Genética del Color del Plumaje en Palomas: Colombicultura
Anuncio
El COLOR DEL PLUMAJE EN COLOMBICULTURA I (J.Rubio. Sevilla, 2004) Uno de los principales retos que surge a la hora de escribir sobre los colores de las palomas es con la diversidad de nombres y calificativos que se utiliza en distintas regiones para describir una misma característica (un prieto en Sevilla es lo mismo que un magaño en Murcia o un Pizarro en Almería); en otros casos la misma palabra tiene significados distintos y opuestos según la región donde se trate, (bayo en Sevilla equivale a rojo mientras que en Jaén equivale a marrón), por lo que hemos usado la terminología mas comúnmente utilizada e intentado describir e ilustrar cada una de las propiedades. Aunque se trata de describir de una forma amena y coloquial la herencia de los colores en las palomas, en algún momento, puede que tenga que volver a echarle un vistazo a la parte de “Algo sobre Genética” en www.colombicultura.com para poder interpretar adecuadamente lo que se expone. El termino "tipo salvaje" o “wildtype” se usa en genética para indicar un ejemplar con respecto al cual comparar el resto, esto es, el tipo de paloma que comúnmente se ha aceptado como patrón base, y que en nuestro caso es la paloma “Zurita”. El “wildtype” representa una paloma azul, con dos barras negras en la cobertura de las alas, barra oscura en el borde de la cola, plumas extremas de la cola con borde blanco, de tamaño normal, pico negro, patas sin plumas, sin cresta, ojos anaranjados, etc. En otros términos, representa un pájaro semejante a la paloma zurita “columba livia”. Pues bien, cada vez que nos refiramos a alguna propiedad genética que esté visible en la wildtype o zurita, la representaremos con el signo (+), así para definir el pigmento de una paloma wildtype o zurita lo escribiremos como (+/+) ó (B+/B+) La apariencia que presenta una paloma en cuanto al colorido y la forma de distribución del mismo, lo que de forma coloquial los palomeros llaman “color” (por ejemplo: azul gotado, prieto aliblanco, gabino rojo, etc.) es consecuencia de la combinación de una serie de factores (asociados a pares genéticos) que nosotros de una forma simplificada clasificaremos en tres: pigmento, patrón y modificadores. EL PIGMENTO En las palomas existen tres tipos de melanina que son las responsables del pigmento de nuestras palomas, estos son: “marrón”, “negro o azul” y “rojo” en orden ascendente de de dominancia, es decir, el azul es dominante sobre el marrón y el rojo sobre el azul y el marrón. No hay que olvidar un cuarto pseudo-pigmento, o mejor dicho, ausencia de pigmento, el “blanco“. De forma simplificada y general, podemos decir que cualquier característica de la palom@ esta asociada a dos celdas genéticas donde se ubican sendo genes. Si un palom@ tiene el mismo gen en ambas celdas se considera "homocigótica" para esa característica y en caso contrario, es decir, que en una celda tengamos un gen y en la otra tengamos uno de sus alelos, decimos que es heterocigótica. Los genes que determinan el pigmento de una paloma están ubicados en el cromosoma del sexo, y este a su vez es distinto en su longitud para los machos y para las hembras así mientras en los machos existen dos celdas genéticas para el pigmento, en las hembras sola tienen un celda, la otra está atrofiada, (de esto se dio cuenta un americano llamado Thomas H. Morgan). Así podemos deducir que en el caso de las hembras son siempre homocigóticas para el pigmento, puesto que solo tienen un gen que lo determina. En función del tipo de melanina que presente el pigmento podemos clasificada según orden de dominancia descendente en: (BA) rojo ceniza, (B+) negro ó azul y (b) marrón. Como hemos dicho, las hembras tienen un solo gen que determina su pigmento, (la representación genética de una hembra sería del tipo (b/.) para el marrón, (B+/.) para el azul y (BA/.) para el rojo ceniza). Los machos, por el contrario, tienen dos genes que determinan el pigmento, así por ejemplo se usa (B+/B+) para el azul homocigótico; (B+/b) para el azul llevando marrón; (b/b) para el marrón homocigótico, (BA/b) para el rojo llevando etc. Vamos a describir los pigmentos, intentando hacer una descripción de forma que podemos identificar el color en nuestras palomas, pero no podemos olvidar que en muchos casos, dichos pigmentos no son visibles, están ocultos por otros genes o “modificadores” y para conocer el pigmento utilizaremos los métodos descritos más adelante; en otros casos, el resultado visible puede parecernos un pigmento, pero en el fondo sea otro (lo que habitualmente conocemos como “rojo” puede no ser un rojo ceniza, sino un azul con el gen índigo homocigótico, o un azul con el gen rojo recesivo). Para averiguar el verdadero pigmento de una paloma nos apoyaremos siempre en el fenotipo que presenta y en la ascendencia y la descendencia del palom@ en cuestión. (B+) Azul, (Blue; black). Representado por (+) ó (B+), el pigmento azul es un carácter el cual se manifiesta como un color gris que se puede dar en varias intensidades (o patrones como veremos más adelante) presentando las palomas aspectos distintos. Wildtype Azul (B+/B+)(C+/C+) Siendo recesivo sobre el rojo ceniza y dominante respecto al marrón, el pigmento o melanina azul lleva aparejado una serie características como son: - Borde de las alas oscuros - Barra oscura en el borde final de la cola - Plumas extremas de la cola con borde blanco - Pico negro - Ojos anaranjados Marrón con patrón barrado (b/b)(C+/C+) propiamente dichos, gabinos rojos, etc. El pigmento o melanina roja lleva aparejado una serie características, la más evidentes y claras son: - Cola Ceniza sin barrar - Ojos anaranjados - Bordes de las alas de tonalidad clara Su nomenclatura BA viene de Brown (en mayúscula) es decir marrón dominante, con el superíndice A (en mayúscula) indicando un alelo “Ask” ó “ceniza”. El hecho de que al rojo se le asigne la misma letra aunque en mayúscula que al marrón, es precisamente por ambos genes “b” y “B” son genes alélicos ubicados en la misma posición o celda genética del cromosoma del sexo. LOS PATRONES Azul con patrón checker oscuro ó prieto (B+/B+)(Cd/Cd) En otros términos, el pigmento azul representa una paloma semejante a la paloma zurita “columba livia”; entre los que se incluyen los típicos “azules claros”, “prietos, pizarros ó magaños”, “ahumados ó fumados”, ”negros” etc. (b) Marrón (brown) El marrón es un carácter que causa una transformación en las áreas negras en una paloma de "wildtype" para convertirse en “castaño ó pardo”; puede darse en varias intensidades llegando a ser en algunos casos, muy parecido al rojo. El pigmento o melanina marrón lleva aparejado una serie características como son: - barra oscura en el borde final de la cola - ojo rosáceo de falsa perla distinto del naranja del tipo salvaje - las plumas marrones tienden a blanquear con la exposición continuada al sol, habitualmente hacia el exterior de la pluma, a la inversa que el wildtype que se oscurece en el borde. Marrón con patrón prieto (b/b)(C+/C+) La nomenclatura (b) viene del inglés “brown” que significa “marrón o castaño”, escribiéndose en minúscula para indicar que es recesivo respecto al azul y al rojo, y por lo tanto, para que sea visible tiene que ser homocigótico, es decir, estar por duplicado en sendos genes del par genético (se tiene que haber heredado de ambos progenitores). En este apartado se incluyen los típicos “castaños ó marrones”, “pulevas”, “melados” etc. (BA) Rojo ceniza (Brown ASk) Existen diversos genes que interactuando con los tres pigmentos básicos dan como resultado palomas de apariencia “roja”. A nivel de pigmentación de la pluma solo consideramos el rojo ceniza, siendo el resto de genes “rojos” modificadores independientes del pigmento y que merecen un estudio aparte. El rojo ceniza es un carácter que causa una transformación en las áreas negras de la paloma "wildtype" para convertirse en un color marrón-rojizo. Puede darse en varias intensidades (o patrones como veremos más adelante). En este apartado se incluyen los, rojos