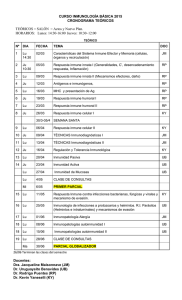
ACTUALIZACIÓN Respuesta inmune innata y sus implicaciones fisiopatológicas D. Díaz Martín*,a,b, M. Úbeda Canteraa, A. López Suáreza y *M. Álvarez de Mon Sotoc a Laboratorio de enfermedades del sistema Inmune y Oncología. Departamento de Medicina. Universidad de Alcalá. Alcalá de Henares. Madrid. España. Centro de Investigación Biomédica en Red de Enfermedades Hepáticas y Digestivas (CIBERehd). Instituto de Salud Carlos III. Madrid. España. c Servicio de Enfermedades del Sistema Inmune y Oncología. Hospital Universitario Príncipe de Asturias. Universidad de Alcalá. Alcalá de Henares. Madrid. España. b Palabras Clave: Resumen - Receptores de reconocimiento de patrones Generalidades. La inmunidad innata o natural constituye la primera línea de defensa contra los microorganismos. El término “innato” hace referencia a que los sensores implicados en el reconocimiento de los patógenos están codificados por genes en línea germinal que no sufren reordenamiento somático para generar variantes. - Patrones moleculares asociados a patógenos - Patrones moleculares asociados a daño Componentes de la inmunidad innata. Las armas del sistema inmune innato incluyen mecanismos de defensa celulares y moleculares que existen antes de que tenga lugar una infección, lo que les permite responder frente a ella con rapidez. Los sensores de la inmunidad innata son específicos de las estructuras comunes a grupos de patógenos relacionados, pero no pueden distinguir diferencias sutiles entre ellos. Receptores de la inmunidad innata. En el primer bloque analizamos los receptores que utiliza la inmunidad innata para identificar moléculas características de grupos de patógenos, así como de daño y estrés celular, que se denominan en su conjunto receptores de patrones moleculares. Moléculas de la inmunidad innata. En el segundo bloque examinamos en profundidad las moléculas solubles que median la inmunidad innata. Estas moléculas actúan rápidamente contra patógenos extracelulares de tres maneras importantes: como opsoninas, como agentes líticos y como agentes proinflamatorios. Keywords: Abstract - Pattern recognition receptors Innate immune response and their pathophysiological implications - Pathogen-associated molecular patterns Overview. Innate or natural immunity is the first line of defense against microorganisms. The term "innate" refers to the sensors involved in the pathogen recognition are encoded by genes in germline suffering non-somatic rearrangement to generate variants - Damage associated molecular patterns Components of the innate immunity. The weapons of the innate immune system include molecular and cellular defense mechanisms that exist before an infection takes place, allowing them to respond to it quickly. Sensors of the innate immunity are specific structures common to groups of related pathogens, however they cannot distinguish subtle differences between them. Receptors of the innate immunity. The first block analyzes receptors used by the innate immunity to identify characteristic molecules of groups of pathogens, as well as damaged and stressed cells, named pattern recognition receptors. Molecules of the innate immunity. The second block examines in depth the soluble molecules that mediate innate immunity. These molecules act quickly against extracellular pathogens in three important ways: as opsonins, lytic agents and as pro-inflammatory agents. *Correspondencia Correo electrónico: [email protected] 1388 Medicine. 2017;12(24):1388-97 RESPUESTA INMUNE INNATA Y SUS IMPLICACIONES FISIOPATOLÓGICAS Generalidades de la respuesta inmune innata Durante años, los inmunólogos han ignorado en gran medida al sistema inmune innato (SII). Sin embargo, el estudio del sistema inmune adaptativo (SIA) ha conducido a una nueva concepción del papel que tiene el SII, no solo como un sistema pasivo y extremadamente rápido de segunda línea de defensa (las barreas físicas serían la primera línea) sino también como un activador que inicia, dirige y regula al SIA. La importancia de la veloz respuesta del SII a patógenos comunes como las bacterias es fácil de entender si pensamos en las consecuencias de una infección incontrolada. Las bacterias proliferan muy rápidamente. De hecho, una bacteria dobla su número en unos 30 minutos. Esto implica que una sola bacteria puede generar 100 billones (1014) de bacterias en un solo día. Un litro de medio de cultivo que contenga 100 billones de bacterias es tan denso y turbio que no se puede ver a través de él; efectivamente, una sola bacteria que prolifere un día generará un cultivo denso de unos 100 litros. Si recordamos que nuestro volumen total de sangre son unos cinco litros, podemos apreciar ahora lo que una infección bacteriana incontrolada podría provocar a un ser humano. Comenzaremos la actualización profundizando en las generalidades de la respuesta inmune innata, continuaremos con los receptores de patrones moleculares de la inmunidad innata y finalizaremos analizando en profundidad las moléculas solubles que median la inmunidad innata. El análisis del componente celular de la respuesta inmune innata se describe en otra actualización de esta misma unidad temática (actualización 1). Componentes y características Como suele suceder en la ciencia, inicialmente se estableció una dicotomía clara entre el SII y el SIA. Sin embargo, actualmente esta frontera es muy difusa, pues ambos sistemas comparten mecanismos efectores, células, receptores y moléculas solubles como veremos más adelante. El término «innato» hace referencia a que los sensores implicados en el reconocimiento de los patógenos están codificados por genes en línea germinal que no sufren reordenamiento somático para generar variantes y que reconocen estructuras moleculares compartidas por grupos de microorganismos patógenos, siendo a menudo esenciales para la supervivencia de los mismos. Las armas del SII incluyen mecanismos de defensa celulares y moleculares que existen antes de la infección y pueden responder con rapidez a ella. Estos mecanismos reaccionan con patógenos y células dañadas, y responden de una forma prácticamente idéntica a infecciones repetidas. Los sensores de la inmunidad innata son específicos de las estructuras comunes a grupos de patógenos relacionados y no pueden distinguir diferencias sutiles entre ellos. Los principales componentes de la inmunidad innata son: a) componentes celulares: células fagocíticas (neutrófilos, macrófagos), células dendríticas (DC), linfocitos natural killer (NK) y otras células linfocíticas innatas; b) receptores de patrones moleculares asociados a patógenos o a daño y c) sustancias químicas antimicrobianas, proteínas solubles y mediadores de la inflamación (tabla 1). La inmunidad innata y la adaptativa (actualización 4) se integran perfectamente para constituir un sistema robusto que nos protege contra los microorganismos patógenos. Para ello, presentan características particulares que