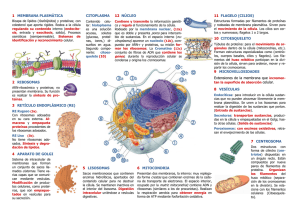
Last Universal Common Ancestor ARN→ PROTEÍNAS (1) ARN→ ADN (2) LUCA Conocido también como progenote o como último antecesor común, ya que dio lugar a los demás mediante divisiones sucesivas. A partir de este momento comienza la evolución biológica o biótica. Esto supone que los nuevos organismos que se creen van a sufrir mutaciones que en los casos más beneficiosos les conceden a los organismos características para adaptarse al medio, así como dotarles de una mayor diversidad. DESARROLLO DE LAS CÉLULAS 1) Procariotas anaerobias (no hay oxígeno en la atmósfera) y heterótrofas (estructuras más básicas). 2) Procariotas fotosintetizadoras anaerobias facultativas: liberan oxígeno a la atmósfera, esto supone un problema para el resto de las células anaerobias ya que o se escondían o morían. (1 Glucosa= 2 ATP). 3) Células aerobias heterótrofas: aunque son heterótrofas presentan ventajas biológicas como que por cada molécula de glucosa se producen 36 ATP. 4) Células eucariotas: surgen a raíz de la aparición de compartimentos que ofrecen ventajas (mayor organización, lo que supone una mayor facilidad para colaborar y regular las rutas metabólicas, y protección). Un ejemplo de ello es el material genético, este se encuentra mayoritariamente en el núcleo para así no ser destruido por enzimas del citosol como la nucleasa. TEORÍA ENDOSIMBIÓTICA DE LYNN MARGULIS Explica el origen de los cloroplastos y las mitocondrias basándose en la fagocitosis. Lo que Margulis sostiene es que las mitocondrias son origen de una digestión sin finalizar por parte de una célula anaerobia a una célula aerobia, a cambio de que la célula huésped contara con protección y comida. Los cloroplastos surgirían de la misma forma, a excepción de la naturaleza de la célula huésped, ya que, en este caso, sería fotosintetizadora. Las pruebas que demuestran la autenticidad de la teoría de Lynn Margulis son: -Membranas: ambos orgánulos poseen dos membranas, aunque son distintas entre ellas. -ADN (material genético): poseen su propio ADN. Es parecido al de las bacterias. -Ribosomas parecidos a los de las bacterias. Es decir, su velocidad de sedimentación corresponde a 70s (50s+30s). TEORÍA AUTÓGENA DE TAYLOR Y DODSON Proponen que algunos orgánulos celulares se originaron gracias a invaginaciones de la membrana celular. Un ejemplo de este proceso es el núcleo: -ADN unido a las membranas -Invaginación del ADN es cada vez más profunda -Aíslan el ADN. Lo rodean. Todas estas teorías son hipotéticas. Se sospecha que la realidad es una mezcla de todas ANTES DE LAS CÉLULAS EUCARIOTAS Antes de llegar a ser organismos pluricelulares, se formaron colonias. Se diferencian de las células eucariotas en que si se separan los organismos que componen las colonias estas no se desintegran. Estas colonias se forman con asociaciones de células porque TOGETHER WE ARE STRONGER, lo que además supone que se reparten el trabajo según las características de cada uno, esto implica que se pierdan sus otras capacidades, lo que supone la generación de una dependencia entre ellos. Así aparecieron los organismos pluricelulares. Uno de los ejemplos de colonias más conocidos es el VOLVOX (colonia protegida por los polisacáridos que los unen). TEORÍA CELULAR S. XVII 1) Robert Hooke: descubre las denominadas “celdillas” en las láminas de corcho que observa a través del microscopio. Aunque estas corresponden a los restos de paredes celulares de células muertas. 2) Anton Van Leeuwenhoek: era un comerciante de telas que, gracias a su curiosidad por observar la calidad de los tejidos, desarrolló microscopios lo suficientemente potentes como para observar lo que él denominaría como “animalículos” (microorganismos). Observaba muestras de semen, sangre (observó glóbulos rojos), … S. XIX 1) Dutcrochet: Todos estamos formados por células. 2) Schleiden (botánico) y Schwann (zoólogo): Apoyan a Dutcrochet y afirman además que la célula es una unidad funcional básica e independiente. 3) Rudolf Virchow: Estableció que toda célula proviene de otra preexistente. Así pues, niega la generación espontánea, pero no será posible demostrarlo hasta el experimento de Pasteur. 4) Weismann: Habla de los antecesores comunes POSTULADOS DE LA TEORÍA CELULAR 1) Toda célula viene de otra preexistente 2) La célula es capaz de mantenerse “per se” 3) Todo ser vivo está constituido por células 12) Membrana citoplasmática (manejan nutrientes y sustancias de desecho) 13) Poseen un programa genético (construcción, control actividades, autoduplicación) 14) Se reproducen CÉLULA PROCARIOTA • 4) Además, gracias a Golgi (reticularista) y a J. Ramón Jiménez se demostró que, incluso las células del tejido nervioso poseen independencia del resto. Es decir, no trabajan como una red como afirmaba el italiano. CARACTERÍSTICAS DE LAS CÉLULAS 1) Estructuras son muy complejas 2) Orden y regularidad internos. Este proceso para mantener el orden supone un elevado gasto de energía. Se aprovecha la primera Ley de la Termodinámica, que dice que la energía permanece constante. Un ejemplo de ello serían las mitocondrias (realizan hasta cuatro cambios energéticos: electrones, cadenas de transporte, movimiento, gradientes, …) 3) Tolerancia muy baja a los errores 4) Alto grado de regulación y control 5) Requiere aporte de energía. Para ello es fundamental 1ª Ley de la Termodinámica que permite intercambios en las formas de energía. E 6) Los mismos compuestos químicos básicos 7) Estructuras (fosfolípidos, proteínas, …) y procesos muy similares (dogma central de la biología molecular) 8) Reacciones químicas son muy variadas (metabolismo celular) 9) Se autorregulan (control metabólico, corrección de errores→ si un error se perpetúa es por la elevada velocidad a la que se produce) 10) Sistemas de sensores que regulan procesos 11) Actividades mecánicas (transporte, síntesis, degradación, …) • Arqueobacterias: viven en ambientes hostiles (similares a los que podía haber hace miles de millones de años), como, por ejemplo: metanógenos (atmósfera de metano), acidófilos (ambientes con un bajo pH), basófilos (ambientes con pH elevado), termófilos (altas temperaturas), halófilos (elevada concentración salina, es decir, podían sobrevivir a la salazón). Este tipo de bacterias no son patógenas, porque no se adaptan a las condiciones de nuestro organismo. Eubacterias: Son células patógenas ya que logran sobrevivir en las condiciones habituales de las células. Algunos ejemplos de eubacterias son las micoplasmas (no tiene pared celular) y las cianobacterias (fotosintetizadoras). Los ejemplos anteriormente mencionados corresponden a eubacterias que poseen características excepcionales. MORFOLOGÍA BACTERIANA La morfología nos puede indicar el género o la especie de las bacterias. Los tipos de morfología son: ✓ Cocos: 1) Diplococos (la parejita) 2) Estreptococos (fila india). Ej.: microorganismos epiteliales 3) Estafilococos (racimos) ✓ Bacilos (bastoncillos) ✓ Vibrios/ vibriones (coma). Ej.: cólera ✓ Espirilos/ espiroquetas (hélices). Ej.: sífilis. ESTRUCTURAS OBLIGADAS Pared celular (algunas poseen moléculas que actúan como antígenos) Membrana celular (similar a la nuestra pero los lípidos son distintos, no suele haber esteroles) Citosol Cromosoma bacteriano Ribosomas cabo el proceso de transcripción, en el que destaca la acción de las topoisomerasas. FACULTATIVAS Flagelo Pili Plásmidos Cápsula Esporas PARED CELULAR Las paredes