2 Citoesqueleto
Anuncio

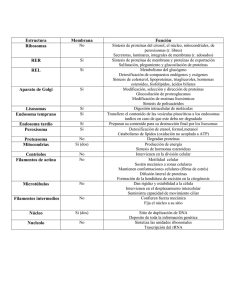
CITOESQUELETO (Organelos filamentosos) DRA. GUÉRNICA GARCÍA Las células epiteliales poseen una trama o malla tridimensional de material fibrilar citoplasmático, llamada citoesqueleto, que interconectan al núcleo, a los organelos y a la membrana celular. Esta trama permite a la célula mantener su forma y textura, además de participar en procesos de organización, diferenciación, división, motilidad, secreción y absorción. El citoesqueleto está constituido por cuatro clases de elementos fibrilares: Ø Ø Ø Ø a) Microfilamentos. Filamentos intermedios. Filamentos gruesos (no presentes en células epiteliales, sí en fibras musculares) Microtúbulos. Los microfilamentos miden 5 a 7 nm de diámetro. Se ubican preferentemente en la periferia celular sosteniendo organelos. Se constituyen por una malla de actina y otras proteínas que cumplen funciones de mantención del orden interno de organelos, de anclaje entre células y de contracción. Los filamentos de actina tienen disposición en espiral. Se encuentran en gran cantidad en las uniones adherentes y en el interior de las microvellosidades b) Los filamentos intermedios reciben este nombre por poseer un diámetro de 7 a 11 nm. Todos los filamentos intermedios poseen la misma morfología, pero distinta composición química, según el tejido del cual se trate. Esta diferencia se emplea para clasificar tumores y sus metástasis, ya que conservan sus proteínas originales: v v v v v Citoqueratina, en todos los tejidos epiteliales, queratinizados o no. Vimentina, en los tejidos conectivos. Desmina, en tejidos musculares. Neurofilamentos, en las neuronas. Proteína fibrilar glial acídica, en los filamentos gliales de astrocitos. Estos filamentos cruzan la célula de un lado a otro y se anclan a los desmosomas, dando forma y rigidez celular. c) Los microtúbulos tienen un diámetro de 20 a 25 nm y se constituyen por una proteína llamada tubulina (fracciones a y b tubulina). 1 Los microtúbulos participan en el transporte intracitoplasmático de proteínas y vesículas, en el transporte axonal en neuronas, en la división celular (formando el huso mitótico y los centríolos), en la estructura de cilios y flagelos (formando el axonema) y en el transporte de pigmentos como la melanina. DIFERENCIACIONES O ESPECIALIZACIONES EPITELIALES Los tejidos epiteliales presentan como característica, la tendencia a establecer un estrecho contacto entre sus células, con un escaso o nulo contenido de sustancia intercelular, formando láminas compactas que recubren superficies y cavidades. Para cumplir con su función, estas células poseen diferenciaciones o especializaciones de sus caras celulares laterales, basal y apical. ESPECIALIZACIONES APICALES O DE LA SUPERFICIE LIBRE Estas estructuras especializadas, permiten aumentar la eficiencia del epitelio, para sus funciones de absorción y/o transporte: v Microvellosidades: son evaginaciones digitiformes (con forma de dedo), presentes en algunos tipos de epitelios cúbicos o cilíndricos con función absortiva o secretora. Tienen por objeto aumentar la superficie de intercambio entre la célula y su medio (la incrementan hasta en 20 veces). Mediante el empleo del microscopio óptico, se describió la presencia en la superficie apical de ciertas células, una delgada línea refringente, que fue llamada borde estriado o en cepillo. Mediante el empleo de microscopía electrónica se describieron en forma exacta estas estructuras. Las microvellosidades (M.V.) son expansiones celulares cilíndricas y múltiples (hasta 2.000 por célula), de 80 a 90 nm de diámetro y 1 a 2 mm de longitud, cubiertas por membrana plasmática y envueltas más externamente por el glicocalix. El Glicocalix es una capa especializada que se ubica en la superficie libre o apical de todas las células epiteliales. El glicocalix corresponde a oligosacáridos terminales de las proteínas integrales de membrana, que sobresalen de la membrana celular. Esta capa resulta fundamental en el reconocimiento de sustancias, actuando como receptores para nutrientes y elementos inmunológicos. Tiene por lo tanto, importancia en la nutrición y en la defensa inmunológica. En el interior de las M.V. existen entre 20 y 30 filamentos de actina y pequeñas proteínas (villina y fimbrina), unidos en la base al citoesqueleto, que presenta en esta zona una orientación transversal, también conocido como velo terminal. Estos filamentos tienen por función otorgar rigidez a la estructura, manteniendo la disposición paralela entre una y otra. 2 v Cilios: corresponden a proyecciones celulares móviles, muy numerosas (50 a 100 por célula), de largo considerable (7 a 20 mm), pudiendo coexistir con microvellosidades. Son observables a M.O. En el interior de cada cilio se encuentra una compleja estructura interna, llamada axonema, esta estructura contiene un par de microtúbulos centrales y nueve dobletes periféricos, que se extienden a lo largo del cilio, hasta implantarse en un cuerpo basal, semejante a un centríolo, el que presenta nueve tripletes