barrados, rojos Los patrones son un conjunto de formas o agrupaciones en las que puede presentarse el pigmento en una paloma. Habitualmente se define por la letra “C” del inglés “checker” Todas las palomas están equipadas con dos genes que definen en patrón; así en función de la intensidad con la que se presente o agrupe el pigmento podemos formar una serie. Su nomenclatura y los posibles alelos, clasificada por el orden de dominancia ascendente son: - (c) barless; con tonalidad clara y sin barras oscuras en la cobertura de las alas - (C+) barrado; con tonalidad clara y barras oscuras en la cobertura de las alas (wildtype) - (Cl) checker claro o gotado; tonalidad clara con pequeñas motas en forma de gotas sobre la cobertura de las alas. (checker ligh) - (C) checker o moteado; tonalidad clara con pequeñas motas en forma de gotas sobre la cobertura de las alas (checker), con cierto equilibrio entre zonas claras y el moteado oscuro. - (Cd) checker oscuro o prieto; predomina el moteado oscuro sobre el claro. (checker dark) - (Ct) terciopelo; es un patrón oscuro homogéneo sin zonas claras (T-pattern) LOS MODIFICADORES - (C?) indica un patrón desconocido para el ejemplar en cuestión, pero que ha de ser igual ó inferior según la tabla de dominancia al de su gen emparejado (terciopelo” es dominante sobre “checker” y este a su vez dominante sobre “gotado”, y así sucesivamente). El patrón es una característica autosomal, por lo tanto está controlado por 2 genes, uno de cada progenitor y por lo tanto, NO es sexable. Una paloma que muestre el patrón (CT/C?) puede llevar cualquiera de los cinco alelos en el gen no visible Azul Checker (B+/B+)(C/C?) (CT/CT) (CT/CD) (CT/CL) (CT/C+) (CT/c). Solo se muestra visible el más dominante, en nuestro caso el CT. Una paloma con patrón prieto puede ser a su vez para el gen no visible ser prieto, gotado, barrado o sin barra. De la misma forma una barrada puede llevar barra o ser barless en el otro cromosoma, pero no puede llevar el gen Terciopelo ni el checker, pues de lo contrario, presentaría visible el patrón terciopelo o checker, por ser dominantes sobre el barrado. Una barless o sin-barra ha ser barless en ambas cromosomas porque el gen barless es recesivo a todos los demás genes del patrón. Ejemplo: - (Ct/Cd) se muestra visible como Ct - (Cd/C+)se muestra visible como Cd - (C+/c) se muestra visible como C+ Los modificadores son pares genéticos que pueden cambiar la apariencia de una paloma afectando al pigmento y al patrón e incluso a otros modificadores, como puede ser el caso de gabinos, aliblancos, figuras, rojos, platas, negros, etc. De entre la multitud de modificadores conocidos vamos a hacer una exposición exclusivamente de algunos de los más frecuentes en nuestras palomas deportivas “de pica y de raza buchona”, sin olvidar que pueden mostrarse yuxtapuestos con otros y el resultado final del color de una paloma sea la suma de (pigmento+patrón+X modificadores) Marrón Barrado (b/b)(C+/C+) Marrón gabino homocigótico (b/b)(G/G) Marrón con Patrón-T o terciopelo (b/b)(Ct/C+) Los modificadores de por si, existen Autosomales (no están situados en el cromosoma del sexo; cuando se opera con ellos la hembra presenta 2 genes), y ligados al Sexo (situados en el cromosoma del sexo; cuando se opera con ellos la hembra presenta un solo gen, el otro se considera neutro o nulo y se representa por un punto “.”). MODIFICADORES AUTOSOMALES Rojo ceniza checker o prieto (Ba/?)(C/C?) Azul Barrado (B+/B+)(C+/C+) Rojo ceniza Patrón-T o Terciopelo (Ba/?)(Ct/C?) (S) Extendido (Spread) Este gen autosomal contribuye a cambiar la apariencia externa de una paloma transformando los patrones barrados y checker, coloreando todas las plumas con tonos homogéneos e intensos. Las palomas con pigmento azul se transforman en negro; las marrones en color chocolate oscuro, y las rojas en ceniza homogéneo. El factor Spread es opaco al patrón; por ejemplo, una paloma azul al tener el gen “spread o extendido” oculta el patrón y se muestra como negro sólido. Aunque oculta el patrón, se hereda de forma independiente de él, por lo que para saber el patrón que porta hay que averiguarlo a base de cruces y observación de la descendencia. No se debe confundir el gen Spread (S) con rojo recesivo (e) pues son totalmente distintos e independientes. Su nomenclatura y los posibles alelos son: - (+) wildtype o no extendido - (S) spread o extendido (G) gabino (grizzle) Grizzle es un gen dominante parcial que decolora la pluma desde el cañón hacia fuera. Esta mutación es independiente y se puede dar con cualquier color y patrón (incluso con otros modificadores). Su nomenclatura y los posibles alelos son: - (G) grizzle o gabino - (GW) gabino blanco - (+) wildtype o No gabino Azul barrado Ahumado (B+/B+)(C+/C+)(sy/sy) Ahumado azul checker (B+/B+)(Cd/Cd)(sy/sy) Gabino Azul barrado (B+/B+)(G/+) Extendido azul o negro (B+/B+)(C?/C?)(S/S) Extendido rojo (Ba/B+)(C?/C?)(S/+) Gabino Marrón Checker (b+/b+)(G/+) La combinación heterocigótica (G/+), presenta mas pigmento que blanco, es el gabino barrado. La combinación (G/G) homocigótica, en una barrada azul o marrón produce el típico blanco con bordes de las alas oscuras (ala de cigüeña) (sy) ahumado (smokey) Extendido marrón (b/b)(C?/C?)(S/+) Cruzando un macho negro con una hembra azul barrada, observamos la descendencia para ver si salen pichones con patrón barrado, checker, etc. que comparándolos a su vez con el patrón barrado de la madre y teniendo en cuenta la tabla de dominancia, podremos deducir cual es el patrón que porta el macho negro. Ahumado marrón barrado (b/b)(C+/C+)(sy/sy) Es un gen recesivo, autosomal, muy común en palomas pica y de raza que transforma el pigmento base a otro más apagado. Los ahumados llevan aparejadas una serie de características como son: - Pico claro, llegando en algunos casos al tono marfil. - Supresión de los bordes blancos de las plumas extremas de la cola. Ahumado rojo barrado (Ba/Ba)(C+/C+)(sy/sy) Ahumado rojo checker ahumado (Ba/Ba)(Cd/Cd)(sy/sy) CONTINUARÁ El COLOR DEL PLUMAJE EN COLOMBICULTURA II (J. Rubio. Sevilla, 2004) En el artículo “El color de las palomas” publicado en el número 3 de Colombicultura Andaluza esbozábamos los primeras nociones para comprender como se trasmite genéticamente del color del plumaje; aquí, como continuación de dicho artículo tratamos “otros modificadores no sexables ó autosomales” habituales en los palomos deportivos. (Adicionalmente publicamos el artículo “Algo sobre Genética en Colombicultura” que alumbrará pormenorizadamente a los neófitos en la materia). Los modificadores son pares genéticos que pueden cambiar la apariencia de una paloma afectando al pigmento, al patrón e incluso a otros modificadores. De por si existen nosexables o autosomales (con dos genes en machos y hembras) y ligados al sexo o sexables (con dos genes en los machos y un solo gen en las hembras). De entre la multitud de modificadores conocidos haremos una segunda exposición de los más frecuentes en nuestras palomas deportivas “de pica y de raza buchona”, sin olvidar que pueden mostrarse yuxtapuestos con otros y el resultado final del color de una paloma será la suma de (pigmento+patrón+x modificadores). He de aclarar que en los ejemplares mostrados, la codificación genética del color no se obtiene exclusivamente de la visualización de la fotografía, sino que a veces es consecuencia del conocimiento del propio animal y de la ascendencia y descendencia del mismo; “Lo que aparentemente parece un determinado color de plumaje, en realidad puede ser otro con uno o varios modificadores yuxtapuestos”. EL MODIFICADOR “BLANCO” El “blanco” es un tema bastante complejo por la simple razón que una paloma blanca puede ser el resultado de varias combinaciones genéticas. No es propiamente un color, sino la ausencia del pigmento de cualquiera de los colores, presentando al final un color blanco. Existen genes que inhiben totalmente la formación de melanina como es el caso del “blanco recesivo” o del “albino”. Existen otros que inhiben parcialmente la formación de melanina y que al