celulares se clasifican según su reacción ante la tinción de Gram: 1) Gram- = moradas→ biestratificado (bicapa lipídica membrana exterior) 2) Gram+ = rosa→ monoestratíficada (membrana interna y pared celular de peptidoglucano y ácidos teitoicos). El proceso que se lleva a cabo para la realizar la tinción es: 1) 2) 3) 4) Teñimos con el colorante violeta Lavamos Teñimos con el colorante rosa Lavamos CROMOSOMA BACTERIANO Doble hélice→ ADN en forma de anillo cerrado, que está superenrollado gracias a la acción de las topoisomerasas. Este superenrollamiento presenta ventajas como la protección frente a la acción de nucleasas. Sin embargo, también posee desventajas como la necesidad de desenrollarse para llevar a PLÁSMIDOS “Es como el cromosoma bacteriano en versión mini”. Llevan información que puede aportar ventajas a las bacterias a la hora de sobrevivir y de adaptarse al medio que las rodea. Ej.: algunos plásmidos llevan genes que proporcionan la resistencia a antibióticos como la penicilina. Sin embargo, este tipo de información solo es útil en el momento de enfrentarse a los antibióticos. FLAGELOS: Los flagelos de las células procariotas también son distintos a los de las células eucariotas debido a que estos últimos poseen proteínas estructurales como la actina. El funcionamiento de los flagelos consiste en giros. Los flagelos confieren la ventaja de elegir la dirección hacia la que se quieren desplazar. Hay diversos tipos dependiendo de su localización y su cuantía. PILI Hay dos tipos: los pili sexuales (sirven para la reproducción sexual y el paso de información de una bacteria a otra) y los pili de adhesión que aportan a las células la capacidad de adherirse a las superficies de un determinado organismo y así ser eliminado más difícilmente. Por ello se denominan factores de virulencia. Ej.: burlan mecanismos de limpieza como la urea. CÁPSULA Situada por encima de la pared está formada principalmente por polisacáridos lo que le confiere una mayor capacidad de adherencia. Además, también se encarga de esconder los antígenos, de tal manera que el sistema inmune solo detecta los polisacáridos. Debido a su composición, no se tiñen bien con colorantes, así que hay que emplear otros métodos de tinción (acidófilos, basófilos). ESPORAS Mientras que organismos como los hongos las emplean para reproducirse, para las bacterias es un método de adaptación a un medio que ha sufrido cambios. Ej.: Bacillus anthracis (aerobia estricta). Las esporas consisten en el recubrimiento mediante la membrana y otras dos capas duras de elementos obligados para la supervivencia de la bacteria: cromosoma bacteriano y ribosomas. Cuando el medio vuelve a la normalidad, la bacteria recupera el 100% de su funcionalidad. A este proceso se le denomina germinación. DUPLICACIÓN DE LAS CÉLULAS PROCARIOTAS: FISIÓN BINARIA 1) Antes de la división se tienen que dar una serie de condiciones: tamaño crítico y la unión de los cromosomas a la membrana. 2) Durante la síntesis se crean también la membrana y la pared, que equivalen a un tabique. CÉLULAS EUCARIOTAS TIPOS: 1) Protistas: unicelulares. Ej.: diatomeas. 2) Hongos: unicelulares (Ej.: levaduras) o pluricelulares (Ej.: setas) que poseen pared celular distinta a la de las bacterias. 3) Vegetales: posee pared celular distinta a las anteriores. 4) Animales: Estudiaremos la célula prototipo= Lo tiene TODO. FUNCIONES DE LOS DISTINTOS COMPONENTES DE LA CÉLULA 1) Membrana: Forma + Intercambios 2) Citoesqueleto: movilidad + forma + aguante de la presión osmótica + cilios + flagelos + microvellosidades 3) Citosol: medio fluido que actúa como contenedor en el que se llevan a cabo reacciones químicas 4) Ribosomas: (síntesis de proteínas, las del RER y el citosol son distintas) 5) RE: liso (lípidos) y rugoso (proteínas) 6) Aparato de Golgi: maduración + empaquetamiento de vesículas para trasnporte+ síntesis de glúcidos 7) Lisosomas y peroxisomas: digestiones 8) Mitocondrias: energía 9) Núcleo: CONTROL TABLA COMPARATIVA ENTRE PROCARIOTAS Y EUCARIOTAS FORMAS ACELULARES VIRUS También denominados agentes filtrables ya que eran capaces de filtrarse por los minúsculos agujeros que se realizaban en matraces de Erlenmeye (cosa que las bacterias no podían hacer). • • Características: 1) Pequeño tamaño 2) Solo poseen proteínas (podían ser todas del mismo tipo o de distinto tipo) y material genético 3) El mat.genético solo puede ser o ADN o ARN 4) Agentes patógenos obligados. Por sí mismos no puede llevar a cabo acciones fundamentales para la vida como un correcto desarrollo del metabolismo. Por ende, siempre se van a beneficiar de otros seres vivos para que realicen esas funciones por ellos. 5) Para su supervivencia es fundamental que posean la capacidad de penetrar, adaptarse y salir de lás células. 6) Cápside: formada por proteínas (=capsómeros) que van a proteger al material genético formando una nucleocápside. Asimismo, pueden poseer una envuelta (=forma irregular) compuesta por una bicapa lipídica y proteínas. 7) Capacidad de autoensamblaje (se juntan como piezas de un puzzle en un medio acuoso), que se facilita cuanto mayor sea el número de copias del virus. Importante destacar: · No son seres vivos→ tienen que ser infecciosos para sobrevivir→ se aprovechan de las enzimas de la célula huésped para generar copias de sí mismo y asói poder replicarse. “La célula trabaja para él”. TIPOS DE CLASIFICACIÓN 1) Estructura: -Tamaño, morfología, ácidos nucleicos. Ej.: picornavirus→ Pico= tamaño; Rna= tipo de ácido nucleico. Ej.:hepadnavirus→ Hepa= hepatitis; Dna= tipo de ácido nucleico. 2) Características bioquímicas: -Estructura, genoma, modo de replicación. -Clasificación según su genoma: ADN ARN Bicatenario Solo hay que replicarlo para que se introduzca en el genoma celular. Monocatenario Es necesaria la creación de la cadena complementaria Para poder replicarse creando copias de sí mismo, es necesario que el virus lleve consigo el genoma para sintetizar la enzima ARNpolimerasaARNdependiente. Se emplea la cadena ARNpara sintetizar ARN+. ARN +: se sintetiza su cadena complementaria, que ha de ser leída para generar más copias del ARN+. Estas van a dar lugar a la formación de proteínas virales y, cuando haya suficientes, sintetizarán las nuevas partículas virales. ARN-: mismo proceso pero al revés. ARN+→ADN: para este proceso se emplea la ADNpolimerasa- ARN dependiente= trasncriptasa inversa= retrotrasnscriptasa. 