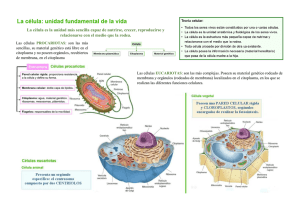
CITOPLASMA CELULAR GENERALIDADES DE LA CÉLULA Y EL CITOPLASMA Las células: son las unidades estructurales y funcionales básicas de todos los organismos multicelulares. Los procesos que asociamos con las actividades de los organismos son reflejos de procesos similares que ocurren dentro de cada una de las células que constituyen el cuerpo humano. Algunas células desarrollan una o más de estas funciones en un grado tal de especialización que son identificadas por la función y las estructuras celulares asociadas con ellas. La actividad o función especializada de una célula puede ser un reflejo no solo de la presencia de una gran cantidad del componente estructural específico que realiza la actividad, sino también de la forma de la célula, su organización con respecto a otras células similares, y sus productos. Las células pueden dividirse en dos compartimientos principales: Citoplasma: región de la célula localizada fuera del núcleo. Contiene orgánulos y citoesqueleto, formado por proteínas polimerizadas que forman microtubulos, filamentos intermedios y filamentos de actina, e inclusiones suspendidas en un gel acuoso denominado matriz citoplasmática Núcleo: orgánulo más grande dentro de la célula y contiene el genoma junto con las enzimas necesarias para la replicación de ADN y la transcripción de ARN. Matriz: solutos que incluye iones inorgánicos (Na +, K+, Ca+2) y moléculas orgánicas como Metabolitos intermedios, hidratos de carbono, lípidos, proteínas y ARN. Las células tienen el mismo conjunto básico de orgánulos, los que pueden clasificarse en dos grupos: 1) Orgánulos membranosos: con membrana plasmática que separan el ambiente interno del orgánulo del citoplasma. 2) Orgánulos no membranosos: carentes de membrana plasmática. Los orgánulos membranosos: Membrana plasmática: una bicapa lipídica que forma el límite de la célula como los limites de orgánulos dentro de la célula. Retículo endoplásmico rugoso (RER): una región del retículo endoplásmico asociada con ribosomás en donde se sintetizan y modifican proteínas. Retículo endoplásmico liso (REL): una región del retículo endoplásmico carente de ribosomas, involucrada en la síntesis de lípidos y esteroides. Aparato de Golgi: un orgánulo membranosos compuesto por cisternas aplanadas responsables de la modificación, clasificación y empaquetamiento de pretinas y lípidos para su transporte intra-celular o extra-celular. Endosomas: compartimientos limitados por membrana que participan en los mecanismos de endocitosis, clasifica las proteínas que le son enviadas por las vesículas endocíticas, y redirigirlas a diferentes compartimientos celulares que serán sus destinos finales. Lisosomas: orgánulos con enzimas digestivas que se forman a partir de vesículas que se desprenden del aparato de Golgi. Vesículas de transporte: están involucradas en la endocitosis y la exocitosis Mitocondria: orgánulo que proporciona la energía a la célula al producir adenosina trifosfato (ATP) Peroxisoma: orgánulo involucrado en la producción y degradación de H2O2 y en la degradación de ácidos grasos Los orgánulos no membranosos: Microtubulos: forman elementos del citoesqueleto y constantemente se alargan y se acortan. Filamentos: son parte del citoesqueleto y pueden clasificarse en dos grupos: filamentos de actina y filamentos intermedios. Ambos grupos proporcionan resistencia a la tracción para soportar la tensión. Centriolos: par de estructuras cilíndricas cortas que se encuentran en el centro de organización de microtubulos o centrosoma. Ribosomas: estructuras para la síntesis de proteínas compuestas por ARN ribosoma (ARNr) y proteínas ribosomales. Proteasomas: complejos de proteínas que degradan proteínas dañadas o innecesarias en polipéptidos pequeños y aminoácidos. MEMBRANA PLASMÁTICA Estructura dinámica que participa en forma activa en muchas actividades fisiológicas y bioquímicas esenciales para el funcionamiento y la supervivencia de la célula. El espesor total de la membrana plasmática es alrededor de 8 a 10nm. Organización molecular de la membrana plasmática consiste en el llamado modelo de mosaico fluido modificado Compuesta por moléculas de fosfolípidos, colesterol, y proteínas. Las moléculas de lípido forman una bicapa lipídica de carácter anfipático (tanto hidrófoba como hidrófila). Las cadenas de ácidos grasos de las moléculas lipídicas se enfrentan entre sí, tornando hidrófoba (es decir, que no tiene afinidad por el agua) la porción interna de la membrana. Las superficies de la membrana están formadas por los grupos polares de las cabezas de las moléculas lipídicas, haciendo de este modo que las superficies se tornen hidrófilas (es decir, con afinidad por el agua) Balsas lipídicas: dominios focalizados contienen altas concentraciones de colesterol y glucoesfingolípido. Debido a la alta concentración de colesterol y a la presencia de cadenas largas de ácidos grasos altamente saturados, la superficie de la balsa lipídica es más gruesa y muestra una menor fluidez que la membrana plasmática circundante Balsas lipídicas planas: contienen una familia de proteínas de 47 kDa conocidas como flotilinas, además de la composición específica de lípidos y colesterol Balsas caveolares, o caveolas: representan pequeñas invaginaciones de la membrana plasmática en forma de botella (50 a 100nm de diámetro), enriquecidas con pequeñas proteínas integrales de membrana (18 a 24kDa) Proteínas integrales de membrana Las bombas: sirven para transportar iones, como el Na+ , a través de las membranas. También transporten precursores metabólicos de macromoléculas, como los aminoácidos y monosacáridos. Los conductos: permiten el paso de pequeños iones, moléculas y agua a través de la membrana plasmática en cualquier dirección (mediante difusión pasiva). Permiten el paso de iones y moléculas pequeñas implicadas en los procesos de señalización, desde el citoplasma de una de las células al citoplasma de células contiguas. Proteínas receptoras: permiten el reconocimiento y la unión especifica de ligandos (moléculas que se unen a la superficie extra-celular de la membrana plasmática) en procesos como la estimulación hormonal, endocitosis de vesículas con cubierta, y reacciones con anticuerpos. Los receptores que se unen a moléculas de señalización transmiten la señal a través de una secuencia de interruptores moleculares (segundos mensajeros) a los mecanismos de señalización internos, iniciando así una repuesta fisiológica. Proteínas de enlace: fijan el citoesqueleto intra-celular a la matriz extra-celular. Enzimas: desempeñan papeles específicos en el bombeo de iones: la ATP sintetasa y las enzimas digestivas como di-sacaridasas y di-peptidasas. Proteínas estructurale:s forman uniones con células vecinas. Procesos de señalización: participan de la regulación de la expresión de genes, la exocitosis, la endocitosis, la diferenciación, el crecimiento y la muerte celular, la re-organización del citoesqueleto, el movimiento, la contracción y/o la relajación celular. Señalización celular: proceso por el cual las células reciben, procesan y transmiten los estímulos extra-celulares para regular sus propias respuestas fisiológicas. Vías de transducción de señales: son mecanismos mediante los cuales las células responden al ambiente externo. Son cascadas de eventos moleculares que median la especificidad celular y tisular, permitiendo la amplificación y la modulación de la señal y están involucradas en la regulación bioquímica y fisiológica. Se inducen por moléculas de señalización externa (también conocidas como mensajeros primarios o ligandos). Las señales que se originan desde los receptores son transmitidas a moléculas diana dentro de la celul por el sistema de segundos mensajeros. Transporte de membrana y transporte vesicular Algunas sustancias (moléculas lipo-solubles y moléculas pequeñas sin carga) cruzan la membrana plasmática por difusión simple a favor de su gradiente de concentración. Las otras moléculas necesitan proteínas de transporte de membrana que les proporcionen un pasaje individual a través de la membrana plasmática: Proteínas transportadoras que transfieren moléculas hidro-solubles pequeñas. Son altamente selectivas, con frecuencia solo transportan un tipo de molécula. Algunas proteínas transportadoras, como la bomba de Na+ /K+ o la bomba de H+, requieren energía para el transporte activo de moléculas en contra de su gradiente de concentración. Otras transportadoras no requieren energía y participan en el transporte pasivo. Proteínas canal: transfieren moléculas hidrosolubles pequeñas. Los