las definen y que es instructivo destacar. Las respuestas inmunitarias innatas frente a un microbio son inmediatas y no requieren una exposición previa. Además, las moléculas implicadas en el reconocimiento y en su acción efectora están codificadas en línea germinal. Por el contrario, las respuestas inmunitarias adaptativas tienen lugar varios días después de la exposición al patógeno y se desarrolla mediante el crecimiento de clones específicos en respuesta al contacto. No se genera memoria inmune y, por ello, no se produce ningún cambio apreciable ni en la calidad, ni en la magnitud de la respuesta inmunitaria innata frente a un microbio tras una exposición repetida. Por el contrario, en las respuestas inmunitarias adaptativas la exposición previa a un microbio potencia su rapidez, magnitud y eficacia. El repertorio de reconocimiento de la inmunidad innata es muy limitado y no es clonal. Se calcula que el SII reconoce solo unos 1.000 productos de microbios y células dañadas. Como hemos visto, estos receptores están codificados por genes en línea germinal que no sufren reordenamiento somático. Por el contrario, el repertorio de reconocimiento de la inmunidad adaptativa es prácticamente ilimitado, incluso capaz de reconocer moléculas actualmente inexistentes, cuyos receptores presentan una distribución clonal y se originan por reordenamiento genético somático y que, por tanto, no se heredan. Evolución del sistema inmune innato Se estima que más del 99% de la vida en la Tierra solo presenta SII y desde el punto de vista filogenético es la parte más antigua del sistema inmune1. Este sistema coevolucionó junto con los patógenos para proteger a los organismos multicelulares de las infecciones. De hecho, algunos de sus componentes en mamíferos son muy similares a los componentes de plantas e insectos. Por ejemplo, las defensinas, péptidos tóxicos para bacterias y hongos, se encuentran en plantas y mamíferos y tienen en esencia la misma estructura terciaria en ambas formas de vida. Otro ejemplo son los receptores del tipo toll que reconocen estructuras conservadas de patógenos. Estos receptores se encuentran en todos los organismos pluricelulares desde insectos hasta mamíferos. Casi la totalidad de los mecanismos implicados en la inmunidad innata que desarrollaremos en esta actualización aparecieron muy pronto en la evolución, concretamente cuando evolucionaron los primeros organismos multicelulares complejos, hace unos 750 millones de años. El SIA, por el contrario, evolucionó con claridad reconocible en los vertebrados que aparecieron hace unos 350-500 millones de años. Medicine. 2017;12(24):1388-97 1389 ENFERMEDADES DEL SISTEMA INMUNE Y REUMATOLÓGICAS (I) TABLA 1 Receptores y moléculas solubles de la inmunidad innata Denominación Estructura Localización Ejemplos Ligandos (PAMP, DAMP) Receptores Receptores tipo toll (TLR) Dominios LRR y TIR Membrana plasmática y membrana de endosomas y endolisosomas TLR 1-10 PAMP: LPS, ácido lipoteicoico, la flagelina y ácidos nucleicos víricos Receptores lectina tipo C (CLR) Lectina dependiente de Ca2+ Membrana plasmática Receptor de manosa Azúcares terminales de la superficie de microbios DAMP: HSP y HMGB1 Betaglucanos unidos en posición 1 y 3 Dectinas Receptores tipo NOD (NLR) Dominio LRR y dominio NACHT Citosol NOD1 y NOD2 Ácido diaminopimélico y dipéptido muramilo bacterianos PAMP: flagelina, dipéptido muramilo, LPS y ARN vírico NLRP (inflamasoma) DAMP: cristales (urato monosódico, LDL), cambios en ATP e iones Receptores tipo RIG (RLR) Dominios CARD y ARN-helicasa Citosol RIG-I y MDA5 ARN vírico Detectores citosólicos de ADN (CDS) Dominio PYHIN y dominio HIN-200 Citosol AIM2 ADN vírico y bacteriano Receptores scavenger (SR) Estructuralmente heterogéneos Membrana plasmática de células fagocíticas CD36 Catelicidinas LPS Defensinas Membranas microbianas DAI Vía STING PAMP: microorganismos o partículas extrañas DAMP: células apoptóticas y proteínas dañadas Moléculas solubles Péptidos de defensa del huésped (HDP) Péptidos cortos, catiónicos y anfipáticos Plasma Complemento Zimógenos inactivos Plasma C3 Membranas microbianas Pentraxinas Proteínas pentaméricas Plasma Proteína C reactiva Residuos de fosfocolina Colectinas Dominio tipo colágeno y un dominio de lectina tipo C Plasma y alveolos pulmonares Lectina de unión a manosa Manosa en superficie de patógenos Proteína A surfactante Estructuras microbianas Dominio tipo colágeno y dominios tipo fibrinógeno Plasma Ficolina N-acetil glucosamina y ácido lipoteicoico Ficolinas Dominios repetidos ricos en leucinas (LRR), TIR (dominios toll/IL-1 receptor), patrones moleculares asociados a patógenos (PAMP), patrones moleculares asociados a daño (DAMP), LPS (lipopolisacárido), HSP (proteínas de choque térmico), caja del grupo de movilidad alta 1 (HMGB1). Funciones de la inmunidad innata La inmunidad innata presenta tres funciones principales que son esenciales para la defensa frente a los agentes patógenos, el daño y la renovación tisular. La primera función es impedir, controlar y/o eliminar la infección por microorganismos patógenos. Esto se pone de manifiesto en modelos experimentales que muestran cómo las deficiencias, la inhibición o la eliminación de varios mecanismos de la inmunidad innata incrementan la propensión a las infecciones, incluso cuando el SIA está intacto y funcional. Muchos microbios patógenos han desarrollado estrategias para resistir la inmunidad innata y estas estrategias son cruciales para la virulencia de los mismos. La mayoría del contenido de esta actualización versa sobre esta primera función. La segunda función es eliminar las células dañadas e iniciar el proceso de reparación tisular. Estos mecanismos reconocen y responden a moléculas que las células estresadas, dañadas y muertas producen, liberan o acumulan. Estas respuestas innatas pueden darse en el contexto de la infección o del daño estéril (daño que sucede en células y tejidos en ausencia de infección). Alteraciones en esta función de la respuesta inmune innata pueden tener muchas implicaciones patológicas como veremos más adelante. La tercera función es iniciar, dirigir y regular las respuestas inmunitarias adaptativas, haciéndolas eficaces frente a diferentes tipos de microbios. La inmunidad innata propor1390 Medicine. 