la insulina no cura la diabetes sacarina, pero proporciona alivio de todos los síntomas con inyecciones repetidas. HIPO e HIPERPITUITARISMO La importancia de la hipófisis en la economía corporal se demuestra al extirpar experimentalmente la glándula. Los animales en periodo de crecimiento, dejan inmediatamente de desarrollarse, y los adultos, muestran regresión en los caracteres sexuales, y una correspondiente atrofia de la glándula tiroides y la corteza adrenal. Cuando se inyectan extractos hipofisiarios en animales jóvenes, se estimula el crecimiento y se alcanza el pleno desarrollo sexual en una edad temprana. La corteza adrenal, la tiroides y los órganos sexuales, responden creciendo mucho anormalmente y secretando en exceso. El gigantismo es un síndrome directamente relacionado con el desarrollo hipofisiario. La primera hormona de esta glándula que se descubrió fue la hormona estimulante del crecimiento, aislada finalmente como proteína pura a partir de extractos de glándula hipofisiaria de buey. Esta hormona controla el crecimiento general del cuerpo, y especialmente el crecimiento de los huesos largos. En consecuencia, cuando la hipófisis está hiperactiva durante el periodo de crecimiento, hay una aceleración del proceso que conlleva una persona muy alta, aunque bastante bien proporcionada. Si la hipófisis está hipoactiva durante el periodo de crecimiento, el resultado es una persona pequeña, o enano. La hipersecreción de hormona del crecimiento, después de la época de crecimiento, produce un estado conocido como acromegalia. Como en esta época la mayoría de las partes del cuerpo han perdido capacidad para crecer, solo se desarrollan las manos, los pies, y los huesos de la cara. Las manos y los pies aumentan de tamaño toscamente, las quijadas crecen anormalmente en longitud y anchura y los bordes óseos de los ojos y las mejillas aumentan de tamaño. La hormona del crecimiento, además de su efecto general en el crecimiento, produce algunos efectos específicos en el metabolismo intermediario. Afecta: 1) al metabolismo de las proteínas, provocando retención de proteína en los tejidos y reduciendo la velocidad con que los aminoácido se convierten en urea. 2) al metabolismo de las grasas, conduciendo a una movilización de los lípidos en el tejido adiposo y aumentando la concentración de lípidos en la sangre. 3) al metabolismo de los carbohidratos, aumentando los depósitos de glucógeno en el hígado y el músculo, pero oponiéndose en general a los efectos de la insulina. La hormona del crecimiento produce hiperglucemia, y si se administra largo tiempo a ciertos animales, puede producir un estado diabético permanente. DIFERENTES FUNCIONES DE LAS SECRECIONES INTERNAS TRANSMISORAS DE INFORMACIÓN Quizá el modelo más sencillo de acción hormonal es la clase de transmisor de información ejemplificada por la secretina. La presencia de alimento ácido en el duodeno estimula la liberación de secretina por las células de la mucosa duodenal. La secretina pasa, por la sangre, a todas las células del cuerpo. La disposición 13 específica de aminoácidos en el péptido 27−aminoácido significa poco o nada para cualquier célula, excepto para las células acinares del páncreas, y para ellas significa secrete sus enzimas. Las células de la mucosa duodenal continúan secretando secretina mientras exista alimento ácido. También secretan colecistocinina, otro transmisor de información. Esta hormona estimula la vesícula biliar para contraerse y liberar bilis, que pasa por el conducto biliar hasta el duodeno y desempe_a un papel en la neutralización del contenido duodenal. Así, cesa de producir el estímulo para la secreción de ambas hormonas. CONTROLES DE LÍMITES Un segundo modelo de función endocrina, algo más complejo, es aquel en el que dos hormonas actúan juntas para establecer límites superiores e inferiores para cierta función fisiológica. Como ejemplo, consideremos la regulación de la cantidad de glucosa en sangre por las hormonas insulina y glucagón. Tras una comida rica en carbohidratos o una inyección de glucosa, la concentración de glucosa en sangre se eleva. Parte de la glucosa es almacenada por el hígado como glucógeno, pero la concentración elevada de glucosa es la se_al para la secreción de insulina. Una elevación de la concentración de insulina en la sangre puede ser descubierta dos minutos después de una elevación inducida experimentalmente de la glucosa en sangre. Un efecto importante de la insulina es un espectacular aumento de la velocidad de transporte de la glucosa hacia el músculo esquelético y el tejido adiposo. Esto provoca una reducción de la concentración de glucosa en sangre. La secreción de glucagón por las células alfa del páncreas es controlada también por la concentración de glucosa en la sangre, pero es inhibida por altas concentraciones de glucosa y estimulada por bajas concentraciones de glucosa. Así, cuando las concentraciones de glucosa disminuyen por debajo de al variación óptima, se estimula la liberación de glucagón. Este, activando el sistema glucógeno fosforilasa en el hígado, estimula la conversión de glucógeno en glucosa 1−fosfato el cual es convertido, a su vez, en glucosa libre por medio de glucosa 6−fosfato y es secretada. La glucosa secretada por el hígado vuelve normal la concentración de glucosa en sangre y sirve para definir el limite inferior. El páncreas vigila constantemente la concentración de glucosa en la sangre que pasa por él y secreta insulina o glucagón, según se requiera. Un aumento de la concentración de glucosa en la sangre sirve de estímulo a un sistema −la secreción de insulina por las células beta de las islas del páncreas− que la volverán a la normal. Una disminución de la concentración de glucosa en la sangre sirve de estímulo al sistema opuesto −la secreción de glucagón por las células alfa de las islas pancreáticas−, que la volverán a la normal. Otro ejemplo de un sistema de controles limitadores es la regulación de la concentración de calcio en la sangre por las hormonas parathormona y calcitonina, que tienden a aumentar y disminuir, respectivamente, la concentración de calcio en la sangre. La parathormona secretada por las glándulas paratiroides, provoca la pérdida de calcio por huesos y dientes, y una mayor concentración de calcio en la sangre. La parathormona es