3) Enfermedades (poco útil): -Virus de la hepatitis -Virus encefalitis 4) Tipo de huésped: -Bacterias (bacteriófogos)→ Estructura especial -Vegetales Es necesario que encaje con los receptores de membrana. -Animales Todas los virus tienen un rango de huéspedes: cantidad de especies que pueden infectar. Pueden mutar y realizar un “salto de especie”. Asimismo, los virus animales y vegetales a la hora de infectar introducen la nucleocápside, mientras que los bacteriófagos solo introducen su genoma. Además, antes se pensaba que la forma determinaba la especie a la que un virus infectaba, sin embargo, eso no es así. ESTRUCTURA DE LOS VIRUS 1) Tamaño: 18-300 nm 2) Desnudos: Genoma (ADN o ARN) + Cápside (proteínas= capsómeros) 3) Envuelta: bicapa lipídica + glucoproteínas 4) Si el genoma es ARN llevarán enzimas accesorias o esenciales VIRUS DESNUDOS ESTRUCTURAS 5) Tropismo tisular: No todos los virus pueden infectar a todas las células del organismo. Ej.: adenovirus, enterovirus, … EJEMPLOS DE FAMILIAS VIRALES HELICOIDAL ICOSAÉDRICO BACTERIÓFAGO PROPIEDADES DE LOS VIRUS SEGÚN SU ESTRUCTURA VIRUS DESNUDOS VIRUS CON ENVUELTA Resistencia -Detergentes: SÍ -Ácidos: SÍ -Desecación: SÍ Ya que posee una nucleocápside, formada por capsómeros. Resistencia -Detergentes: NO -Ácidos: NO -Desecación: NO Porque están compuestos por lípidos, cuyos enlaces son más fáciles de romper que los enlaces peptídicos. Mayor dificultad de que se propaguen, aunque lo hacen mediante secreciones. Fácil propagación Fómites: a partir de superficies contaminadas. Fecal- Oral Contacto Al salir de la célula, realizan tantas copias, que van a producir la lisis celular (= lesiones en los tejidos). Fase crítica en el sistema inmune. Forma irregular (=pleomorfos) Pueden evacuar la célula huésped mediante lisis o gemación (mediante la cual, la célula solo muere si las evaginaciones son muy heavys) Ambos poseen espículas (proteínas o glucoproteínas, algunas son VAP). CICLO DE REPLICACIÓN VIRAL 1) Adhesión (necesitan generar enlaces con los receptores) 2) Introducción nucleocápside (vegetales o animales). -Desnudos: Viropexia o por endocitosis mediada por receptores. -Envueltos: Endocitosis mediada por receptores o por fusión de membranas. 3) Descapsidación: destrucción de la cápside para que se pueda liberar el material genético en el citosol. 4) Síntesis. 5) Ensamblaje. 6) Salida mediante lisis o gemación. CICLOS VIRALES LÍTICO El virus entra en la célula Replicación y síntesis de ADN. Ensamblaje. Lisis (=liberación). LISOGÉNICO El virus entra en la célula Se esconde en el ADN celular donde se puede convertir en un profago (fago) o provirus (virus). LATENTE. No se replica de manera tan rápida, lo hacen de manera que se puedan ocultar más fácilmente y sea más seguro propagarse. Cuando el virus sale del genoma de la célula huésped empieza el ciclo lítico. Este cambio se podría deber a factores ambientales. Ej.: 1) Virus influenza→ se introduce en el medio interno mediado por receptores (entra y sale de la célula varias veces). 2) Virus VIH→ Entra por fusión de membranas y contiene información para la síntesis de varios tipos de enzimas. Es importante saber las fases por las que pasa el virus para poderle poner puertas al campo. En el VIH el periodo latente o ventana se puede mantener hasta diez años. FORMAS ACELULARES II -VIROIDE: los viroides son más sencillos que los virus. Están formados únicamente ácidos nucleicos (240-600) aunque su número es menor que en los virus. Afectan a vegetales (el motivo por el que empiezan las investigaciones para saber de qué se trata es la pérdida de dinero). Poseen actividad catalítica. Alteran el funcionamiento de las RNA polimerasas para que la célula solo pueda sintetizar su ARN, lo que produce que la célula disminuya su tamaño, altera también su regulación genética. -Prion (Protein infection): Solo están formados por proteínas, por ello se pensaba que eran virus lentos (ya que al no tener los mismos componentes tampoco tenían la misma velocidad). Los priones afectan tanto a animales como a plantas. • Tipos: -Scrapie (tembladera): afecta al ganado ovino y no es transmisible a humanos. -Kuru (temblor): en humanos podía derivar en locura. Tiene largos procesos de incubación y se transmite mediante la ingestión. Esta enfermedad es de Papúa Nueva Guinea (endémica). -Enfermedad de Creutzfeld-Jacob (CJD): se da en seres humanos. La causa de la enfermedad puede ser: esporádica (>85%) producidas por cambios espontáneos como las mutaciones, familiar (15%), infecciosa (por ingestión= vCJD) o iatrogénica (en procedimientos médicos). -Encefalopatía espongiforme bovina (BSE): afectaba al ganado bovino, produciendo temblores y posible locura y es transmisible a humanos. Produce lesiones en el tejido nervioso que se ven como agujeros (neuronas muertas). Es incurable ya que las neuronas no se regeneran. (vCJD). -Prion: contiene pocos aminoácidos, pero son los mismos que los aa presentes en la membrana celular, aunque plegados de distinta forma. α = aminoácidos normales (Proteína priónica celular)→(PrPc)// predomina la hélice alfa. Β = aminoácidos proteína infecciosa. Proteínas muy estables, las cuales se acumulan formando agregados (Proteína priónica scrapie)→ (PrPsc)// predomina la lámina beta. La replicación de priones se produce mediante el contacto de dos proteínas distintas (una priónica y otra no priónica)→ Mediante la interacción de ambas, la priónica obliga a la otra a transformarse, pasando a ser priónica también. Esto supone un crecimiento exponencial. MEMBRANAS Todas las membranas tienen unas características y unas funciones comunes, aunque existen moléculas que sirven como método de diferenciación. Hay tres tipos de membranas: 1) Membrana citoplasmática (procariotas y eucariotas) 2) Membranas de orgánulos celulares (mitocondria con la ATPasa, RE, Aparato de Golgi, núcleo, lisosomas, peroxisomas) 3) Envuelta del virus MEMBRANAS. CARACTERÍSTICAS - Se caracterizan por poseer una estructura trilaminar. Ya que hay dos zonas más electrodensas y una, situada en medio, menos. Antes se achacaba a los lugares en los que se situaban los lípidos, los glúcidos y las proteínas. - Componentes: 1) Lípidos: pueden ser fosfolípidos (responsables de la forma plana y fina y del paso de sustancias apolares y pequeñas= semipermeabilidad selectiva) y el colesterol (encargado de regular la fluidez). 2) Proteínas (les concedden distintas características, además de estar encargadas de funciones como el transporte). 3) Glúcidos: encargados de “etiquetar a la célula”. -Semipermeabilidad selectiva: permite la creación de gradientes químicos y eléctricos a un lado y al otro de la membrana. En esta semipermeabilidad también participan las proteínas transportadoras. Estos componentes están presentes en distintos porcentajes según el tipo de célula. Por regla general suele ser 50%. Aunque en las neuronas el 70% son lípidos y en los eritrocitos las proteínas son mayoritarias. -Distinta composición a nivel intracelular y extracelular y entre el citoplasma y los orgánulos. Esto preserva los gradientes y es debido a la especialización. Las uniones establecidas entre los diversos componentes son uniones débiles para aportar fluidez a la membrana y que se puedan hacer y deshacer de manera más sencilla. Cada uno de los componentes de la membrana aporta una serie de características: Al pasar de un estado DESfavorable a un estado favorable, el sodio libera energía -Comunicación mediante receptores: esto permite saber las células con las que las diferentes sustancias se pueden unir. Ej.: 1) Bicapa lipídica: estructura general y semipermeabilidad. Caracterizada por los fosfolípidos. CABEZA POLAR→ LUZ División COLA APOLAR CABEZA POLAR→ CITOPLASMA 2) Proteínas: aportan características individuales (cada membrana posee unas proteínas específicas). También se encargan del resto de funciones. 