de microtúbulos dispuestos en espiral. Este cuerpo basal se ubica en la base de cada cilio. El movimiento ciliar se debe a la acción de las proteínas dineína y proteasa flagelar; estas proteínas ensamblan un microtúbulo con otro. A medida que las dineínas se contraen, los microtúbulos se deslizan uno sobre el otro, moviendo al cilio. El pH del medio influye sobre las dineínas, un aumento o disminución de él, así como el frío, inmoviliza estas estructuras. Los cilios realizan complejos movimientos oscilatorios, rápidos en una dirección fija y lentos en el sentido contrario, mediante los cuales barren la superficie epitelial, por esto se encuentran en células especializadas en transporte de líquidos, partículas o secreción mucosa. Se encuentran en los epitelios que tapizan el aparato respiratorio y en el genital de las hembras. El espermatozoide presenta una estructura flagelar similar a la de los cilios. Este flagelo es más largo y se observa uno por cada célula. v Estereocilios: estas estructuras son largas proyecciones citoplasmáticas, de células altas, ya que requieren buena base de sustentación. Su base es ancha, y no poseen movimiento propio. Pueden ser observadas al M.O. en la superficie del epitelio. Cada célula presenta un penacho piriforme, con múltiples estereocilios, muy largos (15 mm) y flexibles. En la región basal son paralelos y se entrelazan en los extremos. Cada estereocilio está cubierto por membrana celular y presentan una manojo central de filamentos de actina, que se anclan en el citoesqueleto. La función de estos estereocilios no es clara, pero contribuyen a aumentar la superficie de intercambio, creando un microambiente que hace más eficiente la función del epitelio epididimario. 3 ESPECIALIZACIONES DE LA MEMBRANA BASAL En el límite entre los tejidos epiteliales y el tejido conectivo que lo sustenta, existe una estructura llamada lámina o membrana basal (M.B.). La superficie basal de la mayoría de los epitelios es lisa, pero en los epitelios de transporte pueden existir pliegues que aumentan la superficie de la membrana basal. · Estructura de la membrana basal: . La membrana basal es una estructura laminar, siempre presente en los epitelios. Esta membrana es elaborada por las células epiteliales y en menor medida por los fibroblastos presentes en el conectivo. Se ubica en el límite de la célula epitelial, separándola del tejido subyacente. No es visible con la técnica de tinción de H-E. Se tiñe intensamente de color negro, con tinción de plata. Al microscopio electrónico se observan dos zonas: la lámina rara o lúcida, de baja densidad electrónica y la lámina densa, de mayor densidad electrónica. Adosada a estas dos zonas se observa una capa contigua al tejido conectivo adyacente, la lámina reticular o fibroreticular. La lámina lúcida, en contacto con la membrana plasmática de la célula epitelial, presenta un aspecto homogéneo y se encuentra constituida por glicoproteínas (laminina, entactina y fibronectina) y por condroitín y heparán-sulfato, dos proteoglicanos. La lámina densa, aparentemente amorfa, está compuesta por colágeno tipo IV, el cual constituye una densa red de filamentos orientados al azar, de 4 nm de diámetro. La lámina reticular contiene innumerables fibras de anclaje correspondiente a colágeno tipo III y cantidades variables de fibronectina. Representa la zona de unión entre el epitelio y el tejido conectivo subyacente. 4 · Funciones de la membrana basal: su función es proporcionar un soporte físico al epitelio, contribuye a la adhesión celular, intervenir en la ultrafiltración en endotelios capilares y participar en la reparación celular. Durante el desarrollo embrionario actúa como sustrato para la migración celular e influye en la diferenciación celular, mediante interacciones inductivas con las células adyacentes. En la vida postnatal, sirve de base para la reparación celular, siendo guía para la migración de células neoformadas, en los márgenes de las soluciones de continuidad que hay que reparar. La presencia de M.B. limita la migración de células epiteliales en proliferación. En histopatología se considera a una neoplasia como benigna (adenoma, epitelioma), cuando las células tumorales no atraviesan esta membrana. En caso contrario, si la M.B. es discontínua o no se visualiza, se considera a la neoplasia como maligna o invasiva (carcinoma), observándose comúnmente metástasis de células tumorales. ESPECIALIZACIONES DE LA SUPERFICIE LATERAL En las superficies laterales de las células epiteliales se diferencian estructuras específicas interdigitadas, que las mantiene firmemente unidas entre sí. Estas especializaciones permiten la adhesión y la comunicación celular. La microscopía óptica, desde hace muchos años, evidenció en intestino, la presencia intercelular de zona oscuras localizadas bajo la superficie apical; estas zonas se llamaron barras terminales. Mediante el empleo del microscopio electrónico, se determinó que esas zonas correspondían a complejos de unión, los cuales son una sucesión ordenada de medios de unión altamente especializados: · · · La zónula occludens o unión estrecha o tight junction. La zónula adherens o unión adherente. La mácula adherens o desmosoma. Un cuarto tipo de medio de unión existente entre dos células epiteliales, es la unión comunicante (también conocida como unión de fisura, nexo, nexus o gap junction). Esta unión permite la comunicación entre células y no constituye parte del denominado complejo de unión. 