secretada como respuesta a una reducción de la concentración de calcio en la sangre, y provocando la disolución de minerales del hueso, libera calcio (y fosfato) y aumenta la concentración de calcio en la sangre. La calcitonina, secretada por los cuerpos ultimobranquiales dispersos en la tiroides como respuesta a un aumento de la concentración de calcio en la sangre, estimula la deposición de fosfato cálcico en los huesos. Así, la calcitonina regula el límite superior de concentración de calcio y la parathormona regula su límite inferior. CONTROLES DE RETROALIMENTACIÓN RECÍPROCOS O NEGATIVOS El principal glucocorticoide producido por la corteza adrenal humana es el cortisol. Esta hormona favorece la movilización de aminoácidos desde el músculo esquelético y otros tejidos periféricos y aumenta la conversión, en el hígado, de las cadenas de carbonos de estos aminoácidos en glucosa y glucógeno, proceso denominado gluconeogénesis (nueva formación de glucosa). Reduce la utilización de glucosa en tejidos periféricos y favorece la movilización de lípidos y la formación de cuerpos cetónicos. Sus efectos en general son opuestos a los de la insulina. 14 La síntesis de cortisol en la corteza adrenal es esimulada por ACTH. Esta hormona hipofisiaria aumenta el tama_o de la corteza adrenal estimulando la síntesis de ARN y prolactina. Además, estimula específicamente la producción de cortisol a partir de colesterol, incrementando la actividad de una o más enzimas que intervienen en esta conversión. Así, la estimulación de la corteza adrenal por ACTH aumenta la producción de cortisol y la concentración de cortisol en la sangre. Esto, a su vez, controla, por una relación de retroalimentación negativa, la secreción de ACTH por la hipófisis. El cortisol puede hacer esto inhibiendo directamente la síntesis de ACTH en la hipófisis o indirectamente reduciendo la producción de factor liberador de corticotropina, CRF, por el hipotálamo. El estímulo primario que provoca la secreción de cortisol es cierta clase de tensión física −una lesión, una quemadura, una enfermedad dolorosa, exposición al calor o al frío− que envía impulsos al cerebro, retransmitidos al hipotálamo. Este secreta el factor liberador de corticotropina, que pasa por un sistema portal especial de vasos sanguíneos directamente al lóbulo anterior de la hipófisis. La CRF estimula las apropiadas células de la hipófisis para que secreten ACTH, y esta es transportada por la sangre a la corteza adrenal, donde estimula la producción y liberación de cortisol. La movilización de aminoácidos y lípidos de los tejidos periféricos y la gluconeogénesis en le hígado proporcionan sustratos para la reparación del da_o y reducen el estímulo que provoca la producción del factor liberador. La mayor concentración de cortisol en la sangre actúa en el hipotálamo o la hipófisis o reduce la producción y liberación de ACTH. Un defecto heredado de cualquiera de varias enzimas que participan en la síntesis de cortisol a partir de pregnenolona y colesterol p_ede producir un aumento del tama_o de la corteza adrenal. El defecto más común es una deficiencia de la enzima que a_ade un grupo hidróxilo en el carbono 21 del esteroide. Tal defecto provoca una acumulación de intermediarios en la vía biosintética. Algunos de estos intermediarios no pueden ser convertidos en cortisol, pero sí en andrógenos como androstenediona. Esta puede ser convertida, en la glándula adrenal o en otra parte del cuerpo, en testosterona, el andrógeno más potente. La imposibilidad por la corteza adrenal de secretar cortisol produce hipersecreción de ACTH, porque no hay nada para cortarla. La glándula adrenal crece más y secreta aún más andrógenos, y el individuo resulta virilizado. Un feto femenino que carece de la enzima tiene órganos genitales externos masculinizados en grado variable, pero un feto masculino con el defecto enzimático puede no mostrar anormalidad al nacer. Después del nacimiento la virilización progresa en hombres y mujeres con aumento del tama_o del falo, desarrollo temprano del vello púbico y axilar, reducción del tono de la voz y otros efectos de los andrógenos. Los pacientes con esta afección, hiperplasia corticoadrenal, pueden ser tratados inyectándoles cortisol para que cese la producción de ACTH por la hipófisis. Un sistema de retroalimentación similar regula la síntesis de TSH por la hipófisis y de tiroxina por la glándula tiroides. La hipófisis anterior secreta TSH que estimula varios procesos de la glándula tiroides que participan en la producción y secreción de tiroxina. Un aumento de la concentración de tiroxina en la sangre provoca una reducción de producción de TSH, mientras que una disminución de la concentración de tiroxina en la sangre provoca un aumento en la producción de TSH. Esta relación recíproca de retroalimentación mantiene una concentración bastante constante de ambas en la sangre. La liberación de TSH por la hipófisis puede elevarse por un factor liberador de TSH secretado por el hipotálamo en respuesta a estimulación neural desencadenada por bajar la temperatura del cuerpo. El factor liberador aumentado eleva la producción de TSH: esta, a su vez, aumenta la secreción de tiroxina, y la tiroxina aumenta el índice metabólico de todas las células corporales y eleva la temperatura de todo el cuerpo. La existencia de estos controles de retroalimentación plantea ciertos problemas en el tratamiento de un paciente con una hormona. Dando tiroxina se deprime la producción de TSH, de igual modo que con la tiroxina endógena, y esto reducirá la producción de tiroxina por la glándula tiroides del paciente. Análogamente, dando cortisol a un paciente se reduce la producción de ACTH por su hipófisis, lo cual reduce su tama_o de corteza adrenal y su secreción de cortisol. Cuando un paciente ha sido tratado largo tiempo con tiroxina y se suspende repentinamente el tratamiento, la súbita reducción de concentración de tiroxina en la 15 sangre puede causar una secreción mayor que la normal de TSH por la hipófisis y producir una estimulación indebida de la tiroides. COMPLEJOS SISTEMAS DE CONTROL: REGULACIÓN DE LOS CICLOS ESTRUAL Y MENSTRUAL Para asegurar la fecundación y la concepción, un ovocito 2º debe ser liberado por el ovario en el momento en que el espermatozoide es probable que está presente