3) Oligosacáridos: se encuentran en la monocapa externa de la membrana citoplasmática y en la monocapa interna de los lisosomas. Unión Hormona MEMBRANAS. PROPIEDADES ·Mosaico fluido→ muchas cosas juntas en continuo movimiento (siempre respetando un orden para cubrir huecos). Esto permite que la membrana no sea ni sólida ni líquida, sino que sea fluida. ·Son muy finas: unos 7,5 nm. MEMBRANAS. FUNCIONES -Define la forma y la extensión de la célula y sus orgánulos -Catálisis de reacciones metabólicas: Enzimas FUNCIONES SEGÚN ESTRUCTURAS Bicapa lipídica Integridad de la célula y orgánulos Semipermeabilidad selectiva Transportadoras: transporte controlado de moléculas polares 1) Fosfoglicéridos: Los fosfoglicéridos están unidos por una cadena hidrocarbonada a la que se unen dos ácidos grasos y un grupo fosfato al que, a su vez, se unen alcoholes como la serina (carga total negativa→no tiene carga), etanolamina (carga total negativa +/-) y la colina (carga total neutra +). Según el alcohol que utilicen tendrán distinta carga y distinto tamaño. Proteínas De membrana: Conexiones entre células Soporte y mantenimiento de la célula Enzimas Regulación de reacciones metabólicas Receptores de membrana Sensibilidad a sustancias químicas reguladoras Componentes de membrana (marcadores) 2) Esfingolípidos/ Glucolípidos: (aminoalcohol de cadena larga). Señalización celular LÍPIDOS Aunque por regla general el número de lípidos sea mayor que el de proteínas, tienen menor peso molecular. - Fosfolípidos: están formados por una cola apolar y una cabeza apolar. Esta cola está formada por 2 ácidos grasos, uno insaturado y otro saturado. Dentro de los fosfolípidos se pueden distinguir dos tipos: Presenta esfingosina Los esfingolípidos están formados por esfingosina a la que se unen un ácido graso, que puede ser saturado o insaturado (formando una ceramida), se unen a una cabeza polar que puede ser una de las mencionadas anteriormente (fosfatidilcolina) o un oligosacárido (gangliósido) o un monosacárido (cerebrósido). Entre la cola apolar de la esfingosina y el ácido graso quedan huecos que permite el paso de pequeñas moléculas apolares como el oxígeno o el dióxido de carbono. Cuando los fosfolípidos se encuentran es un medio acuoso se vuelven inestables, para evitar esta reactividad, forman bicapas lipídicas en forma de micelas→ LIPOSOMAS - Esteroles (colesterol): los esteroles también son anfipáticos, pero la parte hidrófila es menor, ya que contiene un solo OH. Asimismo, al estar formado por cuatro anillos rígidos posee poca capacidad de movimiento (paso estrella= FLIP-FLOP; también realiza flexión). El colesterol aporta hidrofobia (por ello se coloca entre los fosfolípidos) y es más corto que los fosfolípidos. Además, tiene capacidad auto selladora lo que permite reparar la célula y así evitar su lisis. El colesterol aumenta la rigidez, pero impide la cristalización, es decir, pone orden. Esto es posible ya que impide el movimiento en la monocapa en la que se encuentra, pero, a su vez, aumenta la distancia entre los fosfolípidos de la otra monocapa. Eso sí, el colesterol se encuentra en la misma proporción en ambas partes, pero NUNCA hay dos moléculas de colesterol enfrentadas. ÁCIDOS GRASOS Los carbonos de los ácidos grasos se unen mediante enlaces covalentes simples, lo que permite que posean capacidad de rotación. Sin embargo, los ácidos grasos entre sí están unidos por Fuerzas de Van den Waals, lo que permite los movimientos que realizan los fosfolípidos (se suelen mover 1 micra/s). Los movimientos más comunes son: 1) Difusión lateral (rotura de enlaces) 2) Rotación: al rotar un fosfolípido, el ácido graso insaturado que poseía cambia de dirección, permitiendo acercamientos que antes no eran posibles. 3) Flexión: obliga a los demás a moverse (al igual que la rotación). Las colas apolares se acercan o se alejan según los giros del carbono 4) Flip-Flop: es raro que sea espontáneo debido a la cabeza apolar. Tiene que ser rápido para que no se cuelen “cosas”. Para que este movimiento se lleve a cabo necesitan ayuda de las proteínas conocidas como flipasas. Los ácidos grasos influyen sobre la fluidez de varias formas: - - La longitud de las colas (14-20): a mayor longitud, más enlaces se establecen entre ellas y, por ende, más enlaces hay que romper. Las membranas de los orgánulos son especialmente finas lo que supone un menor número de carbonos y de colesterol que hace que sea más fluida. Dobles enlaces (=insaturaciones): a mayor distancia entre átomos, menos fuertes son los enlaces, lo que supone una mayor fluidez. A mayor temperatura, hay una mayor ruptura de enlaces, lo que supone una mayor fluidez. Para conservar esta fluidez son fundamentales los movimientos de difusiones laterales. Las funciones de la fluidez son: 1) Lugar definitivo tras la síntesis: las proteínas de membrana sintetizadas en el RER (evaginación)→ Aparato de Golgi (fusión de membranas)→Membrana citoplasmática (fusión de membranas=recolocación ácidos grasos). 2) Fusiones de membrana. 3) Señalización: Sustancia se une a un receptor de membrana→ Llega una enzima→Enzima= María Antonieta→ Cabeza fosfolípido (¡Hey, Danny!) 4) Reparación de lesiones (“agujerillos ocasionados movimiento”): pueden reparar hasta 0,2 micras. por el ASIMETRÍA -La composición de las dos monocapas es distintas, ya que en la externa es en la que se encuentran los glúcidos. Además, dependiendo de la monocapa, nos encontramos con distintos fosfolípidos. En la monocapa externa suelen ser neutros como la fosfatidilcolina (componente mayoritario de la monocapa externa). Sin embargo, en la monocapa interna suelen ser negativos: fosfatidilserina, fosfatidiletanolamina o fosfatidilinositol (mayoritarios en la monocapa interna). -La función: las células que han muerto, pueden ser un problema si siguen en nuestro organismo, por ello, es necesario un macrófago que las fagocite. Para identificar las células muertas, se emplea la fosfatidilserina. Cuando una célula está viva, existe una flipasa específica que impide que la fosfatidilserina salga a la monocapa externa. Sin embargo, cuando la célula está muerta, la flipasa no actúa como debería, lo que supone que la fosfatidilserina sale de la monocapa interna. -Glucolípidos: siempre se encuentran en el exterior. Representan el 5-10% de los lípidos de la membrana. FUNCIONES DE LA ASIMETRÍA 1) Protección en la zona apical de los epitelios: como los glúcidos se encuentran en el exterior de la célula, actúan como protectores. Pueden protegerlas desde un punto de vista químico (como el medio puede ser especialmente ácido, en vez de desnaturalizar a los lípidos o proteínas de la membrana, desnaturalizan a los oligosacáridos, que se vuelven a regenerar). También puede ser una protección mecánica (les protege frente a la fricción, debido a su viscosidad. 2) Campo eléctrico→ forma gradientes y señales eléctricas. 3) Reconocimiento: mediante las lectinas se reconocen a los oligosacáridos específicos. Ej.: GM1→ receptor del cólera= genera diarrea. Este receptor se encuentra en células intestinales. PROTEÍNAS DE MEMBRANA 1.3) Las proteínas realizan la mayor parte de las funciones: 1) Transporte 2) Anclaje de macromoléculas. Su unión a la matriz extracelular determina su forma, si se puede cambiar esa forma mediante comunicación mecánica (fuerzas) y también la capacidad de enviar mensajes. También se puede unir al citoesqueleto mediante los filamentos de actina 3) Receptores específicos (hormonas, neurotransmisores, …) 4) Enzimas 1.4) Barril β: Suelen ser muy rígidas y por eso no son tan comunes como las anteriores, también se encargan del transporte. Ej.: porinas, MME y procariotas (en las dos últimas están plegados en forma de lámina beta antiparalela). Pueden ser también receptores. Los lazos no iguales en el dominio intracelular y en el extracelular. Hélice α anfipática incrustada en el monocapa interna: encargada de la señalización intracelular. Señalización También conceden a las membranas características específicas. Ej.: bomba sodio-potasio ATPasa= citoplasmáticas // ATP sintetasa= membrana interna mitocondrial. Cuanto más trabajo realice una célula mayor número de proteínas habrá. Tipos proteínas 1) Integrales/Intrínsecas: son necesarias para la supervivencia, si las elimino, crearían un gran agujero en la célula y esta moriría. -Representan el 70% de las proteínas de membrana. -La mayoría son transmembranales: atraviesan por completo la membrana. Pueden de 1 solo paso (solo la atraviesan una vez) o multipaso (la atraviesan varias veces). -Son anfipáticas. -Tanto las transmembranales como las NO transmembranales de la monocapa externa poseen oligosacáridos, es decir, son glucoproteínas (no son proteoglucanos, porque poseen más proteínas que glúcidos). Los tipos más relevantes de proteínas intrínsecas son: 1.1) 1.2) Transmembranales de un solo paso (dominio extracelular y dominio plasmático). Suelen ser receptores. Transmembranal multipaso: encargadas del transporte. También pueden ser receptores. 