2017;12(24):1388-97 ciona las señales de peligro que alertan y polarizan al SIA para que responda de la manera más adecuada en función del modo de vida de ese patógeno particular. Esta flexibilidad funcional permite adecuar la respuesta a cada tipo de patógeno (o célula tumoral) y generar una memoria inmune adecuada a cada situación. Existen nuevas estrategias terapéuticas que tienen como diana esta plasticidad funcional. Receptores de la inmunidad innata El SII detecta la presencia de patógenos y/o el daño e inicia los mecanismos necesarios para eliminar esta amenaza potencialmente infecciosa. La detección de la amenaza se lleva a cabo mediante los receptores de reconocimiento de patrones (PRR —pattern-recognition receptors—). Los PRR están codificados en la línea germinal, vigilan tanto el espacio extra como el intracelular y, cuando se activan, inician la transducción de señales que promueven las funciones antimicrobiana y proinflamatoria de las células que los expresan2. La expresión de los PRR no está restringida a células del SII (macrófagos, neutrófilos, DC, etc.) sino que pueden presentarlos las células epiteliales que componen la barrera entre el cuerpo y el ambiente externo y muchos otros tipos de células que ocupan los tejidos y los órganos. Los PRR reconocen principalmente moléculas conservadas de microbios denominadas patrones moleculares asociados a patógenos (PAMP —pathogen-associated molecular pat- RESPUESTA INMUNE INNATA Y SUS IMPLICACIONES FISIOPATOLÓGICAS terns—) pero también pueden identificar señales de peligro o daño provenientes de células propias que son expresadas o liberadas en respuesta al estrés, daño tisular y/o muerte celular por necrosis3. A estas moléculas endógenas se las denomina patrones moleculares asociados a daño (DAMP, —damage associated molecular patterns—) (tabla 1). Los PAMP presentan estructuras químicas muy diversas, pero comparten tres características: a) están presentes en los microorganismos, pero no en sus huéspedes; b) son esenciales para la supervivencia o patogenicidad de los microorganismos; c) muchos de ellos son compartidos por microorganismos diferentes. Estructuralmente, los PAMP pueden ser lípidos, hidratos de carbono, proteínas, lipoproteínas/glicoproteínas o ácidos nucleicos microbianos como detallaremos más adelante. Los DAMP pueden producirse como resultado del daño celular causado por infecciones, pero también pueden indicar una lesión estéril de las células causada por alguna otra razón, como toxinas químicas, quemaduras, traumatismos o reducción del riego sanguíneo. Las células que mueren por apoptosis no suelen liberar DAMP, pero las que lo hacen por necrosis sí. En algunos casos, se estimula a las células sanas del sistema inmunitario para que produzcan y liberen algunos DAMP, también llamados alarminas, que aumentan la respuesta inmunitaria innata frente a las infecciones. En esta sección examinaremos los principales PRR utilizados por el SII centrándonos en su especificidad, localización y funciones. Posteriormente, describiremos las proteínas de la sangre y los líquidos extracelulares que reconocen PAMP y que facilitan la eliminación de los microbios de la sangre y de los líquidos extracelulares, pues aumentan su captación por los fagocitos y activan los mecanismos microbicidas. La mayoría de los PRR pueden clasificarse en una de las cinco familias definidas por dominios de homología4. Estas cinco familias son: los receptores tipo toll (TLR —toll-like receptors—), los receptores lectina tipo C (CLR —C-type lectin receptors—), los receptores tipo NOD (NLR —nucleotidebinding oligomerization domain receptors—), los receptores tipo RIG (RLR, —RIG-I like receptors—) y los detectores citosólicos de ADN (CDS, —citosolic DNA sensors—). Estas familias pueden agruparse en dos clases principales: los receptores unidos a membrana y los receptores intracelulares no unidos a membrana. La primera clase está constituida por los TLR y los CLR que se encuentran anclados en la membrana plasmática o en compartimentos endocíticos. Estos receptores detectan la presencia de ligandos microbianos en el espacio extracelular y en endosomas. Los NLR, RLR y CDS forman el segundo grupo y están localizados en el citoplasma, donde detectan la presencia de patógenos intracelulares (tabla 1). El componente principal de la respuesta inmune innata inducida por PRR es transcripcional y conduce a la producción de citoquinas proinflamatorias e interferones (IFN). Estos mensajeros químicos son esenciales para iniciar las respuestas inmunes, tanto innatas como adaptativas. La activación de los PRR también inicia una respuesta no transcripcional como la inducción de fagocitosis, autofagia, muerte celular y procesamiento de citoquinas5,6. Estas respuestas inmunes innatas transcripcionales y no transcripcionales es- tán vinculadas a la detección microbiana por PRR mediante vías de transducción de señales delicadamente controladas. La coordinación de esas vías de señalización orquesta la respuesta inmune desde el control inicial de la infección hasta el desencadenamiento de una respuesta inmune adaptativa apropiada. El estudio exhaustivo de estas vías de señalización queda fuera del alcance de esta actualización. Los receptores tipo toll Los TLR son el grupo de PRR más estudiado y están muy conservados a lo largo de la evolución. Son receptores transmembrana de tipo I que presentan dominios repetidos ricos en leucinas (LRR —leucine-rich repeat—) y dominios TIR (toll/IL-1 receptor). Los dominios LRR les confieren su capacidad para interaccionar con distintos tipos de ligandos y los dominios TIR intracitoplasmáticos inician la vía de señalización. Se han identificado 10 miembros de la familia TLR en seres humanos y 13 en ratón y se conocen tanto sus ligandos como su señalización intracelular4,7. El reconocimiento mediado por TLR puede ocurrir en la membrana plasmática o en la membrana de endosomas y endolisosomas. TLR1, TLR2, TLR4, TLR5 y TLR6 se localizan principalmente, aunque no exclusivamente, en la membrana plasmática y reconocen componentes microbianos como lípidos, lipoproteínas, LPS y proteínas. Por el contrario, TLR3, TLR7, TLR8 y TLR9 se localizan en compartimientos vesiculares intracelulares y están implicados principalmente en el reconocimiento de ácidos nucleicos (fig. 1). Los TLR reconocen moléculas expresadas por los microbios pero que no se expresan en células sanas y, si lo hacen, será en otro compartimento celular. Ejemplos de productos bacterianos que se unen a los TLR son el LPS (Gram negativas) y el ácido lipoteicoico (Gram positivas) y la flagelina, presente en los flagelos de las bacterias móviles. Ejemplos de ácidos nucleicos que son ligandos para los TLR producidos por los virus son: a) los ARN bicatenarios, que componen los genomas de algunos virus y se generan durante el ciclo vital de la mayoría de los virus ARN, pero que no producen las células eucariotas y b) los ARN unicatenarios que se distinguen de los transcriptos de ARN unicatenario citoplásmicos celulares por su localización dentro de los endosomas, por su elevado contenido en guanosina y uridina, y por los dinucleótidos CpG no metilados que son frecuentes en los genomas de los procariotas e infrecuentes en los de los vertebrados (fig. 1). Como vimos al principio de la actualización, los TLR también reconocen moléculas propias cuya expresión o localización refleja daño celular. Ejemplos de estas moléculas son las proteínas de choque térmico (HSP) y la caja del grupo de movilidad alta 1 (HMGB1), una proteína abundante ligadora de ADN implicada en la transcripción y reparación del mismo8. La unión del ligando a los TLR da lugar a la iniciación de la transducción de señales por varias vías y a la activación de factores de transcripción que, finalmente, inducen la expresión de genes cuyos productos son importantes para las respuestas inflamatoria (actualización 2) y antivírica (interferones tipo I). Medicine. 