en el oviducto para fecundarlo y cuando el revestimento del útero, el endometrio, se encuentre en el estado apropiado para permitir la implantación y desarrollo del óvulo fecundado. La sincronización y coordinación de estos hechos es controlada por un conjunto de ocho hormonas y una serie de hechos ocurridos en el sistema nervioso central. Tres de las hormonas son factores liberadores específicos secretados por el hipotálamo, que pasan por el sistema portal hipofisiario de vasos sanguíneos hasta la hipófisis anterior (fig. 25−13). El sistema portal comienza con un plexo de capilares en la eminencia mediana situada en la base del hipotálamo y forma vasos que se extienden por el tallo hipofisiario hasta el lóbulo anterior de la hipófisis, donde se unen con una segunda red de sinusoides y capilares. El factor liberador de hormona estimulante de los folículos, FRH, el factor liberador de hormona luteinizante, LRH, y el factor liberador de hormona luteotrópica, LTRF, son peque_os péptidos que específicamente provocan la liberación de la correspondiente hormona proteínica por la hipófisis anterior. La hormona estimulante de los folículos actúa sobre el ovario y hace que algunos de sus folículos primarios comiencen a desarrollarse. Las células que rodean al huevo proliferan rápidamente y comienzan a secretar estradiol. Una segunda hormona hipofisiaria, la hormona luteinizante, provoca mayor crecimiento del folículo, y una liberación de LH, unida a una liberación de FSH, hace que el folículo se rompa, desprendiendo el óvulo, proceso llamado ovulación. Las restantes células foliculares bajo la estimulación de LH y hormona luteotrópica LTH aumentan de tama_o, adquieren un aspecto grasoso amarillo y forman un cuerpo luteo en el espacio anteriormente ocupado por el folículo. Las células del cuerpo amarillo secretan progesterona y estrógenos. El cuerpo amarillo humano persiste dos semanas si no se produce fecundación, y luego degenera. La hormona estrógena primaria secretada por el ovario, estradiol, estimula la aparición de los caracteres sexuales femeninos y desempe_a un papel en la regulación de los cambios cíclicos del ciclo estrual o menstrual. El estradiol estimula el crecimiento del útero en la pubertad, haciendo que las células musculares aumenten en número y tama_o. El crecimiento de la vagina, el desarrollo de los labios, el clítoris y otros órganos genitales externos, el crecimiento del vello púbico, el ensanchamiento de las caderas y cambios en la estructura de los huesos pélvicos hasta adquirir el tipo femenino, el crecimiento de los senos, la proliferación de las células glandulares en ellos y la deposición de grasa en caderas y muslos característicos de las mujeres adultas son provocados por estradiol. El crecimiento del revestimiento endometrial del útero durante la primera parte de la fase proliferativa de cada ciclo menstrual es controlada por estradiol (fig. 25−14) La progesterona, segundo tipo de hormona sexual femenina, produce poco efecto en el desarrollo de los caracteres sexuales femeninos, pero uno importante en la estimulación del desarrollo del endometrio durante la fase secretoria del ciclo menstrual hasta un estado apropiado para la implantación de un óvulo fecundado. Las células del endometrio acumulan glucógeno y grasas, y sus glándulas se desarrollan y secretan un líquido nutritivo. Los vasos sanguíneos del endometrio crecen, se alargan y se enrollan, La progesterona también prepara los pechos para la secreción de leche aumentando la actividad secretora de las células de las glándulas. Las hembras de muchas especies de mamíferos muestran periodos cíclicos de impulso sexual y permiten la copulación sólo en ciertas épocas, conocidas como periodos de estro o calor, cuando las condiciones son óptimas para la fecundación del óvulo. Muchos animales silvestres sólo tienen un periodo de estro por a_o, la perra y la gata tiene dos o tres, y las ratas y ratonas tienen periodos de estro cada cinco días. El estro se caracteriza por aumento del impulso sexual, ovulación y cambios en el revestimiento del útero y de la vagina. Después del estro el endometrio engruesa y se desarrollan sus glándulas y vasos sanguíneos para proporcionar 16 un medio ambiente óptimo para la implantación del embrión. El ciclo menstrual de los primates se caracteriza por periodos de sangrado vaginal llamados menstruación (del latín menstrualis, mensual), que son resultado de la degeneración y esfacelación del revestimiento endometrial del útero. La ovulación se produce en el punto medio entre dos menstruaciones sucesivas. Los primates, a diferencia de otros animales mamíferos muestran poco o ningún cambio cíclico en el impulso sexual y permiten la copulación en cualquier tiempo del ciclo menstrual. Un punto clave en los ciclos estrual y menstrual es la ovulación, la liberación de un óvulo por el ovario. Un grupo de mamíferos −conejo, gato, hurón, visón− son ovuladores reflejos y la estimulación nerviosa del apareamiento actúa reflejamente para provocar la ovulación. Este acoplamiento directo de copulación y concepción puede explicar la conocida rápida procreación de los conejos. La estimulación eléctrica directa de la región apropiada del hipotálamo, el tuber cinerum y las regiones preópticas, puede causar ovulación en la coneja, gata o la mona. En la coneja, la estimulación neural del coito provoca la liberación de LRF, factor liberador de hormona luteinizante, por el hipotálamo. El LRF pasa por el sistema portal y llega a la hipófisis anterior, provocando una notable elevación, una ola, de liberación de LH por la hipófisis. Esta secreción pasa al ovario y hace que el folículo se rompa y desprenda el óvulo.