2) Extrínseca/ Periférica: se pueden quitar sin producir la muerte celular. Pueden estar unidas a la monocapa interna o externa→ son hidrófilas. Los tipos de unión pueden ser: 2.1) Enlaces covalentes con la cadena lipídica. 5. 2.2) Enlaces covalentes con el grupo fosfatidilinositol (GPI). 6. 2.3) Enlaces débiles con proteínas de membrana (Ej.: puentes de hidrógeno). Si estos enlaces se establecen el interior también tiene actividad catalítica o de transducción de señales. 7 y 8. PROTEÍNAS TRANSMEMBRANALES Presentan diferente forma, carga y tamaño dependiendo del tipo que sean. Uno de los ejemplo de proteínas transmembranales más conocidos es la cisteína, que se aprovecha de que el medio externo está oxidado (a diferencia del medio interno que está reducido), para establecer uniones puentes disulfuro. La cisteína es de un solo paso. Las proteínas transmembranlales atraviesan toda la bicapa lipídica, pueden tener diversas formas y cuando se encuentran en el interior de la membrana pueden ser: hélice alfa, beta lámina. Asimismo poseen una orientación específica en el espacio, lo que dota a las células de asimetría y de diferentes funciones dependiendo del dominio (citosólico o externo). Esto permite que las células estén polarizadas. GLUCOPROTEÍNAS -La mayoría de las proteínas de membrana son sintetizadas por el RER (menos las proteínas de la monocapa interna) ya que les añade oligosacáridos (Golgi), también se les puede añadir sulfuro (cisteína). En cualquier caso, siempre se encuentran en la monocapa externa. -Los oligosacáridos no son especialmente grandes (menos de 15 unidades). Sin embargo, suelen están ramificados (=gran variedad). Esto es fundamental para aportar información de la célula y, sobretodo, recibirla. FLUIDEZ DE LAS PROTEÍNAS Las proteínas presentan una mayor dificultad de movimiento debido a la gran cantidad de enlaces que forman: uniones con el citoesqueleto, a la matriz extracelular (como el colágeno), a oligosacáridos (que a la vez establecen enlaces con otros). Sin embargo, también se mueven. Es imposible que realicen movimientos como el flip-flop (cargas y enlaces), pero realizan otros como la difusión lateral (más despacio que los fosfolípidos), rotación y movimientos similares a la flexión (=cambios conformacionales). Para demostrar que las proteínas se movían hicieron el siguiente experimento: Tintaron proteínas de membrana de una célula humana y otras de una célula de un ratón. Al unirlas, formaron un heterocaryon, ya que al ser células de dos especies diferentes, los núcleos no se podían mezclar. Y observaron, hasta que vieron que los distintos colores de las proteínas habían cambiado de lugar. Así también se descubrió que las proteínas no llevaban a cabo los movimientos de flip-flop, dado que el número de proteínas seguía siendo el mismo. Sin embargo a nuestras células les interesa que se limite el movimiento de estas proteínas, ya que estas han de situarse en regiones específicas para poder llevar a cabo su función de manera efectiva. Ej.: si las proteínas se encargan del transporte de nutrientes, han de estar cerca de la fuente que se los proporciona. UNIONES ESTRECHAS PARA CREAR DOMINIOS Algunos ejemplos del empleo de este tipo de uniones para la creación de dominios son los epitelios renales o los intestinales. DOMINIO/ POLO APICAL Para ello se crearon los dominios y polos, posibles gracias a las balsas lipídicas. BALSAS LIPÍDICAS/ LIPID RAFT Exceso de nutrientes Zonas en las que las colas de los ácidos grasos son más largas, hay menos insaturaciones, más colesterol y gran cantidad de oligosacáridos. Es importante la creación de dominios cuando una proteína realiza una acción distinta al resto (caveolas→tipo de balsa lipídica y endocitosis, proteínas G, transducción de señales) DOMINIO/ POLO BASOLATERAL STOP ERITROCITOS DOMINIOS A parte de las balsas lipídicas, existen otras formas de crear dominios: 1) Agregados proteicos: si uno un gran número de proteínas entre sí, el movimiento será mucho más complejo. Ej.: espermatozoides. 2) Unión de proteínas a la matriz extracelular. 3) Unión de proteínas al córtex celular (citoesqueleto). Ej.: eritrocito. 4) Uniones estrechas. Los eritrocitos unen sus proteínas de membranas para así tener una mayor resistencia mecánica. Esto les permite resistir los cambios de presión del plasma, ya que las células no son las que soportan la fuerza mecánica que se ejerce sobre ellas, sino que lo hace el citoesqueto, haciendo a los eritrocitos más resistentes. A este fenómeno también ayuda su forma bicóncava y su gruesa membrana (que permite el paso de un mayor número de moléculas de oxígeno y de dióxido de carbono). GLUCOCÁLIX Está formado por hidratos de carbono (=oligosacáridos mayoritariamente). En el glucocálix se distinguen los glucolípidos y las glucoproteínas, los cuales varían su forma según el tejido o célula a los que pertenezcan. Es decir, funcionan como una etiqueta, por eso se encuentran en la parte más externa. Funciones: 1) Adhesión célula-célula: gracias a su viscosidad. Las lectinas de una célula renocerá los oligosacáridos de la otra y se unirán. Esto les permitirá llevar a cabo procesos como la coagulación, la fecundación, respuesta inflamatoria (endotelio). 2) Protección frente a agresiones químicas (Ej.: intestino, donde las microvellosidades se encargan de la absorción y el glucocálix de que no se desintegre) o mecánicas (eficaz lubricante). 