5 1) La zónula occludens se encuentra inmediatamente bajo la superficie libre del epitelio. Es una unión ocluyente que constituye una especie de cinturón o anillo, de 0,1 a 0,3 mm, que abarca todo el perímetro de la célula, asegurando de esta manera la completa obliteración o cierre del espacio intercelular. En esta zona se originan múltiples puntos de fusión de las hojas externas de las membranas celulares involucradas, mediante la unión de partículas proteicas de las membranas celulares adyacentes (proteínas integrales ocludinas y claudinas), conformando filamentos rectilíneos anastomosados, que obliteran el espacio extracelular. Entre las zonas de fusión, las membranas están separadas por 10 a 15 nm de distancia. Esta adhesión celular, con forma de bandas circunferenciales en el extremo apical de la célula, impide que las moléculas puedan atravesar el epitelio por el espacio extracelular, a la vez que juega un rol mecánico en el mantenimiento de la integridad celular. Esta unión es de particular importancia en epitelios simples o monoestratificados. Existen factores tales como la disminución en la concentración de calcio, variaciones de pH o traumatismos, que alteran este tipo de unión, desprendiendo las membranas. 6 2) La zónula adherens se ubica por debajo de la zónula occludens. A este nivel las membranas se encuentran paralelas y separadas por un espacio de 15 a 20 nm, ocupado por un material amorfo de baja densidad electrónica. Esta estructura rodea a la célula a modo de banda o cinturón, en forma paralela a la unión ocluyente. Esta compuesta por una densa red de material filamentoso citoplasmático en el que se insertan microfilamentos de actina, que refuerzan la superficie de la membrana celular y constituye una banda de adhesión firme, entre células vecinas. En este tipo de unión, las proteínas integrales de membrana cateninas y E-cadherinas, se unen entre sí y permiten el anclaje de elementos del citoesqueleto. 3) La mácula adherens o desmosoma, es el tercer elemento del complejo de unión. Es una estructura discoidal, de 0,5 mm de diámetro. En este sector las membranas celulares se encuentran separadas por un espacio de 20 nm, el que se encuentra ocupado por matriz extracelular compuesta por glicoproteinas, glicolípidos y proteínas integrales del tipo cadherinas (desmogleina y desmocolina). En la zona intercelular central se encuentran filamentos finos, que divide en dos el espacio existente y conforman una placa densa, la denominada bisectriz. 7 La superficie citoplasmática de la membrana celular se encuentra engrosada, constituyendo la placa de adhesión (constituida por las proteínas placoglobina y desmoplaquina), en la cual se anclan los filamentos intermedios de citoqueratina del citoesqueleto. Los filamentos al contactar con esta zona se doblan en horquilla y retornan al citoplasma. Se produce entonces, una integración total entre las estructuras intra y extracelulares. Estas características convierten a este medio de unión en la forma más eficaz y resistente de anclaje entre células. Permiten mantener la unión, la forma y la rigidez celular. La integridad de los desmosomas depende de la presencia de calcio, en su ausencia se separan las dos mitades del desmosoma (hemidesmosomas). La formación de un hemidesmosoma induce la formación de la mitad complementaria en la célula vecina. Sin embargo, se encuentran hemidesmosomas en la superficie basal del epitelio. Los desmosomas también se pueden encontrar dispersos sin formar parte del complejo de unión. Se ubican especialmente entre células constituyentes de epitelios pluriestratificados. 4) La unión comunicante (también conocida como unión de fisura, nexo, nexus o gap junction) es una unión discoidal en la cual las membranas celulares involucradas quedan separadas en toda su extensión, por una distancia de 2 a 3 nm, sin llegar a fusionarse nunca. Las membranas plasmáticas de ambas células, son atravesadas por varios “puentes”. Estos puentes, en cada célula, se constituyen por seis subunidades globulares protéicas denominadas conexinas, que se disponen hexagonalmente, dejando una abertura central o poro de 1,5 a 2 nm de diámetro. Gracias a la alineación y a la unión de ambos extremos de los puentes de ambas células, se forman unidades como canales, llamadas conexones. 8 A través de estos canales se produce un flujo de iónes y de moléculas, lo que permite un acoplamiento eléctrico en células epiteliales y musculares, entre otras, frente a un estímulo hormonal o nervioso. Esta estructura se encuentra dispersa y en número variable en las células epiteliales, dependiendo de la función que cumplan. 9