(fig. 25−15) El segundo y mayor grupo de mamíferos son ovuladores espontáneos en los que la ovulación no es estimulada por el coito, sino que la sincronización y la frecuencia de la ovulación pueden ser influidas por factores ambientales. La rata de laboratorio, mantenida en condiciones normales de iluminación diurna y nocturna, ovula muy temprano por la ma_ana, entre las 1:00 y 2:30 horas. Si se mantienen las ratas dos semanas o más en condiciones artificiales en las que los periodos de luz y oscuridad se invierten, el tiempo de ovulación se modifica 12 horas. Ratas expuestas a luz continua durante las 24 horas del día terminan por cesar de ovular y muestran persistente cornificación vaginal. Los primates con ciclos menstruales, experimentan ovulación espontanea. Existen considerables pruebas de que el ritmo del ciclo menstrual humano, y probablemente también de la ovulación, es influido por factores ambientales. Enfermeras de turno nocturno y azafatas de líneas aéreas que recorren largas distancias hacia el este y el oeste en diferentes zonas horarias informan de frecuentes cambios en sus ciclos menstruales. Los ovuladores espontáneos parece que tienen cierta clase de reloj hipotalámico dependiente de la luz que proporciona el estímulo neural para la liberación de los factores liberadores hipotalámicos. Los factores liberadores son producidos por neuronas terminadas en la eminecia mediana, donde son liberados y pasan a los vasos portales hipofisiarios, por donde llegan a la hipófisis. Esto estimula la liberación de una ola de LH que inicia la ovulación en el ovario (fig. 25−16) La hipófisis de machos y hembras produce y secreta FSH y LH, que controlan le desarrollo y la función de los testículos. La hipófisis de un macho trasplantada a una hembra hiposectomizada soportará un ciclo estrual o menstrual normal. Un ovario transplantado a un macho castrado desarrollará folículos maduros, pero no sufrirá ovulación −no hay liberación cíclica de una ola de LH por la hipófisis del animal macho. La diferencia entre los dos sexos −el desarrollo del reloj hipotalámico− aparece en una etapa temprana crítica del desarrollo del sistema nervioso central. En el macho en desarrollo la testosterona inhibe el desarrollo del centro cíclico. Hembras jóvenes que carecen de testosterona desarrollan un reloj hipotalámico que regula la liberación rítmica de gonadotropinas que regulan los ciclos sexuales. Si se inyecta testosterona en una rata hembra en cualquier momento entre el segundo y el quinto día después de nacida se suprime permanentemente la actividad del centro cíclico del hipotálamo. Nunca ovula, pero permanece en un estado de estro constante, similar a una rata mantenida con iluminación constante. En la regulación de los ciclos estrual y menstrual participan dos tipos de mecanismos de retroalimentación. De igual modo que la tiroxina y el cortisol reducen la secreción de TSH y ACTH respectivamente por la hipófisis, las concentraciones de estradiol y progesterona en la sangre afectan a la producción de gonadotropinas hipofisiarias producen un efecto de retroalimentación negativo en el hipotálamo y reducen la producción de factores liberadores. 17 La creación de métodos sensibles y precisos para medir FSH y LH por radiovaloración inmunológica y para medir el estradiol y la progesterona por métodos específicos de enlace de proteínas ha hecho posible la medición de cada uno de estos en una peque_a muestra de sangre tomada cada día de un ciclo menstrual. Tales valoraciones muestran que las concentraciones de LH y FSH se elevan repentinamente y luego descienden durante un periodo de dos a tres días en la mitad del ciclo menstrual. La concentración de FSH en la sangre se eleva también durante la primera semana de fase proliferativa, empezando durante la menstruación anterior y continuando después de cesar el flujo. La concentración de estradiol en la sangre es baja durante los 10 primeros días de la fase proliferativa pero se eleva bruscamente y alcanza un máximo cuando la concentración de LH comienza a elevarse. La concentración de estradiol baja luego y ha alcanzado casi el nivel basal bajo en el momento en que la concentración de lH llega a su punto máximo. Un segundo pico, más ancho y bajo, de concentración de estradiol se ve típicamente durante la fase secretoria del ciclo. La concentración de progesterona en la sangre comienza a elevarse en el momento del pico de LH y alcanza un pico seis días después. Permanece alta hasta casi el final del ciclo, luego baja y la retirada de la progesterona precipita una reducción del flujo sanguíneo al endometrio, la muerte y esfacelación de las células del revestimiento endometrial y el flujo menstrual. Este, constituido por tejido endometrial necrótico, sangre que rezuma de los extremos rotos de los vasos sanguíneos endometriales y líquido tisular de la superficie uterina, es gradualmente expulsado del útero en un periodo de tres a cinco días. La porción basal del endometrio permanece intacta durante la menstruación y es la fuente del nuevo epitelio y glándulas que se desarrollan bajo la influencia del estradiol después de la menstruación. Análisis de muestras de sangre extraidas cada 25 minutos de mujeres normales mostraron que la LH no es liberada a una velocidad constante y no está presente en la sangre en una concentración constante. Por el contrario, el LH es liberado a borbotones, resultando un breve pico; la concentración disminuye entonces gradualmente en una curva de desintegración exponencial hasta que se libera el segundo borbotón una hora después. Así, la liberación de LH, y probablemente de otras hormonas hipofisiarias, es un proceso rítmico regido probablemente en gran parte por la liberación rítmica de LRF por el hipotálamo. La regulación del ciclo menstrual sigue la siguiente serie de hechos (fig. 25−18). 1. Después de la menstruación anterior, la retirada de progesterona elimina su influencia inhibidora sobre el hipotálamo y se libera FRF, estimulando la liberación de FSH por la hipófisis. Esto explica el elevado nivel de FSH circulante en la sangre al principio del ciclo. 2. La FSH hace que uno o más folículos del ovario aumenten rápidamente de tama_o y comienzen a secretar estradiol. El estradiol secretado por las células foliculares provoca la proliferación de las células endometriales. Cuando el folículo alcanza cierto tama_o, secreta una oleada de estradiol. La oleada de estrógeno desencadena, a su vez, una oleada de FSH y LH que causa ovulación. 3. El pico de LH, junto con la hormona luteotrópica secretada por la hipófisis, hace que las células foliculares experimnten luteinización, formando un cuerpo amarillo y aumentando la secreción de progesterona. 4. La concentración de progesterona en sangre se eleva y p_rmanece alta durante la mayor parte de la fase secretora del ciclo. La progesterona causa el continuo crecimiento del revestimiento endometrial y estimula las glándulas endometriales para que secreten un líquido nutriente. 