ir combinándolos entre si pueden dar animales cada vez con menos pigmento hasta llegar al blanco, tal es el caso de la combinación del gabino homocigótico, del gabino y del rojo ambos homocigóticos, del gazzi, etc. Vamos a estudiar algunos de ellos: (zwt) Blanco recesivo (rec.white) Existe el gen “zwt” blanco recesivo, el cual inhibe la formación de melanina. Cuando este gen se combina en forma homocigótica (zwt/zwt) el animal resultante es completamente blanco. La mayor parte de los blancos recesivos “zwt” tienen ojos oscuros u "ojo de toro". Blanco recesivo (zwt/zwt) Para saber cual es el color de base que porta, habría que emparejar el ejemplar en cuestión con una azul wildtype sin el gen zwt, y en función del resultado obtenido, deduciremos que color hay debajo de ese blanco aparente. Si un par de blancos recesivos son emparejados, todos los pichones serán blancos. Si un blanco recesivo es emparejado con una paloma que no lleve el gen blanco recesivo, todos los pichones se pigmentan y por tanto el blanco se porta oculto. Si dos ejemplares pigmentados portadores del blanco recesivo son emparejados, la descendencia responderá a la relación mendeliana clásica de 1:2:1 25% blancos; 50% pigmentados llevando blanco; 25% pigmentados sin llevar blanco (z) Figura o pío (gazzi) Detalle del ojo oscuro o de toro El blanco recesivo funciona de forma similar al “rojo recesivo” pero aplicando el “blanco”; es opaco a todos los demás colores, patrones y modificadores, con una posible excepción, como es la del propio color rojo recesivo. (Como regla nemotécnica hemos de pensar en un animal pigmentado, al cual le hemos dado una capa de pintura blanca en su totalidad) Es un gen recesivo simple no-sexable, el cual inhibe la formación de melanina a base de manchas homogéneas semejantes a las que presentan los caballos píos. Una característica diferencial al margen del plumaje, es que presenta los ojos manchados parcial o totalmente. (Como regla nemotécnica hemos de pensar en un animal pigmentado al cual le hemos dado una capa de pintura blanca dejando huecos o zonas sin pintar a modo de parches). El gabino homocigótico (G/G) provoca en la paloma una marca o pigmentación similar a las cigüeñas (cuerpo blanco con bordes de las alas y cola oscuros). Azul con patrón barrado y Gazzi (B+b+)(C+/C+)(z/z) (ab) Albino (albino) El albinismo es una mutación que inhibe totalmente la formación de melanina, por lo que puede darse también sobre cualquier color y patrón. La forma mas fácil de distinguirlo es observando los ojos de la paloma, que son de color rosa. (Ba/*)(G/*) Blancos de gabino y rojo (white of grizzle and red) Hay formas de conseguir un blanco a través de la inhibición de formación de melanina de rojos y gabinos, que al combinarlos dan animales blancos. El rojo pigmenta la pluma desde la base hacia el exterior y el gabino lo hace a la inversa, por lo que al combinar ambos, dan como resultado una pluma blanca sin pigmentar. Gabino homocigótico (G/G) Otros blancos Son el resultado de uno o más genes de razas específicas, que provocan palomas figuras o de color pío (saddle, whiteside , lahore, etc.) y que en los palomos deportivos no suelen darse. (G/G) blanco de gabino homocigótico (white of grizzle) (f) Melado, florido o Puleva (flowery) El melado es un gen recesivo no sexable opaco, el cual al igual que el rojo recesivo enmascara tanto el pigmento como el patrón que porta la paloma, mostrando un plumaje mezclado de plumas blancas y rojizas oscuras, portando el pico claro. (Está por comprobar que dicho gen sea un alelo del rojo recesivo, circunstancia bastante probable). La gama de pigmento de este modificador va desde el mayoritariamente blanco con algunas manchas rojizas, hasta el mayoritariamente rojizo con plumas blancas, aunque a nivel genético, por ahora, no entremos en tal distinción. MÁS MODIFICADORES (e) Rojo recesivo (rec. red) El color rojo recesivo es un gen recesivo no sexable el cual enmascara el pigmento del color básico y el patrón, provocando una coloración roja oscura uniforme. Muchos rojos recesivos poseen una suciedad que embota el color y la cola es aún más marrón, esto es particularmente cierto en el rojo recesivo de palomas mensajeras. Detalle de plumas roja y gabina Un gabino-rojo homocigótico será casi blanco, con tal vez unas cuantas plumas rojas. Estos blancos se pueden distinguir normalmente de otras combinaciones genéticas porque tendrán ojos anaranjados en vez de los ojos “oscuros” o "de toro" de los blancos recesivos y pintos. amarillo recesivo es similar a marrón de las gallinas. Melado o florido blanco (f/f) Melado o florido rojizo (f/f) (Di) ó(V) Sucio (dirty) Rojo recesivo (e/e) Según muchos aficionados el modificador “índigo ó añil” mejora bastante perceptiblemente el color, y el factor S “extensión” hace lo mismo. El amarillo recesivo es la combinación del gen “rojo recesivo” con el gen “d” diluido, que es también recesivo. El Gen dominante no sexable que provoca un oscurecimiento tanto el pico como las patas y las plumas, que tienden a un tono “sucio” nunca mejor dicho. Esta mutación a veces se ve acompañada por lagrimales blancos. Sucio us Wildtype (B+/B+)(C+/C+)(V/+) Jaspeado azul barrado con brazalete (+/+)(C+/C+)(So/*) (In) índigo u oxido El índigo, añil u oxido es un gen no sexable “parcialmente dominante”, por lo que cualquier ejemplar que lo porte lo mostrará. Presenta dos estados: 1.- En su forma heterocigótica generan barrado color bronce ú oxido, con la grupa mas oscura de lo normal y el cuello bronceado. Sucio us Wildtype (B+/B+)(C+/C+)(V/+) (Ug) Fajado (undergrizzle) Gen dominante no sexable que provoca un aclarado desde la parte más próxima al cañón hacia la mitad de la pluma, que afecta principalmente a las remeras primarias y a las timoneras de cola, tendiendo dicha zona a un tono blanquecino o nevado. Indico Heterocigótico (+/+)(C+/C+)(In/+) Indico Heterocigótico (+/+)(Cd/C+)(In/+) a) En los "azules" cambia el color dándole un tono "terrizo oscuro" y las barras y el checker o moteado se muestran de color "oxido". Lava también el barrado de la cola, así en lugar de ser negra, es más clara que el resto de la cola. En una base extendida-negra produce una paloma con un color terrizo casi blanco y bordes oscuros. b) En combinación con el marrón el resultado que se asemeja al rojo ceniza, pero con vivos marcadores en las alas y algunos "chismes" sobre la cobertura de la cola. c) El índigo con el color rojo ceniza es virtualmente indistinguible del color rojo ceniza, aunque cuando se combina con el modificador extendido a menudo dé a un efecto de “caoba” vivo. La combinación del índigo tanto heterocigótico como homocigótico con el extendido es el conocido como “añil andaluz” ALIBLANCOS Y TREPADOS (whitewing) Fajado azul ahumado plumeta (+/+)(C+/C+)(sy/sy)(L/+)(Ug/*) (So) Jaspeado (sooty) Gen dominante no sexable que provoca un oscurecimiento de la zona central de las plumas cobertoras dando la apariencia de escamas. Índigo prieto (+/+)(Cd/C+)(In/+) 2.- En su estado homocigótico: se puede confundir con un ceniza rojo, aunque presenta grupa y cuello más oscuros El aliblanco es un plumaje que se da habitualmente en los palomos “pica”, consistente en mostrar una o varias plumas blancas en las remeras primarias de una o ambas alas. Partiendo de la base de que podemos tener pichones aliblancos acoplando un macho y una hembra donde ninguno de los dos muestra el aliblanco, deducimos que el “aliblanco” es un gen recesivo. (l/l). De la misma forma, acoplando un macho y una hembra, ambos aliblancos visibles, podemos obtener una descendencia NO aliblanca, por Aliblanco recesivo azul barrado (+/+)(C+/C+)(l/l) Aliblanco dominante, azul, checker y fajado (+/+)(Cd/?)(L/l)(Ug/+) TABLA DE COLOR EJEMPLOS En la siguiente tabla expresamos la codificación de algunos de los colores más habituales en los palomos deportivos, sin olvidar que los pares genéticos sexables en las hembras cuentan con un único gen. Ejemplo 1. Pigmentos Macho B+/b = azul portando marrón Hembra Ba/. = roja ceniza 1 2 3 4 B+ / b Ba / . = (B+/Ba) (B+/.)