conductos están formados por proteínas transmembrana, las proteínas de canal contienen un dominio poro que penetra parcialmente la bicapa de la membrana y sirve como filtro selectivo de iones. Se regula mediante los potenciales de membrana, neurotransmisores o estrés mecánico. Transporte vesicular mantiene la integridad de la membrana plasmática y contribuye a la transferencia de moléculas entre los diferentes compartimientos celulares. Proceso que implica cambios de configuración en la membrana plasmática en sitios localizados y la consecuente formación de vesículas a partir de la membrana o fusión de vesículas con ella: Endocitosis: es el transporte vesicular en los cuales las sustancias ingresan a la célula. Cumple funciones clave en la incorporación de nutrientes, señalización celular y cambios en la forma celular Exocitosis: es el transporte vesicular en los cuales las sustancias abandonan la célula. ENDOCITOSIS Pinocitosis: es la ingestión inespecífica de líquido y pequeñas moléculas de proteína mediante vesículas pequeñas, es constitutivo, implica una formación dinámica continua de vesículas pequeñas en la superficie celular. Fagocitosis: incorporación de partículas grandes como detritos celulares, bacterias y otros materiales extraños. La membrana plasmática emite seudópodos que rodean las partículas a fagocitar formando vesículas grandes, denominadas fagosomas. Mediado por receptores en la superficie celular reconocen el dominio (región del anticuerpo que no se une al antígeno) de los anticuerpos que revisten la superficie de un microorganismo invasor o de una célula invasora. Endocitosis mediada por receptor: permite la entrada de moléculas específicas en la célula. Los receptores para moléculas específicas, receptores de carga, se acumulan en regiones bien definidas de la membrana celular. Se convierten en fositas recubiertas. Los receptores de carga reconocen y unen moléculas específicas que entran en contacto con la membrana plasmática. Las moléculas de clatrina se agrupan para armar una jaula, similar a un cesto, que ayuda a cambiar la forma de la membrana plasmática en una invaginación de tipo vesícula. EXOCITOSIS Proceso mediante el cual una vesícula se desplaza desde el citoplasma hacia la membrana plasmática, donde descargo su contenido al espacio extra-celular. se logra mediante la presencia de proteínas específicas en su superficie. Las moléculas que viajan por esta ruta sufren modificaciones químicas (glicosilación, sulfatación) a medida que atraviesan diferentes compartimentos celulares Vía constitutiva: las sustancias se envían en forma continua hacia le membrana plasmática en las vesículas de transporte. Las proteínas se secretan en forma inmediata después de su síntesis y salen del aparato de Golgi. Vía de secreción regulada: las células endocrinas, exocrinas y las neuronas, concentran proteínas de secreción y las almacenan en vesículas secretoras dentro del citoplasma. Para que se produzca la secreción debe activarse un fenómeno regulador (estimulo hormonal o nervioso). Los estímulos de señalización causan la entrada transitoria de Ca+2 en el citoplasma, estimula las vesículas secretoras para que se fusionen con la membrana plasmática y descarguen su contenido. ENDOSOMAS Endosomas tempranos: compartimientos limitados por membrana relacionados con las vías endocíticas, restringen a una porción del citoplasma cerca de la membrana celular donde se fusionan las vesículas que se originan de la membrana celular. Función: es clasificar y reciclar proteínas interiorizadas por vías endocíticas. Después de la clasificación, las proteínas se reciclan y el exceso de membrana se devuelve a la membrana plasmática Endosomas tardíos: Su pH es más acido, con un promedio de 5,5, suelen posicionarse cerca del aparato de Golgi y del núcleo, maduran hasta convertirse en lisosomas. Modelo de compartimento estable: describe a las endosomas tempranos y tardíos como orgánulos celulares estables conectados mediante el transporte vesicular con el ambiente externo de la célula y con el aparato de Golgi. Modelo madurativo: la composición de la membrana endosómica temprana cambia en forma progresiva a medida que algunos compo- nentes son reciclados entre la superficie celular y el aparato de Golgi. Este proceso conduce a la formación