5. La progesterona tiene también una importante función en inhibir la liberación de FSH y evitar el desarrollo de más folículos y óvulos. Finalmente, el cuerpo amarillo comienza a regresar, la concentración de progesterona en la sangre desciende y el hipotálamo, liberado de sus efectos inhibidores, libera FRF y comienza un nuevo ciclo. 18 Si el óvulo es fecundado y se implanta en el endometrio, las células del trofoblasto de la placenta en desarrollo secretan gonadotropina coriónica. Sus fuertes actividades uteinizantes y luteotrópicas mantienen el cuerpo lúteo y estimulan la continua secreción de progesterona. En la decimosexta semana de gestación la placenta produce suficiente progesterona, por lo que no se necesita ya el cuerpo amarillo y experimenta involución. La progesterona bloquea la ovulación no por un efecto directo en el ovario, sino impidiendo la secreción de los factores liberadores FRF y LRF por el hipotálamo. Los contraceptivos bucales contienen estrógenos sintéticos y progestinas que bloquean la ovulación en forma similar. La s hormonas naturales estradiol y progesterona son rapidamente metabolizadas en el cuerpo, pero las hormonas sintéticas producen ligeros cambios en la estructura molecular que reduce notablemente la velocidad con que son destruidas. Los contraceptivos bucales, como las hormonas naturales, inhiben la liberación de LH por la hipófisis, impidiendo así la ovulación. Una mujer que toma la píldora no tiene oleada de LH y FSH en la mitad del ciclo y no ovula. Algunas progestinas sintética alteran el caracter del moco secretado por el cuello uterino y hacen que sea penetrado menos fácilmente por el espermatozoide, reduciendo así la probabilidad de fecundación. Después de liberado un óvulo por el ovario y de pasar al oviducto, conserva la capacidad para ser fecundado un tiempo relativamente corto, 24 horas. El espermatozoide depositado en el conducto reproductor femenino durante el coito retiene su capacidad para fecundar un óvulo por espacio de 48 horas o menos. El periodo de fecundidad máxima en los seres humanos se reduce así a unos pocos días en la mitad de dos periodos menstruales sucesivos, en el tiempo de la ovulación. Es dificil determinar con precisión cuándo se produce este periodo de fecundidad máxima, a causa de la variabilidad en el tiempo de la ovulación del ciclo menstrual. COMPLEJOS SISTEMAS DE CONTROL: REGULACIÓN DEL DESARROLLO Y FUNCIÓN DE LA GLÁNDULA MAMARIA El crecimiento y desarrollo de las glándulas mamarias después de la pubertad y la producción y secreción de leche tras el parto son controlados por una compleja serie de acontecimientos hormonales. Después de la pubertad se desarrollan los senos y sus elementos glandulares proliferan bajo el control del estradiol. La progesterona produce mayor desarrollo de las glándulas que han sido previamente estimuladas por el estradiol. Durante la gestación los senos experimentan más desarrollo, estimulados por grandes cantidades de estradiol y progesterona secretadas por la placenta. Las células glándulares se desarrollan y se convierten en células secretorias, y se desarrollan también los conductos y alveolos de las glándulas mamarias. A pesar de sus notables efectos en le desarrollo de los pechos, el estradiol y la progesterona juntos inhiben la producción real de leche. Después del parto hay una considerable reducción de la concentración de progesterona circulante en la sangre materna y cesa la inhibición y producción de leche. Durante el embarazo la producción de gonadotropinas hipofisiarias ha sido inhibida por las gonadotropinas placentarias. Con la desaparición de la placenta y sus hormonas, la hipófisis comienza a secretar grandes cantidades de hormona luteotrópica (prolactina) que estimula los pechos para producir leche. Si los pechos se vacían de leche regularmente, continúa la producción de prolactina por la hipófisis. Si no se retira la leche de los pechos, cesa la producción de prolactina en la hipófisis y se detiene la producción de leche por los pechos. El mecanismo de ésto (fig. 25−19) consiste en impulsos nerviosos que pasan de los pezones, por la médula espinal, al hipotálamo, el cual produce LTRF, factor liberador de hormona luteotrópica. Este pasa por el sistema portal hipofisiario a la hipófisis y estimula la secreción de prolactina. Si los pezones no son estimulados por succión, no se secreta LTRF ni prolactina y cesa la producción de leche. La liberación de leche por la glándula mamaria está sujeta a otro sistema de control hormonal. Cuando un ni_o succiona un pezón, generalmente no obtiene leche en el primer minuto. Luego, aparece leche en los conductos de ambos pechos, aunque la succión se halla producido sólo en uno de ellos. El hecho de que aparezca leche en ambos pechos indica que se ha producido cierto proceso estimulador general, en vez de uno local confinado al pecho succionado. Los impulsos nerviosos iniciados por la succión del pecho pasan, 19 atravesando la médula espinal, al hipotálamo y estimulan la liberación de vasopresina y oxitocina. La oxitocina es mucho más eficaz que la vasopresina en producir evacuación de leche. La oxitocina circula en la sangre hasta los pechos y estimula la contracción de las células mioepiteliales que rodean los alveolos, elevando la presión dentro del pecho y exprimiendo la leche de los alveolos hacia los conductos que conducen a los peones. La insulina y el cortisol desempe_an papeles secundarios en regular el desarrollo de los pechos y la secreción de leche. REGULACIÓN HORMONAL DE LAS INTENSIDADES METABÓLICAS La supervivencia de un mecanismo complejo como el hombre requiere, entre otras cosas, mantener el orden entre la multitud de posibles reacciones metabólicas. Cada una de estas reacciones debe aumentar o disminuir de forma apropiada como respuesta a cambios específicos en el medio ambiente. En parte, estos cambios son resultado de las propiedades cinéticas de las enzimas: las intensidades aumentan o disminuyen cuando las concentraciones de substratos y cofactores se elevan y descienden. Por ejemplo, la glucosa 6−fosfato puede convertirse en el hígado en glucosa 1−fosfato, en fructosa−6−fosfato, en 6 fosfogluconato o en glucosa libre. Cada reacción es catalizada por una enzima específica y