(b/Ba)(b/.) Modif.3 Modif.2 Modif.1 Topónimo Patron TABLA DE COLOR PigmentoI+Patron+Modificadores. (JRubio) Color tanto, deducimos que el aliblanco es un gen dominante. (L/?) De la contradicción de los exponendos anteriores queda probado que existen dos alelos o tipos, uno, el aliblanco dominante (L) y otro, el aliblanco recesivo (l). Ambos genes (L) y (l) son alélicos, pudiendo llegar a presentarse en una forma mixta (L/l) “aliblanco dominante y recesivo”, aunque lo que apreciaríamos sería igual que el aliblanco recesivo (l/l); en otras palabras, cuando el aliblanco recesivo está presente en forma homocigótica (l/l) adquiere el efecto del patrón dominante, sin tener efecto alguno la dominancia. COLORES CON PIGMENTOS Y PATRONES BASE Castaño bb C? Azul +? +? Azul Gotado + ? CL ? Mosqueado o rodado +? C? Prieto o pizarro + ? CD ? Magaño + ? CT ? Bucaro o bayo +? BA * Rojo C? BA * COLORES CON MODIFICADORES AUTOSOMALES Marrón bb S* Negro +? S* Negro Azabache Brillo +? S* G* Plateado o ceniza S* BA * Gabino barrado ** G? Gabito Tigre ** GT ? Gabino Homocigotico o ** GG Cigüeña Ahumado +? + ? sy sy Acerado +? C ? sy sy Moracho + ? CT ? sy sy Blanco ** zwt zwt Figura o Pinto ** zz Albino ** ab ab Rojo recesivo ** ee Melado, Florido o ** ff puleva Sucio ** V* Fajado ** Ug * Jaspeado ** So * Indigo u Oxido ** In * Aliblanco (recesivo) ** ll Aliblanco (dominante) ** L* COLORES CON MODIFICADORES SEXABLES Borrado o reducido ** rr Decolorado o lavado Stf ? Salpicado St * Canela bb dd Plata +? dd Negro Ala de Mosca +? +? dd Rosado C? dd BA * OTROS MODIFICADORES Inverso u opal dominante Inverso recesivo Cabeza negra Mosaico y Otros S* Od * oo pc pc Nomenclatura gen del wildtype (azul barrado) cualquier gen gen recesivo respeto a su par + * ? Serie marrón Serie Azul Serie roja Cualquier Serie sy sy Si apareamos un macho heterocigótico azul llevando también marrón, este portaría en un gen el azul “B+” y en el otro el gen marrón “b”, de aquí (B+/b); como hembra tomamos una paloma roja, por lo que su cromosoma del sexo portaría el gen “Ba” y el neutro o inexistente “.” ; (Ba/.) En este ejemplo se han considerado todas las posibilidades de mezcla de los pares genéticos y obtenemos que todos los machos serán rojos, unos portadores del pigmento azul como factor recesivo (1) y otros portadores del pigmento marrón también recesivo (3) y todas las hembras serán azules (2) o marrones (4). De la combinación anterior hacemos básicamente dos interpretaciones: Aquellos que en su par genético lleven “.” son hembras. El color dominante es el que se muestra fenotípicamente y se expresa en Mayúsculas, mientras que el recesivo u oculto se expresa en minúscula. Ejemplo 2. Patrones Macho Ct/C+ = azul terciopelo (magaño) Hembra Cl/C+ = azul gotada 1 2 3 4 Ct / C+ Cl / C+ = (Ct/Cl) (Ct/C+) (C+/Cl) (C+/C+) Para el siguiente ejemplo elegimos una pareja de palomas con pigmento azul homocigótico, (de la misma forma se puede hacer para cualquier otro pigmento); el macho “magaño” con patrón terciopelo-barrado (Ct/C+) y la hembra “azul gotada” con patrón gotado-barrado (Cl/C+). Del acople en cuanto a patrones se refiere obtenemos: 1 será terciopelo llevando gotado, 2 será patrón-T llevando barrado, 3 será gotado llevando barrado y 4 será barrado homocigótico. Es importante recordar que fenotípicamente el orden de dominancia de los patrones va en orden descendiente desde los más oscuros a los más claros, por eso, en patrón-T ó terciopelo es dominante sobre el gotado y este a su vez sobre el barrado. De aquí se deduce claramente que de una pareja aparentemente no aliblancos podamos obtener descendencia aliblanca fenotípicamente. Ejemplo 3. Ahumado (recesivo) Ejemplo 6. Aliblanco (dominante y recesivo) macho hembra 1 2 3 4 sy / + sy / sy = (sy/sy) (sy/sy) (+/sy) (+/sy) Formamos una pareja con el modificador Ahumado; para ello tomamos un macho azul portando ahumado heterocigótico, (aunque fenotípicamente no muestra el ahumado por ser recesivo) y una hembra ahumada homocigótica. Aquí, 1 y 2 son ahumados homocigóticos y así lo mostrarán en su fenotipo, mientras que 3 y 4 son ahumados heterocigóticos y aunque porten dicho gen no lo mostrarán en su fenotipo. Ejemplo 4. Gabino (dominante) macho hembra 1 G/ + + / + = (G/+) 2 (G/+) macho hembra 1 L/ + l / + = (L/l) 2 (L/+) 3 (+/l) 4 (+/+) Si formamos una pareja, con un macho de cualquier pigmento, aliblanco dominante heterocigótico, por tanto visible su aliblanco, y una hembra aliblanca recesiva heterocigótica, (no visible su aliblanco), la descendencia será: (1) Aliblanco dominante y recesivo, y por tanto, visible. (2) Aliblanco Dominante (3) Aliblanco recesito heterocigótico (no visible) (4) No aliblanco. Ejemplo 7. Descripción y 3 (+/+) 4 (+/+) nomenclatura Si formamos una pareja, con un macho gabino heterocigótico que fenotípicamente muestra el gabino por ser este un gen dominante y una hembra no gabina. La descendencia será: gabino heterocigótico de una parte (1 y 2) y no gabinos por otra (3 y 4). Ejemplo 5. Aliblanco (recesivo) macho hembra 1 2 3 4 l/ + l / + = (l/l) (l/+) (+/l) (+/+) Si formamos una pareja, con un macho azul barrado y una hembra prieta, ambos fenotípicamente no muestran el aliblanco, aunque si lo portan. La descendencia será: (1) Aliblanco recesivo homocigótico, y por tanto, visible. (2) y (3) aliblanco recesivo heterocigótico, no visible. (4) No aliblancos Veamos como se establecería la nomenclatura del color de algunos ejemplares (indudablemente su cadena genética de color se ha obtenido cruzando el ejemplar en cuestión con hembras de distinto pigmento-patrón-modificadores y observando la descendencia, incluso cruzando hasta con varios niveles de reencaste). 1.- La descripción del palomo “gabino negro” de la foto sería: “azul homocigótico, con patrón barradochecker oscuro, extendido, gabino, ahumado y aliblanco dominante plumeta”, utilizando la notación del pie de foto. Gabino Negro (B+/B+ )(C+/Cd)(S/+)(G/+)(sy/+)(L/+) 2.- La descripción del palomo “figura” de la foto sería: “azul homocigótico, con patrón terciopelo-barrado, figura, aliblanco dominante-recesivo, sucio e índigo,” utilizando la notación del pié de foto El COLOR DEL PLUMAJE EN COLOMBICULTURA III En los artículos publicados en los números 3 y 4 de Colombicultura Andaluza esbozábamos las nociones para comprender como se trasmite genéticamente del color del plumaje, para ello, explicábamos como los modificadores son pares genéticos que pueden cambiar la apariencia de una paloma afectando al pigmento, al patrón, e incluso, a otros modificadores. Los modificadores los podemos dividir en dos grupos: sexables y nosexables o autosomales; en los primeros, la hembra tiene dos genes mientras que en los segundos solo dispone de un gen asociado al modificador en cuestión. En los modificadores existe a su vez un orden de dominancia o prevalencia permitiendo que unos modificadores puedan ocultar a otros total o parcialmente, de ahí que a veces se escuche la expresión "el color rojo recesivo es opaco”; esto significa simplemente que color rojo recesivo cubre u oculta todos los pigmentos, patrones y modificadores. Existen diversidad de modificadores que aunque identificables fenotípicamente, no están identificados genéticamente ni agrupados con los alelos correspondientes. Aquí, como continuación de los artículos anteriores tratamos los “modificadores sexables” más habituales en los palomos deportivos, así como, otros “no sexables” de cierto interés, bien por su singularidad o bien porque han sido descubiertos recientemente. Hemos de aclarar que en los ejemplares mostrados, la codificación genética del color no se obtiene exclusivamente de la visualización de la fotografía, sino que a veces es consecuencia del conocimiento del propio animal y de la ascendencia y descendencia del mismo; “Lo que aparentemente parece un determinado color de plumaje, en realidad puede ser otro con uno o varios modificadores yuxtapuestos”. MODIFICADORES SEXABLES Un emparejamiento sexable es el acople de una pareja de palomas cuyas características de color están “asociada al sexo”; para que