2017;12(24):1388-97 1391 ENFERMEDADES DEL SISTEMA INMUNE Y REUMATOLÓGICAS (I) nio NACHT (proteína inhibidora de la apoptosis neuronal [NAIP], CIITA, HET-E y TP1), que permite al NLR LPS Flagelina Lipopéptidos Lipopéptidos Peptidoglicanos unirse a otro y formar oligómeros y un TLR2 TLR4 TLR5 TLR1: TLR2 TLR2: TLR6 dominio efector o de interacción proteína-proteína que forma complejos Membrana que transmiten la señal y que, de plasmática acuerdo con su naturaleza, permite clasificar los NLR. Concretamente, existen tres dominios efectores que permiEndosoma ten dividir los NLR en tres subfamilias: dominios CARD (dominio de reclutaVirus Bacterias ARNbc miento de caspasa), pirina y BIR (doARNmc CpG ARNmc minios repetidos IAP de baculovirus). TLR3 TLR9 NOD1 y NOD2 contienen domiTLR7 TLR8 nios CARD y reconocen el peptidoglicano, un componente de la pared celular bacteriana. NOD1 reconoce el ácido diaminopimélico (DAP) presente en el Fig. 1. Localización celular y ligandos de los TLR (receptores tipo toll) humanos. Los TLR son receptores peptidoglicanos de todas las bacterias transmembrana de tipo I que presentan dominios repetidos ricos en leucinas (LRR —leucine-rich repeat—) y dominios TIR (toll/IL-1 receptor). Los dominios LRR les confieren su capacidad para interaccionar con Gram negativas y solo en algunas distintos tipos de ligandos y los dominios TIR intracitoplasmáticos inician la vía de señalización. El reconoGram positivas (Listeria y Bacillus), cimiento mediado por TLR puede ocurrir en la membrana plasmática o en la membrana de endosomas y enmientras que NOD2 reconoce una dolisosomas. TLR1, TLR2, TLR4, TLR5 y TLR6 se localizan principalmente, aunque no exclusivamente, en la membrana plasmática y reconocen componentes microbianos como lípidos, lipoproteínas, LPS y proteínas. molécula diferente llamada dipéptido Por el contrario, TLR3, TLR7, TLR8 y TLR9 se localizan en compartimientos vesiculares intracelulares y están muramilo un motivo del peptidoglicaimplicados principalmente en el reconocimiento de ácidos nucleicos. no presente en Gram negativos y Gram positivos12. Tanto la estimulaLos receptores lectina tipo C ción de NOD1 como de NOD2 forma un complejo transductor de señales que se ha denominado La familia de los CRL se denomina así porque estos recepseñalosoma de NOD. Este señalosoma conduce a la activatores se unen a glúcidos (las lectinas son proteínas que recoción del factor de transcripción NFNB el cual estimula la nocen azúcares) de una forma que depende del Ca2+ (tipo C). producción de quimioquinas y citoquinas que inician la respuesta proinflamatoria necesaria para eliminar el patógeno. En general, los CRL reconocen estructuras glucídicas que se NOD1 y NOD2 parecen importantes en las respuestas inencuentran en las paredes celulares de los microorganismos, munitarias innatas frente a las bacterias patógenas del tubo pero no en las células de los mamíferos9. El receptor de manosa es uno de los CRL más estudiado. digestivo, como Helicobacter pylori y Listeria monocytogenes. Este receptor se une a azúcares terminales de la superficie de Los receptores NRL constan de 14 miembros que contienen un dominio pirina (NLRP) y forman complejos mulmicrobios, como la D-manosa, la L-fucosa y la N-acetil-Dtiproteicos transductores de la señal de alto peso molecular glucosamina. Los glúcidos de las células eucariotas suelen denominados inflamasomas13 (fig. 2). Los miembros más coterminar con galactosa y ácido siálico. Parece ser que estos nocidos son NLRP1, NLRP3 y NLRC4. Los NLRP resreceptores no generan transducción de señales en la célula ponden a DAMP y PAMP citosólicos y generan formas actique los expresa y son un paso previo importante para iniciar vas de las citoquinas proinflamatorias IL-1 e IL-18. Los la fagocitosis del microbio. PAMP que activan el inflamasoma son moléculas bacterianas Las dectinas (dectina 1 y 2) son receptores de las DC, como la flagelina, el dipéptido muramilo, el LPS y las toxinas macrófagos y neutrófilos que reconocen beta-glucanos unidos en posición 1 y 3 presentes en hongos, algunas bacterias formadoras de poros, así como los ARN bacteriano y vírico. y plantas. Se unen a una gama de hongos patógenos como Los DAMP que activan el inflamasoma son las sustancias Candida, Aspergillus, Coccidiodes y Pneumocystis y bacterias pacristalinas que pueden proceder del ambiente, como el amianto y la sílice, o pueden tener un origen endógeno, como el tógenas como Mycobacteria10. Su activación promueve la fagourato monosódico, el pirofosfato de calcio deshidratado y el citosis y producción masiva de citoquinas proinflamatorias. colesterol cristalizado. Otros estímulos endógenos de la activación del inflamasoma son: el ATP extracelular, quizás liberado por las células muertas y transportado al citoplasma de Los receptores tipo NOD la célula respondedora, las disminuciones del potasio intracelular provocadas por algunas toxinas bacterianas y las espeLa segunda familia más importante de PRR son los NLR. cies reactivas del oxígeno que se producen a menudo duranEstos receptores representan la contraparte citosólica de los te la lesión celular. TLR e incluyen hasta el momento 23 miembros en seres El hallazgo de que muchos DAMP activan el inflamasohumanos11. Los NLR presentan tres dominios distintos: un ma ha cambiado en gran medida la compresión de ciertas dominio LRR encargado de detectar y unir PAMP; un domiBacterias 1392 Medicine. 2017;12(24):1388-97 RESPUESTA INMUNE INNATA Y SUS IMPLICACIONES FISIOPATOLÓGICAS Cristales: Amianto, silice Urato monosódico Colesterol K+ ATP Toxinas Medio extracelular Fagocitosis frustrada Sales de aluminio Proteína β-amiloide K+ bajo Ca2+ alto Dipéptido muramilo LPS ARN bacteriano y vírico Flagelina Ruptura de lisosoma ROS Catepsina B Citoplasma Inflamasoma Núcleo Gen Pro-IL-1β Piroptosis Pro-IL-1β IL-1β Inflamación TLRs IL-1β Fig. 2. El inflamasoma. En la figura se muestra que la activación del inflamasoma puede ocurrir en respuesta a DAMP (patrones moleculares asociados a daño) y PAMP (patrones moleculares asociados a patógenos) citosólicos, generando así formas activas de las citoquinas proinflamatorias IL-1 e IL-18. Los PAMP que activan el inflamasoma son moléculas bacterianas como la flagelina, el dipéptido muramilo, el LPS (lipopolisacárido) y las toxinas formadoras de poros, así como los ARN bacteriano y vírico. Los DAMP que activan el inflamasoma son las sustancias cristalinas que pueden proceder del ambiente, como el amianto y la sílice, o pueden tener un origen endógeno, como el urato monosódico, el pirofosfato de calcio deshidratado y el colesterol cristalizado. Otros DAMP son: el ATP, las