la velocidad de cada una es controlada por la afinidad de la enzima por la glucosa−6−fosfato y por la velocidad máxima de la enzima cuando está totalmente saturada de substrato. Simplemente cambiar la concentración de glucosa−6−fosfato presente cambiará la dirección en que fluye le metabolismo y no simplemente la intensidad de él. En bajas condiciones de glucosa−6−fosfato, que tiene la máxima afinidad por el substrato y la mayoría de la peque_a cantidad de glucosa−6−fosfato presente se convertirá en ácido−6−fosfoglucónico. Así, con esta baja concentración de substrato, la glucosa−6−fosfatasa, que tiene una afinidad muy baja por el substrato, difícilmente operará y poco o nada de la glucosa−6−fosfato será convertida en glucosa libre. Con altas concentraciones de glucosa−6−fosfato, cuando todas las enzimas están saturadas de substrato, los factores que determinan la dirección del flujo metabólico con las velocidades máximas relativas de las cuatro enzimas. La glucosa−6−fosfatasa tiene una alta velocidad máxima y en altas concentraciones de glucosa−6−fosfato la mayor parte del substrato es convertido en glucosa. Además de los efectos de las concentraciones de substrato y cofactores, la actividad de muchas enzimas es modulada por la presencia de otras moléculas. La actividad de fosfofructocinasa, por ejemplo, que convierte fructosa−6−fosfato en fructosa−1, 6−diosfato, se incrementa por el ácido adenílico, AMP pero es inhibida por ATP. La enzima que cataliza la conversión de fructosa−6−fosfato, la fructosa difosfatasa, es inhibida por AMP. Los efectos opuestos del AMP sobre las dos enzimas impiden que ambas sean activas al mismo tiempo, lo que resultaría en una inútil hidrólisis cíclica de ATP. El tema de la modulación de las enzimas y los detalles de estos cambios en la actividad enzimática son muy complejos. Superpuestos a los controles que dependen de las propiedades innatas de las enzimas y sus respuestas a los metabolitos que modulan su actividad hay otros cambios inducidos por hormonas específicas. Varias hormonas −tiroxina, insulina, hormona del crecimiento y cortisol− producen notables efectos sobre el metabolismo de una amplia variedad de tejidos. Estos controles ayudan también a mantener el orden entre las numerosas posibles reacciones metabólicas en dichos tejidos. Muchos vertebrados, por ejemplo, no alcanzarán su forma y sus dimensiones del estado adulto normal en ausencia de la tiroxina. La tiroxina y la hormona del crecimiento actúan sinérgicamente en la promoción del crecimiento normal del esqueleto. El efecto de la tiroxina en el control de la metamorfosis de los anfibios es muy conocido y la tiroxina afecta al crecimiento y la diferenciación de otros vertebrados, aunque no en forma tan destacada. La tiroxina y la triyodotironina aceleran la intensidad metabólica general y el consumo de oxígeno de casi todos los órganos y tejidos del cuerpo. Se ha informado que las actividades de más de 100 enzimas aumentan después de la administración de tiroxina. Como esta aumenta el consumo de oxígeno y este gas es utilizado principalmente por las mitocondrias, era natural buscar un efecto primario de la tiroxina en la estructura y función de las mitocondrias. Las mitocondrias aisladas se hinchan cuando se exponen a la tiroxina. Aunque concentraciones muy altas de tiroxina pueden desacoplar la fosforilación oxidativa, de modo 20 que se libera energía como calor, en vez de ser retenida en enlaces fosfato ricos en energía, este fenómeno probablemente no guarda relación con los aspectos fisiológicos de la tiroxina. El aumento de la intensidad del metabolismo basal de un individuo hipertiroideo no puede explicarse por un efecto en cualquier proceso metabólico. La insulina regula el metabolismo de las proteínas y los lípidos, así como el de los carbohidratos. La insulina facilita la entrada de glucosa en el músculo esquelético y en el tejido adiposo ; incrementa también la actividad de algunas enzimas, como glucocinasa y sintetiza glucógeno. La entrada de aminoácidos en las células y la incorporación de aminoácidos a las proteinas son favorecidos por la insulina. Esta aumenta la conversión de glucosa en ácidos grasos e inhibe la hidrólisis de triglicéridos en tejido adiposo. El cortisol produce efectos metabólicos opuestos a los de la insulina. Provoca una movilización de aminoácidos desde los tejidos periféricos y acelerá la gluconeogénesis, la conversión de las cadenas de carbonos de los aminoácidos en glucosa y glucógeno. El cortisol inhibe la utilización de glucosa por el músculo esquelético y otros tejidos periféricos, y acelera la movilización de ácidos grasos, elevando la velocidad de producción de cuerpos cetónicos. La hormona del crecimiento produce una multitud de efectos en los tejidos del cuerpo, favoreciendo la transferencia de aminoácidos en las células y aumentando la síntesis de proteinas. Aumenta la síntesis de ADN y ARN en el hígado y músculo, y la síntesis de colágena y mucopolisacáridos en el tejido conectivo y la piel. Una deficiencia de la hormona del crecimiento aumenta la sensibilidad del individuo a la insulina, y una dosis dada de insulina produce una disminución mayor que la normal de la concemtración de glucosa en la sangre. La hormona del crecimiento reduce las concentraciones de urea y aminoácidos en la sangre, reflejando el mayor consumo de aminoácidos y su utilización en la síntesis de las proteinas. La hormona del crecimiento reduce la velocidad de conversión del nitrógeno de los aminoácidos en urea. Estimula la movilización de grasa del tejido adiposo y aumenta la concentración de ácidos grasos en el plasma. Muchos de los efectos de la hormona del crecimiento contrarrestan los de la insulina. MECANISMOS DE ACCIÓN DE LAS HORMONAS Toda teoría del mecanismo molecular por le cual una hormona dada produce sus efectos específicos en tejidos específicos debe explicar el alto grado de especificidad de muchas hormonas y la notable amplificación biológica inherente a los procesos hormonales. Las hormonas circulan por la sangre en concentraciones muy bajas. Las distintas teorías actuales relativas al mecanismo de la acción hormonal sugieren por igual que la hormona se dirige hacia una célula o penetra en ella y se combina con cierto receptor específico. Muchas células no reconocen ciertas