esto ocurra el gen de la hembra ha de ser dominante respecto al gen del macho para esa característica en cuestión. Los modificadores “diluido”, “reducido”, “salpicado” y “decolorado”, combinados con los tres pigmentos azul, marrón y rojo-ceniza son sexables. Cuando se realiza un emparejamiento sexable todos los pichones que tienen el color o característica asociada a la madre serán machos y todos los pichones que tengan el color o característica del padre serán hembras, en otros términos, en un apareamiento sexable los pichones son del color y sexo opuesto al de los padres (descendencia cruzada). La razón deviene de que el macho tiene dos cromosomas sexuales activos y la hembra sólo uno, mientras que el otro es nulo; al combinarlos entre si, ocurre que el gen que se combina con el “nulo” resulta ser hembra. Los ejemplos de apareamientos asociados-sexuales llamémosle “cruzados” (en que los jóvenes tienen los colores opuestos de sus padres) entre otros incluyen a: Macho marrón y hembra azul o roja = hijos azules o rojos; hijas marrones. Macho azul y hembra roja = hijos rojos; hijas azules. Macho diluido y hembra no diluida o intensa = hijos no diluidos; hijas diluidas. Macho reducido y hembra no reducida = hijos no reducidos; hijas reducidas. Macho normal y hembra lavada = hijos lavados, hijas normales. [d+] Intenso, [dP] Pálido, [d] diluido y [dex] super diluido El factor de dilución es un gen sexable recesivo que aclara progresivamente el tono del pigmento sin alterar el patrón. Existen 4 alelos conocidos: intenso, pálido, diluido y super diluido en orden decreciente de tonalidad. Son alélicos, es decir, ocupan el mismo lugar geométrico en el cromosoma Z del sexo, por lo que se da uno u otro, no se pueden presentar conjuntamente De Izda. a Dcha.: normal ó intenso, diluido, súper-diluido – (D+) “Intenso” Es el tono normal presentado por la wildtype. – (dP) “pálido” El efecto del gen pálido es aligerar el color total de la paloma, con un efecto algo más ligero que el color intenso pero sin llegar a la claridad del diluido. Aligera los colores azul y marrón anaranjando los rojos – (d) “diluido” Aligera el color total de la paloma más que el pálido sin llegar al super diluido. Al nacer no presenta pelusa. Tiene distintas denominaciones según al pigmento al que se le aplique: .- Plata = azul diluido .- Caqui o castaño = marrón diluido. Presenta la cola barrada y es susceptible a la decoloración por el efecto continuado de la luz solar. .- Crema o amarillo = rojo diluido, (presenta la cola sin barra) Azul terciopelo prieto diluido (B+B/+)(Ct/C+)(d/d) Azul barrado extendido diluido (B+/B+)(C+/C+)(S/+)(d/d) Azul barrado reducido (B+/B+) (C+/C+) (r/r) Salpicado (B+/B+)(C+/C+)(S/+)(G/+) (St/+) – (StF) Qualmond, Lavado ó Decolorado Es similar al Almond con el pigmento y las manchas más descoloradas y pequeñas pero cambiantes con la edad. Rojo checker diluido (Ba/+)(C/C)(d/d) Magaño ó Azul Terciopelo reducido (B+/B+) (Ct/C+) (r/r) En los rojos es el conocido rosado o puleva Rojo checker gabino diluido (Ba/+)(C/C+)(G/+)(d/d) [r] Reducido Es un gen recesivo sexable que reduce la cantidad de pigmento aplicado a la pluma, por lo que, tiene proporcionalmente menos melanina en todo el cuerpo, transformando los pigmentos a tonalidades rojizas apasteladas. Sus alelos son: – (r+) “no reducido” o “normal” – (r) reducido – (r RU ) sarampión reducido Aunque el gen reducido aligera los colores no pertenece al modificador de dilución, por lo que puede aparecer conjuntamente con los genes diluidos. En los azules provoca un color base iluminado-escarchado, a menudo con un plateado creciente con brillos verdosos en el área del buche, con barras y moteado tirando a rosado (en los barless por supuesto, no). Los Lavados Existe una familia de modificadores que consiste en un color base homogéneo, mas o menos decolorado e iluminado, sobre el que en ocasiones, y dependiendo de con que otros modificadores se acople produce un salpicado de plumas de otras tonalidades. Dicho gen, que es sexable, simplemente es un gen "iluminador" que suprime los efectos de la mayor parte de los pigmentos. Aunque existe una diversidad de variantes o alelos (St; StF; StFr; StH; Stsa; StC;) solo veremos a titulo indicativo algunos de ellos. – (St) Almond o Salpicado. Mutación de las más antiguas conocidas en la cual la paloma presenta manchas o salpicaduras de múltiple colores (rojo azul, marrones, …) y zonas decoloradas que con cada año van cogiendo más pigmentación hasta casi ponerse de su color normal. Es un gen dominante. En una paloma azul provoca un blanco sucio con unos cuantos puntos de color oscuro. Detalle del Decolorado ó Qualmond (B+/B+)(C+/C+)(Stf/+) Decolorado o Lavado (B+/B+)(C+/C+) (StF/+) – (StFr) Frosty, Nevado ó escarchado. Aparentemente es un gen recesivo ligado al sexo que solo se muestra en los machos. Produce un efecto más iluminador o nevado que el qualmond , distinguiendose sobre todo por el contraste de luminosidad entre la cabeza y el cuerpo con el cuello. b) Cuando se combina con el negro los resultados van de un color base plata, a un gris metálico revólver. c) Con el rojo ceniza es a menudo indistinguible de color rojo ceniza ordinario. (o) Opal o Inverso recesivo Escarchado o Nevado StFr (B+/B+)(C+/C+)(Stfr/+) OTROS MODIFICADORES NO SEXABLES Existen dos genes independientes no sexables parecidos fenotípicamete al “reducido” son el Ópalo dominante y el Ópalo recesivo Es frecuente para el aficionado inexperto confundir ópalo recesivo con ópalo dominante que tienden a aclarar o invertir el barrado y el moteado del patrón. (Od) Opal o Inverso dominante El ópalo dominante es un gen dominante autosomal. Es extremadamente variable en su expresión, yendo desde un ligero lavado en las barras y el moteado o checker, hasta barras y checker completamente blancas. Al mismo tiempo la barra de la cola puede ser descolorida o puede ser casi negra. Una vía para identificar al ópalo dominante es el coloreado claro en el tallo o eje de las plumas de la cola. El ópalo recesivo es otro gen recesivo autosomal que transforma las barras y alares y de la cola, convirtiéndolos en más iluminadas y vivas. Ese efecto iluminador es extremadamente variable, y las marcas de las alas pueden ir de un gris medio hasta un tostado grisáceo, a un negro "bronce" ó a un color rosado o rojizo. Este último a menudo imita el color de color rojo ceniza, incluyendo la ausencia de la barra de la cola. Cuando esto sucede, ciertos aficionados piensan que han conseguido rojo ceniza a partir de dos azules, cosa que es genéticamente imposible. Si tal color pseudo-rojo o pseudo-plata es emparejado a un azul que no lleva el gen ópalo recesivo, todo pichones serán azules. En otras palabras cuando el ópalo recesivo está presente en forma homocigótica (o/o) entonces adquiere el efecto del patrón dominante, sin tener efecto alguno la dominancia El opal recesivo extendido presenta un color metálico. Tengamos presente el extendido siempre enmascarará cualesquier patrón. a) En el color base azul, otro efecto común es que se ilumina hasta un gris acerado. El barrado o checker blanco es el efecto más probable para saber si el gen está en combinación con de color azul indigo, aunque esa combinación no siempre produce los mismos resultados. Color Plata correspondiente al Rojo Extendido (Ba/B+)(C+/C+)(S/+) SIN BARRA O BARLESS Azul Checker Ópalo Recesivo (B+/B+)(C/C+)(o/o) Azul Barrado Ópalo Dominante (B+/B+)(C+/C+)(Od/+) gen sexable recesivo diluido homocigótico. 2.- Para mensajeristas el término “plata” también denominado por ellos como “silver dun bar” o "rojo plata", es simplemente un color rojo ceniza barless, a menudo con un lustre escarchado o brillo plata en las plumas del cuello. 3.- Para los criadores de Deportivos (pica) viene a ser un color gris terrizo homogéneo que tiene distintas afecciones según la zona donde nos encontremos: pelorata, ceniza, plateado, etc. y que estadísticamente responde al rojo extendido. El COLOR PLATA El “plata” es un tema bastante complejo por la simple razón que una paloma de color plata puede ser el resultado de múltiples combinaciones genéticas. A su vez el término plata tiene significados distintos: 1.