de endosomas tardíos LISOSOMAS Lisosomas: orgánulos ricos en enzimas hidrolíticas, como proteasas, nucleasas, glucosidasas, lipasas y fosfolipasas. Degrada macromoléculas derivadas de los mecanismos endocíticos, como de la celula, autofagia. Se forman en una serie de mecanismos que convergen en los endosomas tardíos, transformándolos en lisosomas. Contienen una colección de enzimas hidrolíticas y son rodeadas por una sola membrana que resiste la hidrólisis mediante sus propias enzimas. Contienen bombas de protones (H+ ) que transportan iones H+ a la luz lisosómica, manteniendo un pH bajo (4,7). Membrana lisosómica contiene proteínas transportadoras que transportan productos finales de la digestión (aminoácidos, sacáridos, nucleótidos) hacia el citoplasma, donde se utilizan en los procesos sintéticos de la célula o sufren exocitosis. Tres mecanismos de la digestión intra-celula Partículas extra-celulares grandes: como bacterias, detritos celulares y otros materiales extraños son engullidos en el proceso de fagocitosis. Un fagosoma, formado a medida que el material se incorpora dentro del citoplasma, recibe enzimas hidrolíticas para convertirse en un endosoma tardío, el que madura hasta convertirse en un lisosoma. Partículas extra-celulares pequeñas: como proteínas extra-celulares, proteínas de la membra na plasmática y complejos ligando-receptor se incorporan por pinocitosis y endocitosis mediada por receptores Partículas intracelulares: como orgánulos enteros, proteínas citoplasmáticas y otros componentes celulares son aislado de la matriz citoplasmática por las membranas del retículo endoplásmico, transportadas hacia los lisosomas y degradadas. AUTOGAFIA Autofagia: mecanismo celular por el cual varias proteínas citoplasmáticas, orgánulos y otras estructuras celulares son degradadas en el compartimiento lisosómico. Macroautofagia: proceso en el cual una porción del citoplasma o un orgánulo completo es rodeado por una membrana intra-celular doble o multi-laminar del retículo endoplásmico, membrana de aislamiento para formar una vacuola autofagosoma. Después de la entrega dirigida de enzimas lisosómicas, el autofagosoma madura para convertirse en un lisosoma. Microautofagia: es un proceso en el cual las proteínas citoplasmáticas son degradadas en un proceso lento y continuo en condiciones fisiológicas normales. Autofagia mediada por chaperonas: proceso selectivo de degradación proteica y requiere la colaboración de chaperonas citosólicas específicas. Se activa durante la privación de sustancias nutritivas y necesita la presencia de señales de localización en las proteínas que se han de degradar y de un receptor especifico en la membrana lisosómica. DEGRADACIÓN MEDIADA POR PROTEASOMAS Capacidad de destruir proteínas sin la participación de los lisosomas, ocurre dentro de las proteasomas. Se utilizan para destruir proteínas anómalas que están mal plegadas o desnaturalizadas o que contienen aminoácidos anómalos, degrada proteínas reguladoras normales de vida corta que necesitan ser inactivadas con rapidez, como las ciclinas mitóticas que regulan la progresión del ciclo celular Proteasomas: son complejos de proteasa dependientes de ATP que destruyen proteínas que han sido marcadas específicamente para este proceso. RETÍCULO ENDOPLÁSMICO RUGOSO Ergastoplasma: porción del citoplasma que se tiñe con un colorante básico. El citoplasma de células que participan en la síntesis proteica, se tiñe en forma intensa causada por la presencia de ARN El RER: serie de sacos membranosos aplanados e interconectados denominados cisternas, con ribosomas, que están adheridas a la membrana del RER por proteínas de acoplamiento ribosómico. Los ribosomas se componen de una sub-unidad menor y una sub-unidad mayor. Cada sub-unidad contiene ARN ribosómico (ARNR). El RER es continuo con la membrana externa de la envoltura nuclear. Los grupos de ribosomas forman poliribosomas o polisomas en los que muchos ribosomas están adosados a una hebra de ARN mensajero (ARNM). El RER se encuentra muy bien desarrollado en células secretoras activas, que sintetizan proteínas destinadas a abandonar la célula (células secretoras) y en las células con gran cantidad de membrana plasmática, como las neuronas. Ribosomas libres: sintetizan proteínas que permanecerán en la