3) Permite la unión de distintas moléculas como por ejemplo hormonas, enzimas, virus o bacterias. TRANSPORTE A TRAVÉS DE MEMBRANA Las membranas son estructuras semipermeables, es decir, “eligen quíen pasa”. Dependiendo de la función que tenga la membrana necesitará unos transportadores u otros. Gracias a esta permeabilidad selectiva, se producen gradientes, ya que da lugar a distintas concentraciones de solutos a ambos lados de la membrana. Los transportadores de la membrana permiten a esta captar nutrientes, secretar sustancias y regular la concentración de solutos a nivel intracelular. Para todo menos para el oxígeno y el dióxido de carbono se necesitan proteínas transportadoras que, aunque todas tengan un funcionamiento similar, van a ser de distintos tipos en función de la membrana en la que se encuentren. Aunque el interior de la célula sea NEUTRO, como en ella existen proteínas y ácidos nucleicos, el número de cationes tiene que ser muy elevado para compensar las cargas negativas que estas producen. Los gradientes se emplean también para “trabajar” (gradiente de sodio+, gradiente de potasio+, gradiente de cloro- y gradiente de Ca2+) gracias a la energía potencial que generan. Es especialmente importante para saber cómo funciona la célula la concentración del Ca2+, ya que si es muy alta puede indicar (dependiendo del tejido al que pertenezca): - Fibra muscular: necesidad de contraerse. Neurona: necesidad de mandar neurotransmisores. Células secretoras: necesidad de secretar. TASA DE DIFUSIÓN Mide cómo de sencillo es para una determinada molécula atravesar la célula, sin contar con los transportadores. Esto depende de su tamaño y de su carga: -Moléculas hidrófobas: pasan entre los huecos establecidos entre los fosfolípidos. En concreto, como las hormonas esteroideas son derivadas del colesterol, estas atraviesan la membrana mediante el flip-flop o “infiltrándos”". -Pequeñas moléculas polares: como por ejemplo el agua, el glicerol o la urea. En el caso del agua, de todas las moléculas pasará una pequeña parte, pero la tasa de éxito es mayor que la del glicerol. -Grandes moléculas polares: la glucosa o la sacarosa pueden pasar a la célula pero sería un proceso tan lento que la célula no podría obtener toda la cantidad de glucosa que requiere en un determinado lapso de tiempo y moriría. Por eso, son necesarias las proteínas transportadoras. -Pequeñas moléculas polares (iones) con carga: Necesitan proteínas para atravesar la membrana, ya que son repelidas por los fosfolípidos. TIPOS DE PROTEÍNAS TRANSPORTADORAS POROS PERMEASAS BOMBAS No consumen ATP Viajan a favor de gradiente No se establecen uniones No hay cambios conformacionales Ej.: acuaporinas/ canales iónicos Pasivo Se establecen uniones Hay cambios conformacionales (aunque luego se recupera la forma) Activo Se establecen uniones Hay cambios conformacionales (aunque luego se recupera la forma) 2) Canales regulados por ligandos extra o intracelulares: proteína que “tira”. Ej.: hormonas. 3) Canales regulados mecánicamente: regulados por presiones. Ej.: oído. PERMEASAS Y BOMBAS Formación de enlaces entre las sustancias que transportan y la proteína. Esto da lugar a un alto grado de especifidad que puede ser menor o mayor. 1) 2) 3) 4) Formar enlaces DÉBILES. Son específicas. Multipaso. Cambio conformacional reversible. Si la proteína se une al sustrato, se va a poder saturar (= alcanza la velocidad máxima). Es decir, es un funcionamiento muy similar al de las enzimas. Si no hay uniones NO se puede saturar. POROS HIDROFÍLICOS/ CANALES DE FUGA/ CANALES PASIVOS Permiten el paso de moléculas sin ayuda. 1) Difusión simple (siempre abiertos)→ las moléculas pasan por movimientos al azar y a favor de gradiente. 2) Proteínas transmembranales multipaso. 3) Anfipáticas (exterior= no carga // interior= sí carga). 4) Factores limitantes: carga y tamaño. 5) Ej.: acuaporinas. ➔ Canales iónicos: poseen compuertas (abiertos o cerrados) y son más específicos que los poros. · Si no hay uniones, …. ¿cómo seleccionan? Pues mediante limitaciones de tamaño (muy grande= no cabe y muy pequeño= no excita) y de carga. Los canalaes iónicos se pueden regular mediante: 1) Canales regulados por voltaje: receptores captan el cambio de potencial y abrirán las compuertas. Vmáx Concentración de moléculas transportadas La Km (constante de Michaelis) nos indica la concentración perfecta (=Vmáx/2). Un indicador de la especificidad del soluto sería la Km, porque marca la afinidad. Ej.: transportadores de oligosacáridos que transporten mejor a la Glu que a la Man. Ej.: Endocitosis, exocitosis y fagocitosis. FUENTES DE ENERGÍA -Moléculas sin carga: concentraciones químicas. Es decir, se mueven gracias a gradientes químicos. Cuanto mayor sea la diferencia de concentraciones, más facilidad de paso. -Moléculas con carga: Se pueden mover por gradientes electroquímicos (más fuerza), gradientes eléctricos. Favorecen la entrada de cationes en la célula. Si quiero que ese transporte vaya más lento. Emplearé inhibidores o quitaré transportadores. TIPOS DE TRANSPORTES 1) PASIVO: no hay gasto de ATP y va a favor de gradiente. 1.1) Difusión simple: no existe una proteína directamente implicada en el transporte. Ej.: oxígeno, dióxido de carbono (se cuelan entre los huecos) y poros y canales iónicos (abren huecos. 1.2) Difusión facilitada: es necesario una proteína que se implique. Ej.: permeasas con la glucosa. 2) ACTIVO: hay gasto de ATP y va en contra de gradiente. 2.1) Bombas primarias (consumo ATP directo). Ej.: sodio-potasio-ATPasa. 2.2) Bombas secundarias (se aprovechan del ATP generado por las anteriores). Ej.: Na y Glucosa. 2.3) Transporte mediante vesículas (moléculas de mayor tamaño). TRANSPORTE PASIVO 1) Ósmosis (solvente). 2) Diálisis (soluto). 3) Difusión facilitada. Si la difusión es simple (1) y 2)) se establece una competencia entre la molécula y el agua, ya que si la molécula es soluble se va a dividir debido a la elevada constante dieléctrica del agua. Tiende a eliminar gradientes, aunque tiene impedimentos, debido a que los gradientes son fundamentales. 1) ÓSMOSIS Las paredes celulares de los vegetales también pasan por procesos de ósmosis, lo que pasa es que no son tan perjudiciales (están protegidos por la pared), de tal manera que: - Turgencia (medio hipotónico)= es el estado ideal de las vacuolas. Plasmólisis (medio hipertónico)= si no ha llegado a un punto crítico, se puede remediar si las volvemos a pasar a un medio hipotónico. Si no, empezarían a perder volumen, hasta separarse por completo de la pared. 2) DIÁLISIS Separamos las moléculas según su tasa de difusión. a. Moléculas pequeñas= permeables. b. Moléculas de gran tamaño= impermeables. *Presión osmótica: presión ejercida para frenar el flujo neto de agua. RESPUESTAS DE LOS ERITROCITOS A LOS CAMBIOS OSMOLARES Esta información mencionada anteriormente es útil para la separación de distintos tipos de moléculas. Ej.: Glucosa y albúmina. Si la Glucosa no se ha separado del todo de la álbumina y ya no hay más diálisis, la introducimos en otro medio. En esto se basa la hemodiálisis de enfermos renales: ·Si los cambios no son muy graves y se regresa a las condiciones normales, el eritrocito VIVE. I. II. III. Medio isotónico= Eri is fine. Medio hipértonico: Eri se dehidrata→ crenación= apoptosis. Medio hipotónico: Eri se hincha→ lisis= apoptosis. 3) DIFUSIÓN FACILITADA -Requiere de proteínas transportadoras (Permesasas). -Es un transporte pasivo= a favor de gradiente. Las fibras del citoesqueleto sobresalen= pinchar “el globo” -Es fundamental la formación de enlaces entre la sustancia transportada y el transportador. Las permeasas tienen uno o varios sitios de unión. La unión con el soluto es aleatoria. -Su función es acelerar el movimiento. Ej.: las permeasas introducen las moléculas de miles en miles. -Cambios de conformación reversibles. TIPOS DE TRANSPORTADORES ACTIVOS (BOMBAS) 1) Transportadores activos primarios= enzima+transporte. 2) Transportes activos secundarios= aprovecha el ATP que producen las bombas primarias. Siempre tiene dos puntos de unión (1 a favor de gradiente y otro en contra de gradiente). 3) Transportes activos lumínicos= aprovechan la energía que genera la luz. TIPOS DE TRANSPORTE ACTIVO Los transportadoes si mueven un solo soluto se denominan uniporte→ sitios de unión más específicos. Ya que solo transportan una sustancia. Siempre requiere de la acción de bombas. En el transporte activo, el movimiento se produce en contra de gradiente. Si los transportadores mueven dos cosas a la vez se produce un cotransporte que puede ser: Características de los transportadores 1) 1 o más sitios de unión (ATP, Na+, K+) 2) Uniones producen cambios conformacionales reversibles. 