- Para genetistas y criadores de especies de fantasía, significa lo mismo que un “azul diluido sin barrar “, esto es, un wildtype barless con el Es la característica por la cual desaparece la “barra” de la cobertura de las alas, mostrándose el color base sin patrón alguno. Es un gen autosomal (no está en el cromosoma sexual) y recesivo (debe poseer dos genes, para que dicha característica sea visible). .- En los rojos se manifiesta como “rojo barless” aunque a veces se confunde con el “rojo extendido”. Supongamos que tenemos un ejemplar “rojo plata” que no sabemos si es “rojo barless” o “rojo extendido”. Se puede comprobar cruzando dicho ejemplar con otro de color “azul” y patrón “barrado” ó “prieto” y observando el resultado: – Si obtiene algún “negro” es que porque el patrón es “extendido” en vez de “barless” – Si se obtiene descendencia que se parece al progenitor estudiado (es decir del que no sabíamos si era barless o extendido) estamos seguros también que no es “barless”, ya que barless es recesivo. Azul sin barra o barless (B+/B+)(c/c) ALELOS MULTIPLES En 1981, en un artículo del diario americano de “the pigeon” se escribió acerca de un gen mutante recesivo nuevo, responsable del color que paso a llamarse "penciled" -pc- (cabeza negra). Por otra parte, se pensaba que el gen “tail mark” (todo-blanco excepto la cola) era el efecto de un gen independiente. En 1993 Dr. Willard F. Hollander estudió la dominación parcial del gen –pc-, estableciendo una teoría en base a la cual, se deduce que existen una serie de genes alélicos al “figura” o “gazzi” –z- (fajado, figura, cabeza negra, cola negra, blanco recesivo, …) que al combinarlos entre sí dan como resultando colores intermedios a los de partida. Asumiendo que la hipótesis de Hollander tenga razón, tenemos al menos 6 alelos conocidos en la serie del gen “z”. El resultado de la combinación de los distintos alelos es el mostrado en la siguiente tabla: z+ wildtype z fl fajado z figura z pc cab. negra z tm cola negra z wh blanco recesivo z+ wildtype zurita (z+ / z+) wildtype (z+ / z fl) wildtype (z+ / z) wildtype (z+ / z pc) wildtype (z+ / z tm) wildtype (z+ / z wh) wildtype z fl fajado undergrizzle (z fl / z+) wildtype (z fl / z fl) fajado (z fl / z) intermedio ? (z fl / z pc) destello (flash) (z fl / z tm) destello-cola negra ? (z fl / z wh) salpicado (splash) Este artículo no hubiera sido posible sin la colaboración directa en unos casos, y la consulta de las obras publicadas en otros de: W. Hollander, R. Huntley, F. Mosca, B. Mangile, T. Barnath, y J. Espinosa. Juan Rubio Ortiz. Sevilla, 2004. z figura gazzi (z / z+) wildtype (z / z fl) intermedio (z / z) figura (z / z pc) intermedio (z / z tm) intermedio (z / z wh) intermedio z pc cab. negra () penciled (z pc / z+) wildtype (z pc / z fl) destello (flash) (z pc / z) intermedio (z pc / z pc) cab. negra () (z pc / z tm) intermedio (z pc / z wh) intermedio z tm cola negra tail mark (z tm / z+) wildtype (z tm / z fl) destello-cola negra ? (z tm / z) intermedio (z tm / z pc) intermedio ? (z tm / z tm) cola negra (z tm / z wh) intermedio z wh blanco recesivo recesive white (z wh / z+) wildtype (z wh / z fl) salpicado (splash) (z wh / z) intermedio (z wh / z pc) intermedio (z wh / z tm) intermedio ? (z wh / z wh) blanco recesivo ALGO SOBRE GENETICA EN COLOMBICULTURA (J.Rubio. Sevilla, 2004) En las palomas existen diferentes cualidades que reunidas en solo ejemplar le confieren el grado de calidad necesario para triunfar en una competición deportiva. Parte de dichas cualidades son heredadas de sus progenitores (unas del padre, otras de la madre, otras mezcladas) y otras son producto del aprendizaje. Si se quieren criar palomas con cierto éxito, es imprescindible conocer algunos de los principios esenciales sobre las leyes de la herencia. Con estas reglas se puede reconocer ANTES de la cría, algunos factores que tienen un papel importante en la descendencia, y que a la postre, conseguirán influir en la calidad del ejemplar buscado. Tras consultar múltiple bibliografía, y en vistas a lo árido que resulta cualquier tratado sobre genética, vamos a intentar hacer una síntesis “coloquial” que nos permita al menos, poder entender algo sobre la herencia genética, y que más tarde podemos ir corroborando con la aplicación de dicha técnica al “color de las palomas”. Algunos de los términos aquí utilizados derivan directamente del inglés y son comúnmente aceptados en la comunidad científica, por lo que se ha optado por mantenerlos, no obstante se indica cual sería la traducción. CROMOSOMAS Y GENES El cuerpo de cualquier animal se compone de células; en el núcleo de estas células están los cromosomas; en estos están encadenados los genes, que a su vez contienen las moléculas de ADN. Numéricamente podemos expresarlo de la siguiente forma: El Cuerpo de una paloma (10.000 millones células)1 célula (40 pares de cromosomas)1 cromosoma (100.000 genes)1 gen (millones ADN) La cantidad de cromosomas existente en el núcleo de cada célula, es especifica de cada especie, así por ejemplo en el hombre es de 46, el perro 78 y en la paloma de 80. Un cromosoma podemos imaginarlo como una cadena de genes (puede abarcar desde centenares a miles de genes encadenados). Un gen, podemos imaginarlo como un eslabón en dicha cadena. -O-O-O-O-O-O-O-O-O-OLa forma en que se sitúan los cromosomas es por pares, (uno proveniente del padre y otro de la madre) formando una doble hélice, donde de forma general, están enlazados cada uno de los genes de un cromosoma con su equivalente en el cromosoma ligado. Una célula puede regenerarse o duplicarse por sí misma con una copia idéntica (Mitosis) o a través de una combinación con otra célula durante la reproducción sexual (Meiosis) para la creación de un nuevo ser. Los pares de cromosomas se dividen durante la maduración de las células sexuales, en lo que se conoce como división meiótica, por lo que cada célula sexual (esto es el ovulo no fecundado de la hembra y el esperma del macho) tienen solo la mitad del conjunto de cromosomas; Tras la fecundación se unen recombinándose los cromosomas en el nuevo ser, formando de nuevo 40 pares, es aquí donde se crea la oportunidad para la inclusión de genes alternativos o “alelos”, por tanto podemos resumir que las palomas tienen 40 pares de cromosomas en cada célula, los cuales se reciben por mitades de los progenitores, el padre suministra 40 cromosomas y la madre otros 40, emparejándose uno a uno, dando como resultado 80 cromosomas agrupados en 40 pares. Con excepciones, los cromosomas van en pares, por lo tanto cada gen en un cromosoma tiene una copia emparejada en la misma localización de su cromosoma vecino, y está unido a este. Al sitio de la localización de cada gen en un cromosoma se llama “lugar geométrico”, que solo puede ser ocupado por un gen a la vez; es como si fuera un “habitáculo” en el cual se sitúa el gen, por lo tanto, para una “característica genética”, hay dos habitáculos posibles, la del propio gen y la del gen del cromosoma ligado, dentro de las cuales se pueden ubicar el propio gen o alguno de sus alelos o alternativos. A cada uno de estas parejas de genes que son los responsables de la manifestación de una determinada característica en el animal se les conoce como “par genético” (B/b). DOMINANCIA Y RECESIVIDAD Originado por el padre y la madre, el par genético resultante tras la fecundación, puede tener efectos diferentes. Si priman las propiedades de un gen respecto a otro, este se denomina Dominante, por tanto las propiedades del otro gen quedan ocultas y se denomina Recesivo. Para poder operar analíticamente con los genes hemos de compararlos siempre con uno de referencia, por lo que, como animal estándar tomaremos la “Paloma Zurita Salvaje”, “Wildtype” o “Columba Libia” y sus características asociadas a genes las simbolizamos con “+” Si la acción de un solo gen al combinarlo con el wildtype “+” predomina o se muestra visible se considera dominante, y