célula como elementos citoplasmáticos estructurales o funcionales. Las proteínas destinadas al núcleo, la mitocondria o los peroxisomas se sintetizan en estos y después se liberan en el citosol. RETÍCULO ENDOPLÁSMICO LISO El retículo endoplásmico liso: compuesto por túbulos cortos anastomosados que no están asociados con los ribosomas, carece de proteínas de acoplamiento ribosómico. Tiende a ser tubular en lugar de sacular, y puede estar separado del RER o ser una extensión de él. Es abundante en células que participan en el metabolismo de los lípidos. Está bien desarrollado en células que sintetizan y secretan esteroides, como las de la corteza supra-renal y las testiculares de Leydig (intersticiales). Este retículo secuestra el Ca+2 que es esencial para el proceso de contracción y está en estrecho contacto con las invaginaciones de la membrana plasmática que conducen los impulsos contráctiles al interior de la célula. El REL interviene en la desintoxicación y en la conjugación de sustancias nocivas. Modifican y desintoxican compuestos hidrófobos, como pesticidas y carcinógenos, convirtiéndolos en productos conjugados hidrosolubles que pueden ser eliminados del organismo. Participa en: Metabolismo de los lípidos, esteroides, glucógeno, formación y el reciclaje de membranas. APARATO DE GOLGI Está bien desarrollado en las células secretoras y no se tiñe con hematoxilina o eosina. Se encuentra en relación funcional con el RER, aparece como una serie apilada (rimeros) de sacos o cisternas de membranas aplanadas y extensiones tubulares incluidas en una red de microtubulos cerca del centro organizador de los microtubulos. Se observan vesículas pequeñas que participan en el transporte vesicular. Las cisternas aplanadas localizadas más cerca del RER constituyen la cara formadora o red cis-Golgi (CGN); las cisternas ubicadas mas lejos del RER representan la cara madurativa o red trans-Golgi (TGN). Las cisternas ubicadas entre la TNG y la CGN suelen denominarse red intermedia del Golgi. El aparato de Golgi participa en la modificación pos-traduccional, en la clasificación y en el envasado de las proteínas. MITOCONDRIA Las mitocondrias: son abundantes en las células que generan y gastan gran cantidad de energía, como las células musculares estriadas y las células involucradas en el transporte de líquidos y electrolitos. Cuando se presentan en grandes cantidades las mitocondrias contribuyen a la acidofilia del citoplasma. Las mitocondrias poseen dos membranas que delinean compartimientos bien definidos. Membrana mitocondrial interna: rodea la matriz. Membrana mitocondrial externa: está en estrecho contacto con el citoplasma. El espacio entre las dos membranas, espacio intermembrana. Los componentes estructurales de las mitocondrias son: Membrana mitocondrial externa: esta membrana lisa, contiene muchos conductos aniónicos dependientes de voltaje. Son permeables a moléculas sin carga. Las pequeñas moléculas, iones y Metabolitos pueden entrar en el espacio intermembrana pero no pueden penetrar la membrana interna, posee receptores para las proteínas y los polipéptidos que se translocan en el espacio intermembrana. Membrana mitocondrial interna: es más delgada que la membrana mitocondrial externa. Esta organizada en numerosas crestas que incrementan en forma el área de superficie de la membrana interna. Estos pliegues se proyectan hacia la matriz que compone el compartimiento interno del orgánulo. La membrana que forman las crestas contiene proteínas que cumplen tres funciones: llevar a cabo las reacciones de oxidación de la cadena respiratoria de transporte de electrones, sintetizar ATP y regular el transporte de metabolitos hacia dentro y hacia fuera de la matriz. Las enzimas para sintetizar ATP están unidas a la membrana interna y proyectan componentes hacia la matriz. Espacio intermembrana: ubican entre las membranas interna y externa y contiene enzimas específicas que utilizan el ATP generado en la membrana interna. Matriz: la matriz mitocondrial está rodeada por la membrana mitocondrial interna y contiene las enzimas solubles del ciclo del acido cítrico (ciclo de Krebs) y las enzimas involucradas en la beta-oxidación de los ácidos grasos. Los productos principales de la matriz son CO2 y NADH reducido, que es la fuente de electrones para