hormonas, presumiblemente porque no tienen receptor para dicha hormona y porque los receptores son altamente específicos. Difieren las hipótesis en cuanto a cuál es el receptor dónde está situado y que sucede después de que la hormona se liga a su receptor. Experimentos apropiados con hormonas esteroides de alto título han demostrado la presencia de receptores proteínicos específicos para el estradiol en el útero, para la progesterona en el oviducto, para la aldosterona en el riñon y para la testosterona en la glándula prostática. Los efectos de las hormonas en facilitar la entrada de ciertas sustancias en la célula, como el consumo de la glucosa por las células musculares estimulado por la insulina, ha sugerido que la hormona se combina con una proteina u otra sustancia en la membrana celular. Esto provoca un cambio en la arquitectura molecular de la membrana y, por tanto, su permeabilidad a sustratos específicos. Las hormonas proteicas tienen su receptor en la membrana celular cuya actividad enzimática es alterada por la combinación. Esto activa una adenilciclasa específica y estimula la producción de AMP cíclico a partir de ATP. El AMP cíclico es considerado como un segundo mensajero que produce el efecto de la hormona. La adrenalina, por ejemplo, se ha demostrado que estimula la adenil ciclasa de las células hepáticas. El AMP cíclico resultante es un activador de otra enzima, una cinasa proteínica, que transfiere un grupo fosfato de 21 ATP a una tercera enzima, fosforilasa cinasa, y la activa de modo que puede convertir una cuarta enzima inactiva, la fosforilasa b, en fosforilasa a activa. Esta última cataliza entonces la producción de glucosa −1− fosfato a partir de glucógeno. En cada una de estas etapas sucesivas hay una amplificación de 10 a 100 veces, por lo que una cantidad muy pequeña de adrenalina producirá una cantidad muy grande de glucosa −1− fosfato. Las hormonas esteroideas debido a su menor tamaño penetran en el núcleo y activan genes específicos que fueron reprimidos anteriormente, por lo que estos se transcriben. Esto conduce a la producción de nuevas clases de ARN mensajero que codifican la síntesis de nuevas proteinas específicas. Esta teoría también explica la marcada amplificación de los efectos hormonales, pues una cantidad pequeña de hormona, girando alrededor de la transcripción de un gen específico, podría dar por resultado muchas moléculas de ARN mensajero y muchas más de proteina. No está claro si las hormonas son consumidas comunmente cuando regulan el metabolismo en sus tejidos blanco. El estradiol no es consumido o cambiado químicamente cuando estimula el crecimiento del útero. Las hormonas ligadas a sus receptores son relativamente estables, pero las hormonas que circulan en la sangre tienen periodos de semidesintegración biológica relativamente cortos, Son inactivadas y eliminadas del cuerpo, y deben ser sustituidas por nuevas moléculas hormonales sintetizadas en la glándula endocrina apropiada. Parece improbable que todas las hormonas tengan un mecanismo molecular común por el cual se producen sus efectos. Ciertamente, hay pruebas de que algunas hormonas producen sus efectos no por un solo mecanismo, sino por diferentes mecanismos que actúan al unísono. Las teorías aceptadas en un momento dado tienden a reflejar nuestros conocimientos generales de la biología celular y molecular. Teorías que suponen efectos de las hormonas sobre las membranas celulares han dado paso a teorías sobre un efecto de alteración de la actividad de una enzima y estas, a su vez, han sido sustituidas por teorías referentes a efectos de la hormona sobre el mecanismo genético, que da por resultado la síntesis de clases específicas de ARN y proteinas. FEROMONAS En los últimos años se ha llegado a la convicción de que la conducta de los animales puede recibir influencia no sólo de las hormonas, cuerpos químicos liberados en el medio interno por las glándulas endocrinas, las cuales regulan y coordinan las actividades de otros tejidos, sino también de las feromonas, sustancias secretadas por glándulas exocrinas, liberadas en el medio externo y que ejercen su influencia sobre la conducta de otros animales de la misma especie. Hasta ahora se admitía la posibilidad de transferencia de información de un animal a otro por la vista o el sonido ; las feromonas representan un medio de comunicación o de transferencia de información valiendose del olfato. Las feromonas provocan respuestas específicas de conducta, desarrollo o reproducción en el receptor ; estas respuestas pueden ser de gran importancia para la supervivencia de la especie. Algunas feromonas actúan en forma desconocida sobre el sistema nervioso central del receptor y producen efecto inmediato sobre su conducta. Cabe citar entre ellas la de atracción sexual, las feromonas de huella y rastreo y sustancias de alarma secretadas por las hormigas. Otras feromonas actúan más lentamente y desencadenan en el receptor una serie de acontecimientos fisiológicos eslabonados que afectan su crecimiento y diferenciación. Hay ejemplos de feromonas en insectos, como por ejemplo en el caso de algunas clases de hormigas. Cuando regresan al nido después de encontrar comida, secreta una feromona de huella y rastreo que marca el camino para que las restantes hormigas puedan encontrarlo facilmente. Esta feromona es volatil y se evapora en dos minutos, de manera que es casi imposible que puedan confundirse otras hormigas con viejos rastros. Las hormigas también liberan sustancias de alarma ante el peligro que a su vez transmiten la señal a las 22 compañeras en las cercanías. Estas sustancias tienen peso molecular inferior al de las feromonas de atracción sexual, y son menos específicas, de forma que responden a la misma sustancia de alarma miembros de diferentes especies. Las abejas obreras, cuando encuentran alimento, secretan geraniol, que atrae a otras obreras hacia el hallazgo. Estas completan la información valiendose de su danza típica. Las abejas reinas secretan ácido 9−cetodecanoico, el cual, ingerido por las obreras, inhibe el desarrollo de sus ovarios y su capacidad para formar células reales que pudieran dar nacimiento a nuevas reinas. Esta sustancia actúa también como feromona de atracción sexual para la abeja macho durante el vuelo nupcial de la reina. En los