si no es perceptible se considera recesivo. Wildtype es por tanto neutro o estándar. Ej. El tamaño de una paloma es una herencia intermedia Un gen o puede ser dominante sobre otro y a la vez recesivo sobre un tercero, la forma adecuada para poder operar analíticamente es compararlo siempre con el “wildtype” y referirnos a este como base. El carácter Dominante es siempre visible y oculta al recesivo que puede permanecer latente durante generaciones y manifestarse cuando se den las condiciones de combinación adecuadas. Algunas de las características con respecto a las cuales comparar la dominancia o recesividad de la Wildtype son: Los términos homocigótico y heterocigótico son palabras compuestas en la que “zigote” representa una célula que se forma de la unión de dos gametos (sperma+ovulo), hetero significa “distinto” y homo significa “igual”, por tanto, homocigótico implica un par genético con dos genes idénticos, mientras que heterocigótico implica un par genético con genes distintos. Si una paloma ha heredado de sus padres un par genético de las mismas propiedades para una determinada característica, se dice que tiene una dotación “homocigótica” (BABA ó bb). Si se combinan entre sí genes de diferente eficacia, la herencia es “heterocigótica” (BAb; BA+; b+…). Excepcionalmente, en determinados pares genéticos como el del sexo en las palomas hembras, uno de los cromosomas está atrofiado y, por lo tanto, se considera neutro o nulo, por lo que, las cualidades que se ubican en dicho cromosoma se consideran homocigóticas monogeneticas, es decir son homocigóticas a pesar de tener exclusivamente un gen (ya que el otro es neutro). Tal es el caso de los genes que controlan el pigmento, el factor diluido y el factor reducido en las hembras. Si una paloma homocigótica para un patrón dado es emparejada con otra también homocigótica pero con un patrón recesivo respecto al primero, todos los jóvenes tendrán el mismo patrón del padre dominante, pero llevarán el gen Wildtype + + + + + + + + plumaje gris pico negro borde del ojo blanco ojos naranjas patas rojas barras negras en la cobertura de las alas bode cola negra plumas extremas cola con borde blanco + grupa blanca Hemos de tener en cuenta que las características físicas (fenotipo) no se heredan, sino que los que se hereda son los genes (genotipo) que combinados de una determinada forma producen ese fenotipo. Ej. Dominante: Color Rojo, Pico Negro Ej. Recesivo: Color Azul, Pico Blanco Si ninguno de los dos genes es dominante o en la formación de una propiedad entran varios genes (poli-genética o multifactorial), en el proceso de la herencia se dan propiedades Intermedias En la notación utilizada todos los símbolos dominantes con respecto al wildtype se representan empezando por mayúscula y los recesivos por minúscula. HOMOCIGOSIS Y HETEROCIGOSIS para el modelo del padre recesivo, tales pichones son los antes mencionados heterocigóticos para cada patrón, es decir llevan intrínsecamente un gen que no es visible y que puede aflorar posteriormente. (A/A)+(a/a)= (A/a);(A/a);(A/a);(A/a) Si dos heterocigóticos son emparejados, en conjunto conseguimos el modelo mendeliano de 1:2:1 clásico, que significa 25% de esos jóvenes serán los homocigóticos para el gen dominante, 50% heterocigóticos, y homocigóticos 25% para el gen recesivo. En apariencia serán 3:1. (A/a)+(A/a)= (A/A);(A/a);(a/A);(a/a) Las condiciones recesivas son siempre homocigóticas en caso que se muestre visibles, es decir, se manifiesta cuando se ha heredado esta cualidad de ambos progenitores. EL CROMOSOMA DE SEXO En las “palomas hembra” todos los cromosomas tienen su par, excepto uno, el que determina el sexo, por ello, contrariamente a los mamíferos, la hembra es la que posee el cromosoma independiente "Z" que a la postre es el que determina el sexo. Algunas características de nuestras palomas, como el pigmento del color ( marrón=b, azul o negro=+ y Rojo=BA ) están incluidas en un lugar geométrico determinado del cromosoma del sexo. A efectos de explicación, en las palomas Hembras denominamos el cromosoma que debiera estar asociado al cromosoma Z, con el número cero “0” ó “.” pues, este cromosoma es inexistente y por tanto, vacío de color; es por ello que las hembras son homocigóticas en el color, es decir puras, sin mezcla, mientras los machos pueden ser dependiendo del cruce de colores homocigótico o portar dos colores, heterocigótico. Existen al menos nueve genes que se han identificado en 5 localizaciones del cromosoma del sexo de la paloma domestica Propiedades ligadas al sexo b brown marron A B ash-red ceniza rojo d dP dilution dilute-pale diluido claro; palido r reduced reducido St almond StF fadel StQ qualmond almendra lavado almendra de quinn wl mortal web lethal La posición o lugar geométrico de esos genes en el cromosoma del sexo, podríamos representarlo: Posiciones en el cromosoma sexual w eb brow n, dilute reduced lethal almond ash-red d r wl St LEYES DE MENDEL b En 1866, un padre agustino aficionado a la botánica llamado Gregorio Mendel publicó los resultados de unas investigaciones que había realizado pacientemente en el jardín de su convento durante más de diez años. Éstas consistían en cruzar distintas variedades de guisantes y comprobar cómo se transmitían algunas de sus características a la generación siguiente. Su sistema de experimentación tuvo éxito debido a su gran sencillez, ya que se dedicó a cruzar plantas que sólo diferían en una característica externa que, además, era fácilmente detectable. Por" ejemplo, cruzó plantas de semillas verdes con plantas de semillas amarillas, plantas con tallo largo con otras de tallo corto, etc. Mendel intuyó que existía un factor en el organismo que determinaba cada una de estas características. según él, este factor debía estar formado por dos elementos, una que se heredaba del organismo masculino y el otro del elemento. Además estos dos elementos consistirían en versiones iguales o diferentes del mismo carácter; cada valor del factor proporcionaría, por ejemplo, un color distinto a la semilla o una longitud de tallo diferente en la planta. Además, tal y como veremos más adelante, algunas, versiones serían dominantes respecto a otras. Actualmente a estos factores se les denomina genes, palabra derivada de un término griego que significa «generar», y a cada versión diferente del gen se la denomina alelo. Así el gen que determina, por" ejemplo, el color de la semilla en la planta del guisante puede tener " dos alelos, uno para las semillas verdes y otro para las semillas amarillas. Observando los resultados de cruzamientos sistemáticos, Mendel elaboró una teoría general sobre la herencia, conocida como leyes de Mendel. Primera ley de Mendel - Ley de la Uniformidad de los Mestizos. Si se unen padres de pura raza (homocigóticos) para una determinada propiedad, todos los hijos heredarán dicha propiedad por igual. AA+bb = Ab+Ab+Ab+Ab Si se cruzan dos razas puras para un determinado carácter, los descendientes de la primera generación son todos iguales entre sí y, a su vez, iguales a uno de sus progenitores, que es el poseedor del alelo dominante. Mendel elaboró este principio al observar que si cruzaba dos razas puras de plantas del guisante, una de semillas amarillas y otra de semillas verdes, la descendencia que obtenía, a la que él denominaba F1, consistía únicamente en plantas que producían semillas de color amarillo. Estas plantas debían tener, en el gen que determina el color de la semilla, los dos alelos que habían heredado de sus progenitores, un alelo para el color verde y otro para el color amarillo; pero, por alguna razón, sólo se manifestaba este último, por lo que se lo denominó alelo dominante, mientras que al primero se le llamó alelo recesivo. Segunda ley de Mendel - Ley de la División. Si unimos la primera descendencia de una pareja de raza pura, ya con propiedades de raza mixta (heterocigóticos) para una determinada propiedad, su descendencia se divide en la proporción 1:2:1. Ab+Ab=AA+Ab+bA+bb Los alelos recesivos que, al cruzar dos razas puras, no se manifiestan en la