la cadena de transporte electrónico. PEROXISOMAS Peroxisomas: orgánulos limitados por membrana que contiene enzimas oxidatívas. Son esféricos y pequeños, contienen enzimas oxidatívas, en particular catalasa y otras peroxidasas. Prácticamente todas las enzimas oxidatívas producen peróxido de hidrogeno (H2O2) como un producto de la reacción oxidativa. Esta una sustancia toxica. Los peroxisomas, regulan con precisión el contenido celular de peróxido de hidrogeno y lo degrada para proteger la célula. ORGÁNULOS NO MEMBRANO MICROTÚBULOS Los microtúbulos: son tubos huecos, rígidos y no ramificados de proteínas polimerizadas que pueden armarse y desarmarse con la misma rapidez. Se encuentran en el citoplasma, originan a partir del centro organizador de microtúbulos (MTOC). Están presentes en los cilios y en los flagelos, en los centriolos y el huso mitótico; y en los procesos de elongación de la célula. Intervienen en muchas funciones celulares: Transporte vesicular intracelular. Crean un sistema de conexiones dentro de la célula. Movimiento de los cilios y flagelos. Unión de los cromosomas con el huso mitótico y sus movimientos durante la mitosis y la meiosis Elongación y desplazamiento celular (migración). Mantenimiento de la forma celular. Son estructuras poliméricas alargadas compuestas por partes iguales de tubulina alfa y tubulina beta. Consiste en 13 moléculas globulares diméricas de la proteína tubulina dispuestos en forma circular. Los dímeros se polimerizan extremo con extremo y cabeza con cola; la molécula alfa de un dímero se une con la molécula beta del siguiente dímero en un patrón de repetición. Las proteínas moleculares motoras: se adhieren a estos orgánulos o estructuras y los arrastran a lo largo de las guías micro-tubulares. Dos familias de proteínas moleculares motoras permiten el desplazamiento unidi- reccional: Las dineínas: constituyen una familia de motores moleculares. Se desplazan sobre los microtúbulos hacia su extremo minus (-); son capaces de transportar orgánulos desde la periferia celular hacia el MTOC. Las cinesinas: se desplazan sobre los microtubulos hacia su extremo plus (+); son capaces de desplazar orgánulos desde el centro celular hacia la periferia de la célula. FILAMENTOS DE ACTINA Los filamentos de actina: Se arman en forma espontanea por polimerización en una estructura lineal helicoidal para formar filamentos. Son más finos, cortos y flexibles que los microtubulos. Moléculas de actina libres en el citoplasma: se denominan actina G (actina globular), la actina polimerizada se denomina actina F (actina filamentosa). Un filamento de actina o micro-filamento es una estructura polarizada; su extremo de crecimiento rápido se denomina extremo plus y su extremo de crecimiento lento se denomina extremo minus. Existen dos tipos de filamentos en las células musculares: los filamentos de actina o filamentos finos y los filamentos de miosina II o filamentos gruesos. Las funciones incluyen: Anclaje y movimiento de proteínas de la membrana: se distribuyen en redes tridimensionales por toda la célula y se utilizan como estructuras de anclaje dentro de las uniones celulares especializadas. Formación del núcleo estructural de las micro-vellosidades en las células epiteliales absortivas: contribuirían a mantener la forma de la superficie celular apical. Locomoción celular: se alcanza por la fuerza ejercida por los filamentos de actina al polimerizarse a la altura de sus extremos de crecimiento. Como consecuencia las células extienden evaginaciones desde sus superficies al empujar la membrana plasmática por delante de los filamentos de actina en crecimiento. Emisión de evaginaciones celulares: puede observarse en células que exhiben pequeñas protrusiones denominadas filopodios, ubicados alrededor de su superficie. FILAMENTOS INTERMEDIOS Los filamentos intermedios: tienen una función de sostén o estructura general. Están formados por sub-unidades no polares y muy variables. No poseen actividad enzimática y forman filamentos no polares. Las proteínas de los filamentos intermedios se caracterizan por un dominio bastonciforme (en forma de varilla) central con dominios globulares conservados en cada extremo. Se arman a partir de un par de monómeros helicoidales que se enroscan entre sí para formar dímeros