3) Al ser necesaria la unión, también se produce SATURACIÓN. Para una mayor velocidad, se añaden transportadores. 4) Las bombas son muy específicas. Esa especificidad se mide con Km. Inhibición de bombas (puede ser reversible o irreversible dependiendo de si las uniones son débiles o más estables (enlaces covalentes)). 1) Competitiva: posee la misma capacidad que el soluto para que ocupen su lugar. 2) No competitiva: se unen a un lugar distinto del sitio de unión, pero cambian la conformación de la bomba para que el soluto no pueda entrar. También se puede inhibir disminuyendo el número de transportadores. 1) Simporte: 2 entran/ 2 salen. 2) Antiporte: Las dos moléculas se mueven en sentidos opuestos. Ej.: sodio-potasio ATPasa. BOMBAS PRIMARIAS (multipaso) -Tipo P: Se llaman de tipo P porque durante el transporte se produce un proceso de fosforilación, cuando termina el transporte da lugar a una fosforilación. Se encargan de mover iones principalmente. Ej.: Sodio-potasio ATPasa. -Tipo F: funcionan como turbinas (es decir, mediante giros). Ej.: membrana mitocondrial interna ATPasa. • • Si giran en un un determinado sentido : giran en contra de gradiente→ hidrólisis de ATP. Si giran en sentido contrario: es como un canal iónico porque va a favor de gradiente. -Tipo V (vesícula): son de tipo P pero se situán en las membranas de orgánulos como por ejemplo los lisosomas. Todas las anteriores mueven iones principalmente. -Transportadores ABC (ATP-Binding-Cassette): a diferencia de las demás, bombean iones y moléculas pequeñas hidrófobicas. Lo que supone un gasto de 2 ATP en lugar de 1. BOMBAS SODIO-POTASIO-ATPASA -Consumo el 25% del ATP de la célula→ se gastan mucho en ella porque es importante. Funciones -Crean y mantienen gradientes (=actividad constante) que aportan la capacidad de crear señales eléctricas. Esto es crucial para la actividad de los nervios y de los músculos. -Transportes activos de moléculas→ Captación de Glucosa y aas. -Mantiene el pH en sus valores adecuados. -Regulan el equilibrio osmótico y el volumen celular. Funcionamiento 1) El ATP= crea sitios de unión para el sodio. 2) Se realiza la hidrólisis de un enlace P de alta energía= Fosforilación. 3) La fosforilación supone un cambio de forma de los sitios de unión (Na+ y K+). 4) Cuando se produce la unión del K+, se produce una rotura del enlace que había establecido la bomba con el fosfato, y el K+ deja de encajar. 5) Recuperación de la forma original Funciona todo el rato porque siempre hay mecanismos que intentan destruir los gradientes que ella construye. Por ejemplo: los canales iónicos que meten el Na+ y sacan el K+. Saber más… Las vacuolas pulsátiles pertenecen a organismos que viven en medios acuosos y que no tengan Na+-K+-ATPasa. Antes de que llegue toda esa agua al citosol, la vacuola se llena y expulsa H20 al medio extracelular (me contraigo). Características -El transporte que lleva a cabo es un transporte antiporte→ saca Na+ y mete K+. BOMBAS DE TIPO P (BOMBAS DE Ca+) -Es electrogénica, dado a que contribuye al potencial de membrana (saca 1 carga positiva). Esto supone que, cuando los solutos pasen a favor de gradiente, van a liberar energía. Encargadas de que se mantenga el gradiente de Ca2+, y que este se mantenga tan bajo en el citosol(10-7) y externo (10-3). En este caso cuando fosforilo la bomba, esta va a adquirir una conformación de tipo E1, que va a poseer gran afinidad con el Ca2+. Cuando se desfosforila adquiere una conformación E2, que posee un menor grado de afinidad con el calcio. -Permite la entrada de nutrientes (gracias a la energía que se genera con esos potenciales). -Ayuda a controlar el pH, regular la osmolaridad y la tonicidad. -Genera señal eléctrica (lo que supone una contracción del músculo). -Es una proteína de gran tamaño que encierra grandes cadenas polipeptídicas, en las que hay regiones destinadas a la unión del K+ (y también de la oabaína→ inhibidor competitivo) y del Na+ . Pueden estar situadas tanto en: - El retículo sarcoplástico (REL de células musculares (encargándose de introducir los iones en el interior del retículo, eliminándolos así del citosol). - La membrana citoplasmática (saca Ca2+ del medio intracelular). Este mecanismo se pone en funcionamiento en el caso de las células no musculares. Un aumento de la concentración de Ca+ manda una señal de contracción o de secreción (ejemplo: exocitosis de insulina). Por eso se le considera un segundo mensajero. Estos transportes secundarios van en contra de las acciones de la bomba más famosa pa´ la Luisa. Algunos ejemplos de este tipo de transporte son: Para mantener la concentración del catión calcio en el interior celular se emplean los siguientes transportadores: Gracias a la cadena respiratoria, la ATP sintetasa puede sintetizar ATP. Curiosidad: El cianuro mata porque impide la síntesis de ATP. Las bombas de iones en los lisosomas: permiten la disminución del pH (supone la hidrólisis de macromoléculas en monómeros). Para sintetizar estos monómeros hay que sacarlos fuera. 1) Mitocondria= introduce protones (H+) y cationes de calcio. Esto es importante de cara al Ciclo de Krebs. 2) Calmodulina= secuestra al calcio. 3) Transporte secundario= se aprovecha de las ganas de entrar del Na+, para sacar al Ca2+. A diferencia de las primarias, las secundarias siempre realizan un COTRANSPORTE, un soluto a favor y otro en contra. Esto supone que habrá veces que ese cotransporte sea antiporte y otras simporte. Las uniones (sobretodo en los simportes, son cooperativas, lo que significa que hasta que todos los sitios de unión no queden ocupados, no cambian de forma). Esta bomba está compuesta por 10 hélices transmembranales. TRANSPORTADORES ABC -Secretan los productos de desecho en medios acuosos. Por ello, se necesitan transportadores para eliminarlo. Son especialmente caros (gasto de 2 ATP). Están implicados en la resistencia farmacológica (MDR) (ej.: quimio// cloraquina (COVID-19)). Asimismo, también existen los CFRT (cuyas mutaciones pueden derivar en la aparición de fibrosis quística). Si este transportador no funciona, el cloro no puede ser expulsado de la célula, lo que hace que el agua tampoco salga. Esto supone un espesamiento de tejidos como la mucosa. TRANSPORTES ACTIVOS SECUNDARIOS/ ACOPLADOS Antes de que se pueda dar este tipo de transporte, es fundamental que se realice un transporte activo primario, para que esre cree un gradiente (mediante el gasto de ATP). Se aprovecha de la energía potencial (=Energía libre contenida en el gradiente). Con esta energía, impulsa a los solutos en contra de gradiente (introduciendo nutrientes→azúcares, aas, iones). - Ej.: Glucosa y sodio. Para introducir glucosa (en contra de gradiente), se aprovechan de las ganas de entrar del sodio. Este tipo de transporte es especialmente común con el sodio. Este tipo de transporte se emplea en células del epitelio intestinal No solo hay una copia de un tipo de transportador, sino que hay un gran número. Hay distintos tranportadores que mueven las mismas cosas pero con distintos métodos. Ej.: eritrocito -Salida de HCO3- y sale Cl-. → Sale CO2 TRANSPORTE MACROMOLÉCULAS GRANDES 1) Transporte vesicular= se empaqueta en una vesícula membranosa, esto es importante ya que este tipo de transportes dependen de la fusión entre membranas. Existen diversos tipos: - Exocitosis. Ej.: hormonas. Es fundamental en la secreción. Lo hace mediante vesículas, que permiten que las sustancias a excretar permanezcan el citosol. - Endocitosis. Se produce gracias a la necesidad de la célula de captar sustancias externas. Hay diversos tipos, dependiendo de si seleccionas más o menos y el tamaño: 1) Pinocitosis → líquido (no es nada selectivo, introduce el líquido con todo lo que en él está disuelto). 