primera generación (denominada F1), reaparecen en la segunda generación (denominada F2) resultante de cruzar los individuos de la primera. Además la proporción en la que aparecen es de 1 a 3 respecto a los alelos dominantes. Mendel cruzó entre sí los guisantes de semillas amarillas obtenidos en la primera generación del experimento anterior. Cuando clasificó la descendencia resultante, observó que aproximadamente tres cuartas partes tenían semillas de color amarillo y la cuarta parte restante tenía las semillas de color verde. Es decir, que el carácter « semilla de color verde », que no había aparecido en ninguna planta de la primera generación, sí que aparecía en la segunda aunque en menor proporción que el carácter « semilla de color amarillo » Tercera ley de mendel - Ley de la Independencia. Todos los caracteres hereditarios de un ser vivo son heredados independientemente entre sí. Los caracteres que se heredan son independientes entre si y se combinan al azar al pasar a la descendencia, manifestándose en la segunda generación filial o F2. En este caso, Mendel selecciono para el cruzamiento plantas que diferían en dos características, por ejemplo, el color de los guisantes (verdes o amarillos) y su superficie (lisa o arrugada). Observo que la primera generación estaba compuesta únicamente por plantas con guisantes amarillos y lisos, cumpliéndose la primera ley. En la segunda generación, sin embargo, aparecían todas las posibles combinaciones de caracteres, aunque en las proporciones siguientes: 1/16 parte de guisantes verdes y rugosos, 3/16 de verdes y lisos, 3/16 de amarillos y rugosos y por ultimo 9/16 de amarillos y lisos. Esto le indujo a pensar que los genes eran estructuras independientes unas de otras y, por lo tanto, que únicamente dependía del azar la combinación de los mismos que pudiese aparecer en la descendencia. En la práctica, esto significa que mediante la cría se puede reunir en un mismo animal, distintas propiedades de los animales de partida. LA GENÉTICA DESPUÉS DE MENDEL: TEORÍA CROMOSOMICA DE LA HERENCIA A principios de este siglo, cuando las técnicas para el estudio de la célula ya estaban suficientemente desarrolladas, se pudo determinar que los genes estaban formados por acido desoxirribonucleico (ADN) y además se encontraban dentro de unas estructuras que aparecían en el citoplasma justo antes de cada proceso de división celular. A estas estructuras se las denominó cromosomas, termino que significa «cuerpos coloreados», por la intensidad con la que fijaban determinados colorantes al ser teñidos para poder observarlos al microscopio. Además se vio que estos aparecían repetidos en la célula formando un número determinado de parejas de cromosomas homólogos característico de cada especie, uno de los cuales se heredaba del padre y el otro de la madre. También se pudo comprobar que el número de pares de cromosomas no dependía de la complejidad del ser vivo. Asi por ejemplo, en el hombre se contabilizaron 23 pares de cromosomas, mientras que en una planta como el trigo podían encontrarse hasta 28 pares. En base a estos descubrimientos y a los estudios realizados en 1906 por el zoólogo estadounidense Thomas H.Morgan sobre los cromosomas de la mosca del vinagre (Drosophila melanogaster), se pudo elaborar la teoría cromosómica de la herencia donde se establecía de manera inequívoca la localización física de los genes en la célula. Gracias a esta teoría se pudo dar también una explicación definitiva a los casos en los que no se cumplían con exactitud las leyes de Mendel anteriormente citadas. De manera parecida a Mendel, Morgan se dedicó a cruzar de manera sistemática diferentes variedades de moscas del vinagre. Estas moscas ofrecían muchas ventajas con respecto a los guisantes ya que tienen un ciclo vital muy corto, producen una gran descendencia, son fáciles de cultivar, tienen tan sólo cuatro cromosomas y presentan características hereditarias fácilmente observables, como el color de los ojos, la presencia o ausencia de alas, etcétera. Herencia de genes ligados La investigación con las moscas del vinagre proporcionó a Morgan evidencias de que los caracteres no eran heredados siempre de forma independiente tal y como había postulado Mendel en su tercera ley. Supuso que al haber solo cuatro cromosomas diferentes, muchos genes debían estar «ligados», es decir, debían compartir el mismo cromosoma y por ello mostrar una Clara tendencia a transmitirse juntos a la descendencia. No obstante, las conclusiones realizadas por Mendel años atrás, no dejaban de ser correctas para los genes «no ligados». Solo la casualidad hizo que Mendel escogiese para los cruces de sus plantas características determinadas por genes situados en cromosomas distintos. Herencia ligada al sexo En uno de sus primeros experimentos, Morgan cruzó un macho de moscas de ojos rojos (normales) con una hembra que había encontrado casualmente y que tenia los ojos blancos. Las moscas que obtuvo en esta primera generación o F1 tenían todas los ojos rojos, tal y como se describe en la primera ley de Mendel. Pero cuando cruzó entre si estas moscas para obtener la segunda generación filial o F2, descubrió que los ojos blancos solo aparecían en las moscas macho y además como un carácter recesivo. Por alguna razón, la característica «ojos blancos» no era transmitida a las moscas hembras, incumpliendo, al menos parcialmente, la segunda ley de Mendel. Al mismo tiempo, en sus observaciones al microscopio, Morgan había advertido con extrañeza que entre los cuatro pares de cromosomas de los machos, había una pareja en la que los cromosomas homólogos no tenían exactamente la misma forma. Era como si a uno de ellos le faltase un trozo, por lo que a partir de ese momento a esta pareja se la denominó cromosomas XY. Sin embargo en la hembra, la misma pareja de cromosomas homólogos no presentaba ninguna diferencia entre ellos, por lo que se la denominó cromosomas XX. Morgan pensó que los resultados anómalos del cruzamiento anterior se debían a que el gen que determinaba el color de los ojos se encontraba en la porción que faltaba en el cromosoma Y del macho. Por tanto, en el caso de las hembras (xx) al existir dos alelos, aunque uno de ellos fuese el recesivo (ojos blancos), el carácter manifestado era el normal (ojos rojos). En los machos, sin embargo, al disponer Únicamente de un alelo (el de su único cromosoma X), el carácter recesivo si que podía ser observado. De esta manera quedaba también establecido que el sexo se heredaba como un carácter más del organismo. SÍNTESIS Una determinada característica o cualidad se manifiesta en un animal a través de un par genético, resultado de la combinación de dos pares de genes, uno del padre y otro de la madre. Cuando se combinan los genes del padre y de la madre pueden darse hasta 4 combinaciones diferentes para cada hijo, o lo que es lo mismo, cuatro tipos de descendencia para una determinada característica. La forma matemática de expresarlo podría ser: (Aa)+(Bb) = (AB); (Ab) ; (aB) ; (ab) Donde Aa, Bb son los pares genéticos de partida (de los progenitores) para una determinada cualidad (p.e. pigmento del color), y “AB, Ab, aB, ab” son los pares genéticos de cada tipo de descendencia De la misma forma una cualidad se transmite de padres a hijos, dicha cualidad también puede perderse definitivamente para parte de la descendencia, es decir, no todos los hijos de un ejemplar portarán (ni de forma dominante ni de forma recesiva), una determinada característica, de ahí la importancia que tiene el seleccionar un determinado ejemplar como semental, y precisamente el que presenta dicha cualidad, no el hermano, pariente ni cualquier otro.
 0
0
Anuncio

Descargar
Anuncio
Añadir este documento a la recogida (s)
Puede agregar este documento a su colección de estudio (s)
Iniciar sesión Disponible sólo para usuarios autorizadosAñadir a este documento guardado
Puede agregar este documento a su lista guardada
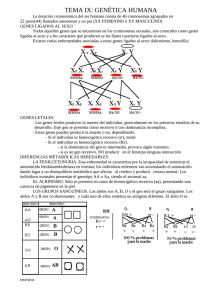
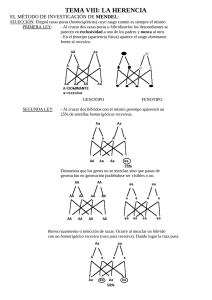
Iniciar sesión Disponible sólo para usuarios autorizados