súper-enrollados. Proteínas asociadas a filamentos intermedios son esenciales para la integridad de las uniones célula con célula y célula con matriz extra-celular. CENTRIOLOS Y CENTROS ORGANIZADORES DE MICROTUBULOS Los centriolos representan el punto focal alrededor del cual se ensamblan los MTOC. Son cilindros citoplasmáticos cortos, en pares, con forma de vara, formados por nueve tripletes de microtubulos. Suelen encontrarse cerca del núcleo, rodeados por el aparato de Golgi y asociados con una zona de material pericentriolar denso y amorfo. Centro organizador de los microtubulos o centrosoma: región de la célula que contiene los centriolos y el material peri-centriolar El MTOC es la región donde se forma la mayor parte de los microtubulos y desde donde ellos se dirigen a sus destinos específicos dentro de la célula. Controla la cantidad, polaridad, dirección, orientación y organización de los microtubulos formados durante la interfase del ciclo celular. Durante la mitosis, los MTOC duplicados sirven como polos del huso mitótico. Los centriolos proporcionan cuerpos basales para los cilios y los flagelos y alinean el huso mitótico durante la división celular. Las funciones de los centriolos Formación de cuerpos basales: se originan en una formación de novo sin contacto con los centriolos pre-existentes (mecanismo a-centriolar) o en la duplicación de centriolos existentes (mecanismo centriolar). Dan lugar a pro-centriolos, los que maduran a medida que migran al sitio apropiados cerca de la membrana celular apical, donde se convierten en cuerpos basales. El cuerpo basal actúa como centro organizador para un cilio. Los microtubulos crecen hacia arriba desde el cuerpo basal, empujan la membrana celular hacia fuera y se alargan para formar el cilio maduro. Formación del huso mitótico: durante la mitosis, la posición de los centriolos determina la ubica- ción de los polos del huso mitótico, son necesarios para la formación de un MTOC funcional, El papel principal de los centriolos en la mitosis es posicionar al huso mitótico en forma adecuada mediante el reclutamiento del MTOC desde el cual pueden crecer los microtubulos astrales y establecer el eje para el huso en desarrollo. La duplicación del centrosoma esta sincronizada con los eventos del ciclo celular y la ciliogénesis. Los cilios se ensamblan durante la fase G1; son los más abundantes en G0 y se desensamblan antes de que la célula ingrese en la fase M del ciclo celular. INCLUSIONES Las inclusiones contienen productos de la actividad metabólica de la célula y consisten principalmente en gránulos de pigmento, gotitas de lípidos y glucógeno. Se consideran componentes celulares sin movimiento y sin vida. MATRIZ CITPLASMÁTICA gel acuoso concentrado que está compuesto por moléculas de diferentes tamaños y formas. Muestra una estructura poco específica con la microscopia óptica Es el sitio de los procesos fisiológicos que son fundamentales para la existencia celular (síntesis y degradación proteica, degradación de sustancias nutritivas). Esta red proporciona un sustrato estructural sobre el cual ocurren las reacciones citoplasmáticas. Características histológicas de diferentes tipos celulares Descripción Células epiteliales en el riñón. CD: células cilíndricas con bordes bien definidos en el conducto colector TS: células escamosas en el segmento delgado de la nefrona VR: células aún más aplanadas revistiendo vasos sanguíneos, los vasos rectos del riñón Células ganglionares de la raíz dorsal. N: Gran tamaño de estos cuerpos de células nerviosas, núcleos pálidos (eucromáticos) con marcados nucléolos S: Células satélite aplanadas El tamaño de la célula ganglionar y la presencia de un núcleo eucromático, un nucléolo prominente y corpúsculos de Nissl (retículo endoplásmico de superficie rugosa visible como gránulos más oscuros dentro del citoplasma) reflejan la extensa actividad sintética necesaria para mantener los procesos excesivamente largos (axones) de estas células. 380X. Células musculares lisas del intestino delgado Células son típicamente alargadas, fusiformes y organizadas en una matriz paralela. Los núcleos también se alargan para adecuarse a la forma general de la célula. 