disminuciones del potasio intracelular provocadas por algunas toxinas bacterianas, la ruptura lisosomal y las especies reactivas del oxígeno que se producen a menudo durante la lesión celular. La activación del inflamasoma puede inducir la muerte celular por piroptosis. enfermedades inflamatorias. Por ejemplo, la gota es una enfermedad inflamatoria producida por una acumulación de cristales de urato monosódico que activan el inflamasoma, por lo que se usan antagonistas de la IL-1 para los casos de gota graves resistentes a la terapia convencional. Otro ejemplo son los síndromes autoinflamatorios que se caracterizan por una inflamación espontánea sin un desencadenante claro. Estos pacientes también pueden ser tratados satisfactoriamente con antagonistas de la IL-1. Por último, hay que destacar que la activación de la caspasa 1 por el inflamasoma puede inducir un modo fisiológico de muerte celular programada denominado piroptosis que culmina con la lisis de la célula y la liberación de contenido citosólico al medio extracelular, entre el que se incluye grandes cantidades de IL-1E e IL-18 (fig. 2). Los receptores tipo RIG Los RLR son receptores citoplasmáticos de ARN vírico que presentan dominios CARD y ARN-helicasa4,14. Están presentes en casi todos los tipos celulares y les permiten responder a la infección vírica mediante el reconocimiento del ARN del virus. La señalización a través de los RLR conduce principalmente a la producción de interferones tipo I por la célula infectada, así como de otras citoquinas inflamatorias. Los dos RLR más estudiados son RIG-I (gen inducible por ácido retinoico I) y MDA5 (gen asociado a la diferenciación del melanoma 5) (tabla 1). Ambos receptores reconocen ARN vírico de diferentes grupos de virus, pero con cierta especificidad que parece que viene determinada por la longi- tud del ARN vírico. RIG-1 suele reconocer el ARN bicatenario corto (menos de 1 kb), especialmente cuando contiene extremos 5’-trifosfato (que no está presente en células eucariotas), mientras que MDA5 reconoce el ARN bicatenario mayor de 2 kb. Detectores citosólicos de ADN Los CDS son receptores ubicados en el citosol que participan en la detección de ADN intracelular y que activan rutas de señalización que inician respuestas antimicrobianas mediante la producción de interferones tipo I y la autofagia. La búsqueda de receptores citosólicos que reconocen el ADN ha implicado a un número desconcertante de genes involucrados en el reconocimiento de este ácido nucleico y en la regulación de la respuesta a IFN15. Una de las rutas más importante es la vía STING (stimulator of IFN genes) (tabla 1). Concretamente, STING es una proteína transmembrana localizada en el retículo endoplásmico que activa de forma directa (receptor) e indirecta (adaptador) el ADN microbiano en el citosol. En la vía indirecta, el ADN citosólico se une a la enzima sintasa del GMP-AMP cíclico (GASc) que sintetiza un dinucleótido cíclico llamado GMP-AMP cíclico (GAMPc) después de encontrarse con el ADN. Posteriormente, GAMPc interactuará con STING que tiene la capacidad de unirse directamente a dinucleótidos cíclicos. Esta ruta de activación de STING induce la producción de interferones tipo I por la célula. En la vía directa, STING reconoce dinucleótidos cíclicos producidos comúnmente por bacterias como segundos mensajeros. La activación de Medicine. 2017;12(24):1388-97 1393 ENFERMEDADES DEL SISTEMA INMUNE Y REUMATOLÓGICAS (I) STING también estimula la autofagia celular con el objetivo de dirigir los microbios a los lisosomas para ser degradados. Otras moléculas muy importantes en la detección de ADN citosólico son los AIM2-like receptor (ALR) que, según varios autores, constituyen por sí mismos un nuevo grupo de receptores de patrones4. Cada uno de estos receptores presenta un dominio PYHIN que permite las interacciones proteína-proteína y un dominio HIN-200 de unión al ADN. El miembro fundador de esta familia, AIM2 (absent in melanoma 2), interactúa con el adaptador ASC y promueve la formación del inflamasoma tras la detección de ADN intracelular. Por último, el activador dependiente del ADN de los factores reguladores del IFN (DAI) se une al ADN de varias fuentes microbianas y activa al IRF3, lo que conduce a una respuesta mediada por IFNs tipo I. Por otro lado, el DAI también puede activar NF-kB. Otros receptores de reconocimiento de patrones Los receptores scavenger (SR —scavenger receptors—) fueron identificados originalmente por su capacidad para reconocer y eliminar las lipoproteínas modificadas. Sin embargo, ahora se aprecia que llevan a cabo una impresionante gama de funciones, no solo debidas al amplio repertorio de receptores que constituyen esta familia, sino a su capacidad de asociarse con varios correceptores16. Se expresan principalmente en células mieloides (monocitos/macrófagos y DC) pero también en ciertos endotelios y epitelios (tabla 1). Los SR son estructuralmente muy heterogéneos. Se subdividen en ocho clases y, aunque los miembros de cada clase comparten características estructurales, hay poca o ninguna homología entre ellos. La fusión de los SR en una superfamilia es sobre todo debida a que comparten propiedades funcionales. En general, los SR identifican y eliminan entidades indeseables mediante el reconocimiento de moléculas propias modificadas (por ejemplo, células apoptóticas, desechos ricos en mineral o proteínas dañadas) o a través del reconocimiento de moléculas no propias (por ejemplo, microorganismos o partículas extrañas). La eliminación se realiza a menudo por simple endocitosis, pero puede implicar procesos más complejos, como la macropinocitosis o fagocitosis, que requiere una elaborada transducción de señal. Otras funciones de estos receptores multifuncionales incluyen la presentación del antígeno y la adhesión celular. Al participar en el reconocimiento y la internalización de la LDL oxidada y la proteína E-amiloide, así como en el transporte de ácidos grasos, los SR han sido implicados en enfermedades tan diversas como la ateroesclerosis, la diabetes mellitus tipo 2 y la enfermedad de Alzheimer, siendo el CD36 uno de los SR más estudiado. El receptor para péptido formilado 1 (FPR1 —formyl peptide receptor 1—), altamente expresado por células fagocíticas y leucocitos circulantes, reconoce péptidos bacterianos que contienen N-formilmetionina y dirige estas células a los tejidos dañados por estos microorganismos. Debido a que todas las proteínas bacterianas y algunas proteínas de mamí1394 Medicine. 