insectos que organizan colonias, como hormigas, abejas y termitas, las feromonas desempeñan un importante papel como reguladoras y coordinadoras de la composición y actividades de la población. En la colonía de termitas es distinta la morfología de la reina, el rey, soldados y ninfas u obreros. Todos provienen de huevos fecundados ; sin embargo, las reinas, reyes y soldados secretan sustancias inhibidoras, feromonas que actúan sobre el cuerpo alado de las ninfas e impiden su desarrollo en tipos más especializados. Si la reina muere, durante cierto tiempo no se libera feromona antirreina de modo que una o más de las ninfas se convierten en reinas. Los miembros de cada colonia sólo permitirán la supervivencia de una reina y devorarán a las demás. En forma análoga, la pérdida del rey termita o la disminución del número de soldados provoca automáticamente la transformación de otras ninfas en castas especializadas para reemplazarlos. Los machos del saltamontes migratorio secretan una sustancia por la superficie de su piel que acelera el crecimiento de las crías. Hay ejemplos de feromonas en mamíferos, igual que en insectos. Cuando se colocan cuatro o más ratones hembra por jaula aumenta notablemente la frecuencia de seudoembarazos, pero este efecto desaparece si se extirpan sus bulbos olfatorios. Al situar varias hembras juntas en una jaula se observa irregularidad en sus ciclos estruales. Ahora bien, si se introduce un macho en esta jaula su olor puede iniciar y sincronizar los ciclos de todas las hembras (efecto Whitten) y disminuir la frecuencia de anomalías reproductivas. Todavía más interesante es el hallazgo (efecto Bruce) de que el olor de un macho extraño bloquea el embarazo de una ratona recientemente fecundada. Los impulsos nerviosos procedentes de la nariz pasan al hipotálamo y bloquean la producción del factor liberador de prolactina. La consiguiente falta de prolactina provoca la regresión de los cuerpos amarillos y la falta de implantación de los óvulos fecundados. En cuanto a las feromonas humanas todavía no se ha podido concluir nada definitivo al respecto, pero parece existir un cierto olor corporal personal que puede ser más agradable o menos según de quién hablemos. También a la hora de las relaciones sexuales parece aumentarse el nivel de hormonas sexuales y producirse un cierto tipo de feromona en forma de olor que ayudaría a que dichas relaciones llegaran a fin. Un dato concreto es el que parece derivarse del olor del ácido 15−hidroxipentadecanoico que sólo es percibido por la mujer madura y con mayor incidencia durante la ovulación. Los varones y las niñas son relativamente insensibles a esta sustancia, pero los primeros adquieren sensibilidad a la misma después de una inyección de estrógenos. Análisis de los ciclos menstruales de las estudiantes de una universidad femenina norteamericana demostraron una tendencia estadísticamente significativa en favor de la creciente sincronización delos ciclos menstruales entre compañeras de habitación y amigas íntimas. El estudio descartó varias posibles explicaciones del fenómeno, pero sugirió cierto efecto feromonal entre jóvenes que estaban juntas gran parte del tiempo. El estudio demostró también un acortamiento importante de los ciclos menstruales de mujeres que estuvieron con compañeros tres o más veces a la semana ; las que se veían con ellos dos veces a la semana o menos tenían ciclos más largos e irregulares. Esto parece ser análogo al efecto Whitten, pero se desconoce si se debe a una feromona. LOS BIORRITMOS Los biorritmos son representaciones, gráficas o digitales, de fuerza de estímulo químicas y sensoriales que 23 varían sobre una base periódica. Como estas influencias desencadenan numerosas funciones corporales, la técnica para determinar la intensidad y duración de los ciclos constituye un método para calibrar las potencialidades relativas de rendimiento del cuerpo en los distintos momentos durante los ciclos. Los estímulos químicos son las hormonas (de distintos tipos y en distintas cantidades) segregadas por las glándulas endocrinas del organismo. Las acciones periódicas de dichas glándulas son controladas por el SNC, pero las actividades químicas son autónomas. Como tales, son extremadamente fiables, a menos que exista una enfermedad del sistema nervioso central o una privación exagerada de proteinas en la alimentación durante un prolongado periodo de tiempo. Los estímulos sensoriales son los estados periódicos de la naturaleza que se producen fuera del organismo, pero sincronizándose siempre con los estímulos químicos. Los estados alternativos de luz−oscuridad, calor y frío, etc, constituyen ejemplos de condiciones externas a las que el organismo responde automáticamente. Los biorritmos son como marcadores de señales del comienzo de cada fase, o de su máximo o mínimo. Nuestros organismos se sincronizan con correlativos externos y fijan las fases y ritmo de los acontecimientos celulares y holobiológicos (Dr. Stanley R. Glasser, de la Facultad Baylor de Medicina). Clasifica además los relojes biológicos en centrales, homeostáticos y periféricos : Los relojes centrales operan a un ritmo perfecto (sin dependencias externas) y están situados en el hipotálamo. Modulan numerosas funciones autónomas y respuestas holobiológicas sin servomecanismos (proceso biológico que se comporta como mecanismo autorregulador). Los relojes homeostáticos son menos exactos, pues varían al mantener correlaciones con influencias externas. El ciclo del estrus (o menstruación) es el biorritmo homeostático más conocido. Las investigaciones médicas sobre las causas y efectos de ciclos de distinta duración son muy amplias, especialmente en el campo de la endocrinología. Los investigadores no conocen aun la naturaleza de los relojes o biorritmos, pero sus efectos son perfectamente observables, predecibles y que repercuten sobre las tendencias conductistas humanas. No se trata de órganos físicos y diferenciados, pero pueden ser muy bien las mismas reacciones observadas, tales como los ácidos nucleicos o las proteinas, pero aun no se les ha conseguido aislar. Cualquier deficiencia de una secrección glandular se indica por el prefijo "hipo", mientras que un exceso de secrección se denomina por el prefijo "hiper". Por tanto, el mixedema es un tipo de hipotiroidismo 24