380X. Fotomicrografía de células secretoras del páncreas. Las vesículas secretoras que contienen proteína lista para ser secretada llenan la porción apical de las células. Este proceso requiere un mecanismo de señalización externo para que la célula elimine los gránulos acumulados. 860X Fotomicrografía electrónica de un endosoma temprano. Muestra la estructura de un endosoma temprano de dictyostelium. Los endosomas tempranos se ubican cerca de la membrana plasmática y presentan una estructura tubulovesicular típica. Fotomicrografía electrónica autofagosomas en un hepatocito. de Se observan varios autofagosomas que contienen mitocondrias en proceso de degeneración. Los lisosomas circundantes que se han teñido con fosfatasa ácida. 12 600 X Fotomicrografía electrónica del retículo endoplásmico rugoso (RER). RER en una célula principal del estómago C: cisternas membranosas dispuestas muy juntas en formaciones paralelas M: mitocondria. En la superficie citoplasmática de la membrana se ven polirribosomas que rodean las cisternas. La imagen de una membrana repleta de ribosomas es el origen del término retículo endoplásmico rugoso. En el citoplasma hay unos pocos ribosomas libres. Fotomicrografía electrónica de vesículas con cubierta de COP-I Vesículas recubiertas de COP-I que inician el transporte retrógrado desde la red cis-Golgi hacia el RER. Se observa la estructura del CGN y las vesículas emergentes Fotomicrografía electrónica vesículas con cubierta de COP-II de Vesículas recubiertas de COP-II que son responsables del transporte anterógrado. Cubierta superficial de estas vesículas es diferente de las vesículas con cubierta de clatrina Fotomicrografía electrónica del REL. Muestra abundantes siluetas del REL en una célula intersticial (Leydig) del testículo, una célula que produce hormonas esteroideas. El REL, es un sistema complejo de túbulos anastomosados. Los pequeños objetos densos son partículas de glucógeno. Fotomicrografía de células plasmáticas. Muestra incrustada en plástico y teñida con azul de toluidina, se ve la lámina propia del intestino delgado. Las células plasmáticas bien orientadas exhiben una región clara en el citoplasma cercano al núcleo Fechas: regiones con tinción negativa representan acumulación de cisternas membranosas que pertenecen al aparato de Golgi. Fotomicrografía electrónica del aparato de Golgi. Muestra el extenso aparato de Golgi en un islote de células del páncreas. Los sacos aplanados de la membrana del aparato de Golgi se organizan en capas. CGN: está representado por vesículas aplanadas en la superficie convexa externa, mientras que las vesículas aplanadas de la región convexa interna constituyen la red trans-Golgi TGN). Estructura de la mitocondria Muestra una mitocondria en una célula de un ácino pancreático. La membrana mitocondrial interna forma la cresta (C) a través de una serie de pliegues, como resulta claro en la región de la flecha. La membrana mitocondrial externa es una envoltura continua y lisa, que está separada y es diferente de la membrana interna Fotomicrografías microtúbulos. electrónicas de Flechas: microtúbulos de un huso mitótico en una célula en división. A la derecha, los microtúbulos están adheridos a los cromosomas Fotomicrografía electrónica de la región apical de una célula epitelial demostrando la presencia de filamentos intermedios. TW: red terminal de una célula epitelial IF: filamentos intermedios subyacentes R: Los largos manojos rectos de filamentos de actina o raicillas se extienden desde las microvellosidades establecen uniones cruzadas con una densa red de filamentos de actina que contiene abundantes proteínas fijadoras de actina. Cuerpos basales y Cilios. Muestra los cuerpos basales y los cilios seccionados transversalmente, La disposición de los microtúbulos de los cilios en un patrón 9 + 2, en la que nueve microtúbulos en la periferia de los cilios rodean dos microtúbulos centrales. Los cuerpos basales carecen del par tubular central. En varios cortes transversales, se puede observar que desde el cuerpo basal se proyecta lateralmente el pedículo basal (asteriscos) Fotomicrografía de cuerpos de Mallory En la cirrosis hepática alcohólica, los hepatocitos presentan estas inclusiones (flechas), las que se conocen como cuerpos de Mallory. Los linfocitos y los macrófagos responsables de una reacción inflamatoria intensa, rodean las células que contienen cuerpos de Mallory