2017;12(24):1388-97 feros (solo aquellas sintetizadas dentro de la mitocondria) comienzan con N-formilmetionina, FPR1 permite a los fagocitos detectar y responder preferentemente a las proteínas bacterianas. Moléculas solubles de la inmunidad innata Se conocen una gran cantidad de moléculas solubles efectoras que están presentes en la sangre y el líquido extracelular que median el reconocimiento de los microbios y promueven respuestas inmunes innatas. Estas moléculas ejercen su acción rápidamente contra patógenos extracelulares y actúan de tres maneras importantes. La primera como opsoninas, potenciando de esta manera la capacidad de las células fagocíticas para eliminar el patógeno. La segunda lisando directamente los microbios mediante complejos de ataque a membrana. La tercera promoviendo respuestas inflamatorias y ejerciendo como moléculas quimioatrayentes. A las moléculas solubles también se las denomina rama humoral de la inmunidad innata por analogía con la respuesta mediada por anticuerpos de la inmunidad adaptativa, aunque esta clasificación ya ha quedado obsoleta. Las moléculas solubles de la inmunidad innata las podemos clasificar en péptidos de defensa del huésped (HDP —host defence peptides—), el complemento, pentraxinas, colectinas y ficolinas (tabla 1). Péptidos de defensa del huésped Los HDP son péptidos cortos, catiónicos y anfipáticos con secuencias diversas que son producidos por diferentes células y tejidos en todas las formas de vida complejas17. Estudios recientes han demostrado una amplia gama de actividades directas de estos péptidos en la modulación de las funciones de células y tejidos propios. Esto ha llevado a la denominación de estos péptidos como HDP, en lugar de péptidos antimicrobianos, pues define mejor sus papeles multifacéticos que van desde su capacidad inmunomoduladora hasta, en algunas circunstancias, su capacidad antimicrobiana. Esta capacidad inmunomoduladora queda patente al observarse cómo la expresión de más de 900 genes cambia si se estimulan los monocitos humanos con algunos de estos HDP17. Existen dos familias de HDP que presentan estructuras diferentes: las catelicidinas y las defensinas. Las catelicidinas son producidas principalmente por células epiteliales de barreras como la piel, el tubo digestivo y el aparato respiratorio, así como por células del sistema inmune como neutrófilos, macrófagos, DC, NK y mastocitos18. Sus efectos están muy extendidos y generan una amplia gama de respuestas en una amplia variedad de tipos celulares (tabla 1). Estos abarcan desde la citotoxicidad directa, hasta actividades pro y antiinflamatorias que varían dependiendo del tipo de célula y estímulos inflamatorios que estén presentes. Concretamente, el fragmento C terminal, llamado LL-37, puede unirse al LPS y neutralizarlo. Por otro lado, el LL-37 también desempeña una función antiinflamatoria al unirse al ADN y RESPUESTA INMUNE INNATA Y SUS IMPLICACIONES FISIOPATOLÓGICAS bloquear la activación del inflamasoma AIM2. Otros efectos de las catelicidinas incluyen: modulación inmune antiinfecciosa (por ejemplo, expresión inducida de varias quimioquinas), actividad quimioatrayente directa de varias células inmunes, actividad antiinflamatoria, curación de heridas, actividad proangiogénica, actividad proapoptótica en algunos tipos celulares como células epiteliales y células T reguladoras, actividad antiapoptótica en neutrófilos, degranulación de mastocitos y actividad adyuvante. Las defensinas son pequeños péptidos catiónicos, entre 29 y 34 aminoácidos, que contienen tres enlaces disulfuro intracatenarios, así como regiones catiónicas e hidrofóbicas. Las defensinas humanas se dividen en dos clases principales según las uniones de los puentes disulfuro, D-defensinas y E-defensinas19. Hay seis miembros en la familia D-defensinas de HDP que se subdivide en péptidos de neutrófilos humanos (HNP), que son las D-defensinas 1-4, y la D-defensina humana 5 (HD5) y HD6 (tabla 1). Los HNP son producidos por los neutrófilos, así como por monocitos y linfocitos NK. Las HD5 y HD6 se expresan en las células de Paneth del intestino delgado, al igual que en células epiteliales de las vías respiratorias, gastrointestinales y del tracto reproductivo de la mujer. Las E-defensinas humanas (HBD) están ampliamente distribuidas por todo el organismo; son expresadas por las células epiteliales y son fácilmente producidas por monocitos, macrófagos y DC20. Las defensinas matan a los microbios mediante diversos mecanismos, muchos de los cuales dependen de su capacidad para insertarse en las membranas microbianas e interrumpir sus funciones. Por otro lado, y de manera similar a las catelicidinas, las defensinas humanas presentan un papel pro y antiinflamatorio en el sistema inmune, aunque suelen ser menos potentes. Por ejemplo, los neutrófilos que mueren por apoptosis o necrosis liberan HNP en el medio circundante. La presencia de HNP puede limitar la respuesta proinflamatoria interrumpiendo la liberación de óxido nítrico y citoquinas inflamatorias por macrófagos. Por el contrario, los HNP liberados por los neutrófilos pueden también aumentar la fagocitosis bacteriana mediada por macrófagos estimulando la producción de citoquinas como TNF e IFN-J por macrófagos, que a su vez conduce a la expresión incrementada de CD32 (también conocido como FcJRIIB) y CD64 (también conocido como FcJRI). Otras actividades de los HNP son: actividad quimioatrayente para múltiples tipos celulares; inducción de la producción de citoquinas y quimioquinas; actividad antiinflamatoria; actividad pro o antiangiogénica; actividad cicatrizante; promoción de la homeostasis intestinal; actividad pro-apoptótica para algunos tipos celulares; formación de nanonets; efecto adyuvante en la activación de APC y maduración de DC y una débil actividad antimicrobiana. Se ha demostrado que la desregulación en los niveles de HDP es característica de varias enfermedades17. También se ha observado la influencia de los HDP naturales en el crecimiento tumoral. Los datos sugieren que muchas defensinas son protumorales aunque, por el contrario, podrían reflejar una aberrante respuesta del huésped. En consistencia con la promoción tumoral, las defensinas promueven el crecimiento celular y la proliferación in vitro e influyen en el microam- biente del tumor mediante la promoción de la angiogénesis y la señalización antiapoptótica21. Además, la producción alterada de HDP por las células epiteliales de las vías respiratorias parece contribuir a varias enfermedades pulmonares, incluyendo la fibrosis quística, la enfermedad pulmonar obstructiva crónica y el asma22. Asimismo, la desregulación de HDP es un factor que contribuye a la patogenia de varias enfermedades autoinmunes como la diabetes tipo 1, la artritis reumatoide, el lupus eritematoso sistémico y la psoriasis. Se cree que esto es principalmente debido a una respuesta proinflamatoria que se activa por HDP, particularmente la producción de interferones tipo I23. Finalmente, como muchos HDP se producen continuamente por la piel sana, es comprensible pensar que algunas condiciones patológicas, tales como la dermatitis atópica y la psoriasis, estén asociadas a una desregulación de las HDP producidas por células de la piel. El complemento El sistema del complemento se compone de cerca de 20 proteínas distintas que trabajan juntas para marcar y destruir invasores, así como para avisar y reclutar a otros «actores» del sistema inmune, indicándoles que el ataque ha empezado. El sistema del complemento es muy antiguo, incluso los erizos de mar, que se desarrollaron hace unos 700 millones años, tienen un sistema de complemento. En seres humanos, las proteínas del complemento comienzan a sintetizarse durante el primer trimestre del desarrollo fetal, por lo que queda patente la importancia de este sistema que debe estar listo antes de que nazca el feto. De hecho, pese a su baja incidencia, los escasos seres humanos nacidos con un defecto en una de las proteínas importantes del complemento generalmente no viven mucho tiempo antes de sucumbir a la infección. En la activación del complemento participan cascadas proteolíticas en las que se hidrolizan zimógenos inactivos que pasan a ser moléculas activas con varias funciones. Esto confiere al sistema una tremenda capacidad de amplificación por la cantidad de los productos proteolíticos que se generan. La activación del complemento requiere el reconocimiento de moléculas expresadas en las superficies microbianas, pero que no están presentes en las células propias, y esto puede ocurrir de tres formas distintas que dan nombre a las tres vías de activación del sistema24. La vía clásica depende de anticuerpos para su activación. La proteína plasmática que reconoce la porción Fc de los anticuerpos unidos a la superficie de un microbio u otra estructura es C1q25. Una vez que se produce este reconocimiento se inicia una cascada proteolítica que afecta a otras proteínas del complemento como veremos más adelante. Esta vía es un ejemplo de la tenue barrera que existe entre la respuesta inmune innata y la adaptativa. La segunda vía de activación del sistema del complemento se denomina vía alternativa aunque, en términos evolutivos, la vía alternativa ciertamente evolucionó antes que la vía clásica. Las proteínas que componen el sistema del complemento son producidas principalmente por el hígado y están presentes en altas concentraciones en sangre y tejidos. La más abundante es C3 que se divide continuamente en dos Medicine. 2017;12(24):1388-97 1395 ENFERMEDADES DEL SISTEMA INMUNE Y REUMATOLÓGICAS (I) proteínas más pequeñas. Una de estas proteínas, C3b, es altamente reactiva y se une tanto a superficies de células propias como de microbios. Sin embargo, en las superficies celulares propias se inhibe por la acción de moléculas reguladoras presentes en las células de los mamíferos. Como los microbios carecen de estas proteínas reguladoras, la activación espontánea puede amplificarse en las superficies microbianas. Es, por tanto, un sistema en el que la opción por defecto es la muerte26. La tercera forma de activar el complemento es la vía de las lectinas y puede ser la más importante vía de activación de todas. La proteína clave de esta vía se produce principalmente en el hígado y está presente en concentraciones moderadas en la sangre y los tejidos. Esta proteína se llama lectina de unión a manosa (MBL —mannose-binding lectin—). La manosa es una molécula de hidratos de carbono que se encuentra en la superficie de muchos patógenos comunes27. Por ejemplo, la MBL se une a hongos como Candida albicans; a virus como el VIH-1 y el de la gripe A; a muchas bacterias como Salmonella y Streptococcus y a parásitos como Leishmania. Por el contrario, la MBL no se une a hidratos de carbono presentes en tejidos y células humanas sanas. La activación del complemento por cualquiera de las tres vías conduce al reclutamiento y ensamblaje de otras proteínas del complemento, siendo la más importante la C3-convertasa que hidroliza la proteína central del sistema del complemento, C3, y genera C3a y C3b. C3b sirve de opsonina que promueve la fagocitosis de los microbios y C3a estimula la inflamación al actuar como sustancia quimioatrayente para los neutrófilos. Además, C3b une otras proteínas del complemento que acaban formando el complejo de ataque a membrana (MAC —membrane attack complex—) que causa la lisis del microbio en el que se activa el complemento28. Resumiendo, el sistema del complemento es multifuncional. Puede destruir invasores mediante la construcción de MAC, puede mejorar la función de células fagocíticas a través de sus receptores de complemento y puede alertar a otras células de que estamos siendo atacados. Y lo más importante es que puede hacer todas estas cosas muy rápido. Pentraxinas Las pentraxinas son el grupo de proteínas pentaméricas con homología estructural más antiguo en la filogenia. Son un tipo de PRR y están implicadas en las respuestas de fase aguda junto con otras proteínas plasmáticas cuya síntesis por el hígado se incrementa en procesos inflamatorios y que se denominan reactantes de fase aguda29. Los miembros más importantes de esta familia son las pentraxinas cortas como la proteína C reactiva (PCR) y el amiloide sérico P (SAP), y la pentraxina larga PTX330. La PCR y el SAP se unen a diferentes especies de bacterias y hongos. La PCR une con alta afinidad residuos de fosfocolina presentes en los hidratos de carbono expresados en virus, bacterias, hongos y parásitos y en células apoptóticas propias. Por lo tanto, la PCR es capaz de reconocer no solo microorganismos, sino también células propias que deben ser eliminadas. La principal función de la PCR es reconocer patóge1396 Medicine. 2017;12(24):1388-97 nos y células dañadas y mediar su eliminación induciendo la activación del complemento y la fagocitosis. El SAP reconoce la fosfatidiletanolamina y tiene funciones similares. Además, PCR, SAP y PTX3 activan el complemento al unirse a C1q e inician la vía clásica. Las concentraciones plasmáticas de PCR son muy bajas en sujetos sanos; sin embargo, pueden aumentar hasta mil veces durante las infecciones o en respuesta a otros estímulos inflamatorios. La IL-6 y, en menor medida, la IL-1 y el TNF-D, producidos mayoritariamente por los macrófagos en el foco infeccioso, estimulan la síntesis de la PCR por los hepatocitos, lo que la convierte, por tanto, en un buen indicador de inflamación sistémica. Colectinas y ficolinas y otras moléculas solubles Las colectinas son una familia de proteínas que funcionalmente son trímeros o hexámeros cuya subunidad monomérica consta de cuatro partes, de las cuales dos son esenciales: un dominio tipo colágeno y un dominio de lectina tipo C cuya unión depende de Ca2+. Los tres miembros más importantes de esta familia son la MBL, que vimos en el capítulo del complemento, y las proteínas del surfactante pulmonar SP-A y SP-D31. La proteína A surfactante (SP-A) y la proteína D surfactante (SP-D) se encuentran en los alvéolos pulmonares y, aparte de ayudar a reducir la tensión superficial alveolar, median respuestas inmunitarias innatas en el pulmón. Se unen a varios microorganismos y actúan como opsoninas facilitando su fagocitosis por los macrófagos alveolares. SP-A y SP-D también pueden inhibir directamente el crecimiento bacteriano y pueden activar a los macrófagos. Por último, actúan como agente protector del surfactante pulmonar frente a las proteasas microbianas. Las ficolinas son proteínas séricas que presentan una estructura similar a las colectinas, salvo los dominios de reconocimiento de los hidratos de carbono que son dominios de tipo fibrinógeno32. Las ficolinas reconocen grupos acetilo presentes en los hidratos de carbono, principalmente N-acetil glucosamina y ácido lipoteicoico. Sus funciones son muy similares a la MBL. Por último, existen otras moléculas solubles que pertenecen a la rama humoral de la respuesta innata. La fosfolipasa A2 se encuentra en los gránulos de los neutrófilos y es producida por células epiteliales. Su función es matar directamente bacterias Gram positivas hidrolizando fosfolípidos de la membrana microbiana33. La lactoferrina pertenece a la familia de la transferrina y actúa secuestrando hierro, lo que limita su disponibilidad para las bacterias34. La lisozima es una enzima abundante en numerosas secreciones como la saliva, las lágrimas y el moco y también en los gránulos de los neutrófilos. Su función es catalizar la hidrólisis de las uniones beta 1,4 entre los residuos de ácido N-acetilmurámico y N-acetil-D-glucosamina del peptidoglicano35. Conflicto de intereses Los autores declaran no tener ningún conflicto de intereses. RESPUESTA INMUNE INNATA Y SUS IMPLICACIONES FISIOPATOLÓGICAS Responsabilidades éticas Protección de personas y animales. Los autores declaran que para esta investigación no se han realizado experimentos en seres humanos ni en animales. Confidencialidad de los datos. Los autores declaran que en este artículo no aparecen datos de pacientes. Derecho a la privacidad y consentimiento informado. Los autores declaran que en este artículo no aparecen datos de pacientes. Bibliografía • Importante •• Muy importante ✔ Metaanálisis ✔ Artículo de revisión Ensayo clínico controlado ✔ ✔ Guía de práctica clínica Epidemiología ✔ 1. • Medzhitov R, Janeway CA Jr. Decoding the patterns of self and ✔ nonself by the innate immune system. Science. 2002;296(5566):298300. 2. Kumar H, Kawai T, Akira S. Pathogen recognition by the innate immune ✔ system. Int Rev Immunol. 2011;30(1):16-34. 3. • Janeway C A, Jr. Approaching the asymptote? Evolution and revo✔ lution in immunology. Cold Spring Harb Symp Quant Biol. 1989;54:1-13. 4. •• Brubaker SW, Bonham KS, Zanoni I, Kagan JC. Innate immune ✔ pattern recognition: a cell biological perspective. Annu Rev Immunol. 2015;33:257-90. 5. Deretic V, Saitoh T, Akira S. Autophagy in infection, inflammation and ✔ immunity. Nat Rev Immunol. 2013;13(10):722-37. 6. Lamkanfi M, Dixit VM. Mechanisms and functions of inflammasomes. ✔ Cell. 2014;157(5):1013-22. 7. •• Broz P, Monack DM. Newly described pattern recognition re✔ ceptors team up against intracellular pathogens. Nat Rev Immunol. 2013;13(8):551-65. 8. Bianchi ME. DAMPs, PAMPs and alarmins: all we need to know about ✔ danger. J Leukoc Biol. 2007;81(1):1-5. 9. Osorio F, Reis E, Sousa C. Myeloid C-type lectin receptors in pathogen ✔ recognition and host defence. Immunity. 2011;651-64. 10. Brown GD. Dectin-1: a signalling non-TLR pattern-recognition recep✔ tor. Nat Rev Immunol. 2006;6:33-43. 11. Elinav E, Trowig T, Henao-Mejia J, Flavell RA. Regulation of the antimicrobial response by NLR proteins. Immunity. 2011;34:665-79. 12. Philpott DJ, Sorbara MT, Robertson SJ, Croitoru K, Girardin SE. NOD proteins: regulators of inflammation in health and disease. Nat Rev Immunol. 2014;14(1):9-23. ✔• 13. • Tschopp J, Martinon F, Burns K. NALPs: a novel protein family ✔ involved in inflammation. Nat Rev Mol Cell Biol. 2003;4(2):95-104. 14. Yoneyama M, Kikuchi M, Matsumoto K, Imaizumi T, Miyagishi M, Taira K, et al. Shared and unique functions of the DExD/H-box helicases RIGI, MDA5, and LGP2 in antiviral innate immunity. J Immunol. 2005;175(5):2851-8. 15. Unterholzner L. The interferon response to intracellular DNA: why so many receptors? Immunobiology. 2013;218(11):1312-21. 16. Canton J, Neculai D, Grinstein S. Scavenger receptors in homeostasis and immunity. Nat Rev Immunol. 2013;13(9):621-34. 17. Hancock RE, Haney EF, Gill EE. The immunology of host defence peptides: beyond antimicrobial activity. Nat Rev Immunol. 2016;16(5):321-34. 18. Nijnik A, Hancock REW. The roles of cathelicidin LL-37 in immune defences and novel clinical applications. Curr Opin Hematol. 2009;16:417. 19. Hazlett L, Wu M. Defensins in innate immunity. Cell Tissue Res. 2011; 343:175-88. 20. Semple F, Dorin JR. E-Defensins: multifunctional modulators of infection, inflammation and more? J Innate Immun. 2012;4:337-48. 21. Winter J, Pantelis A, Reich R, Martini M, Kraus D, Jepsen S, et al. Human E-defensin-1, –2, and –3 exhibit opposite effects on oral squamous cell carcinoma cell proliferation. Cancer Invest. 2011;29:196-201. 22. Lecaille F, Lalmanach G, Andrault PM. Antimicrobial proteins and peptides in human lung diseases: a friend and foe partnership with host proteases. Biochimie. 2016;122:151-68. 23. Kahlenberg J M, Kaplan MJ. Little peptide, big effects: the role of LL-37 in inflammation and autoimmune disease. J Immunol. 2013;191:4895901. 24. Holers VM. Complement and its receptors: new insights into human disease. Annu Rev Immunol. 2014;32:433-59. 25. Lachmann PJ, Hughes-Jones NC. Initiation of complement activation. Springer Semin Immunopathol. 1984;7:143-62. 26. Muller-Eberhard HJ. Molecular organization and function of the complement system. Annu Rev Biochem. 1988;57:321-47. 27 Reid KBM, Turner MW. Mammalian lectins in activation and clearance mechanisms involving the complement system. Springer Semin Immunopathol. 1994;15:307-25. 28. Wetsel RA. Structure, function and cellular expression of complement anaphylatoxin receptors. Curr Opin Immunol. 1995;7:48-53. 29. Christiaans SC, Wagener BM, Esmon CT, Pittet JF. Protein C and acute inflammation: a clinical and biological perspective. Am J Physiol Lung Cell Mol Physiol. 2013;305(7):L455-66. 30. Bottazzi B, Inforzato A, Messa M, Barbagallo M, Magrini E, Garlanda C, et al. The pentraxins PTX3 and SAP in innate immunity, regulation of inflammation and tissue remodelling. J Hepatol. 2016; 64(6):1416-27. 31. Foo SS, Reading PC, Jaillon S, Mantovani A, Mahalingam S. Pentraxins and collectins: friend or foe during pathogen invasion? Trends Microbiol. 2015;23(12):799-811. 32. Endo Y, Matsushita M, Fujita T. Role of ficolin in innate immunity and its molecular basis. Immunobiology. 2007;212(4-5):371-9. 33. Laine VJ, Grass DS, Nevalainen TJ. Protection by group II phospholipase A2 against Staphylococcus aureus. J Immunol. 1999;162(12): 7402-8. 34. Hoek KS, Milne JM, Grieve PA, Dionysius DA, Smith R. Antibacterial activity in bovine lactoferrin-derived peptides. Antimicrob Agents Chemother. 1997;41(1):54-9. 35. McKenzie HA, White FH. Lysozyme and alpha-lactalbumin: structure, function, and interrelationships. Adv. Protein Chem. 1991;41:173-315. ✔• ✔ •• ✔ ✔ ✔• ✔ ✔• ✔ ✔ Medicine. 2017;12(24):1388-97 1397