2) Endocitosis mediada por receptor→ extremadamente selectiva, porque lo primero que tiene que hacer es unirse a un receptor. Se suelen introducir macromoléculas. 3) Fagocitosis→ también hay que unirse a receptores de membrana, pero esta vez, las sustancias que se introducen son enormemente grandes (ej.: macrófagos). - Transcitosis: combinación de endo y exocitosis. EXOCITOSIS REGULACIÓN DEL PH Las sustancias exocitadas suelen ser sustancias que han sido sintetizadas por el RER o el REL, pasan al Golgi para su maduración y los introduce en vesículas para que sean excretados al exterior. Para un pH óptimo se requiere de antiportes. ENDOCITOSIS -pH bajo/ respuesta ante una acidosis puede ser - Cotransporte de tipo antiporte: Entra Na+ y salen H+. Al producir invaginaciones rodea a las sustancias que la célula quiere meter en su interior. Lo que permite que dos trozos de membrana se aproximen, lo que a su vez supone una fusión de membrana, formándose una vesícula. -Cotransporte de tipo antiporte: sale Cl- y entra HCO3-. Tipos: -pH alto/ alcalosis -Pinocitosis: se produce una invaginación de la membrana, así todo el líquido del exterior queda dentro de la vesícula cuando se forma. El líquido suele salir de la vesícula membranosa mediante algunos de los transportes vistos En este caso el eritrocito introduce cloro en la célula y saca HCO3-. -Salen Na+ y entran H+. anteriormente. En el caso de la transcitosis de glucosa del plasma, se emplea la pinocitosis ya que es la más eficaz. -Endocitosis mediada por receptor: una vez encaje con el receptor, se produce una vesícula de endocitosis o endosoma. Para salir del interior de estas vesículas, suelen introducirse en el lisosoma. En nuestras células este es el mecanismo más eficaz para captar nutrientes y lípidos. Ej.: captación de las lipoproteínas (VLDL, LDL (malo) y HDL→ cuanta más densidad =más proteína y viceversa). Son una especie de micelas de fosfolípidos en cuyo exterior están las proteínas y en cuyo interior están los triglicéridos y ésteres de colesterol (emplean las lipoproteínas como transportadores para sustancias muy poco polares→ en su interior caben miles de moléculas de colesterol). La cubierta suele estar formada por 500 moléculas de colesterol y 800 fosfolípidos. Como se utilizan receptores (suelen estar en dominios concretos, formados por las proteínas unidas a la parte interna de la membrana por un lado y a los receptores por otro, esta proteína es la clatrina→ ayuda a formar dominios y hace que la membrana cambie de forma (siempre la mantiene un poco curvada para dentro), ya que recubre la parte interna de la membrana) para introducir el ligando, el proceso es muy específico. A medida que los receptores se van uniendo a las lipoproteínas, la curvatura es cada vez más pronunciada. A medida que se va curvando, se va formando la vesícula recubierta (porque la membrana sigue rodeada de clatrina). Las bombas de protones de tipo V, van a empezar a disminuir el pH de los endosomas, rompiendo los enlaces débiles entre los receptores y las sustancias que has introducido. Esto genera que todos los receptores se queden en una parte de la vesícula, separando los receptores de las moléculas de LDL. Dando lugar a dos vesículas: endosoma secundario (con LDL, transformando estas macromoléculas en moléculas más sencillas→ esas moléculas saldrán (mediante transportadores con posterioridad al citosol para ser utilizadas en diversos procesos celulares) y vesículas con receptores (esta vesícula se fusiona con la zona de la membrana citoplasmática que contiene clatrina). En las células en las que no haya clatrina, nos encontraremos con la caveolina, que recubren el exterior de la membrana celular. *La clatrina tiene forma de trisquelion→ la unión de esos trisqueliones es la que le aporta a la clatrina esa forma hexagonal. Para enviar mensajes desde el Golgi a los lisosomas también se emplea la clatrina. Hay veces que los excesos de colesterol son debidos a la hipercolesterolemia familiar, lo que supone que mis células no captan bien el colesterol y este se queda en la sangre. Este trastorno puede ser debido a: 1) Los receptores no se unen a la clatrina y la membrana no se invagina. 2) Afecta a la zona de unión del LDL con el receptor. FAGOCITOSIS Tras la unión a sus receptores (esto provoca el crecimiento de los filamentos de actina creando pseudópodos, que van siendo cada vez más largos, hasta que los dos extremos chocan y forman una vesícula) forman EVAGINACIONES que rodean los que hay en el exterior de la célula. En este caso, a las vesículas se las llama vesículas de fagocitosis o fagosomas (también existen fagolisosomas, cuando se unen a los lisosomas, los cuales poseen hidrolasas ácidas). La fagocitosis posee diversas funciones 1) Nutrición en organismos unicelulares 2) Protección frente a invasores (macrófagos y neutrófilos)→ células del sistema inmune, eliminando todo lo potencialmente peligroso. También se pueden aprovechar de lo que fagocitan, pero no es su principal función. Los macrófagos también eliminan células dañadas o viejas y sustancias de desecho (ya que son los que poseen más variedad de enzimas para llevar a cabo la destrucción de esas sustancias) En ambos casos las enzimas lisosomales entran en contanto con las sustancias que han entrado a la célula, convirtiéndolas en monómeros. Esta digestión convierte esas grandes estructuras en estructuras de menor tamaño para ganar energía, ... INTERACCIONES ENTRE CÉLULAS; UNIONES INTERCELULARES Los tipos de uniones son: 1) Uniones de anclaje (célula-célula o célula-matriz). Son las únicas en las que podemos tener ambas uniones. 2) Uniones de oclusión: son las únicas en las que las dos membranas están físicamente unidas. Es decir, se forman 2 compartimentos aislados. 3) Uniones de hendidura: se encargan de la formación de “canales” para transmitirles sustancias a sus vecinas. 4) Uniones sinápticas: uniones en las cuales se liberan mensajes. ESTRATEGIAS ARQUITECTÓNICAS Existen dos tipos de tejidos según las cosas a las que se unan las células: 1) Células muy separadas→ matriz extracelular es fundamental. 2) Células muy juntas→ uniones intercelulares. Ej.: epitelios. Lo que supone que tienen muy poca matriz extracelular. El citoesqueleto es el que se encarga de transmitir esa presión de una célula a otra. UNIONES DE ANCLAJE También llamadas uniones de adhesión (adherens). Unidas a los filamentos del citoesqueleto. Que los filamentos del citoesqueleto se unan, supone la posibilidad de que se transmita el estrés. UNIONES DE OCLUSIÓN Existen tejidos en los cuales las células se encuentran muy separadas unas de otras, lo que hace imposible que se unan membrana con membrana. Pero todas las células tienen que estar unidas con lo que hay fuera. En este tipo de tejidos, las uniones se llevan a cabo con la matriz extracelular y permiten a las células enviar y recibir mensajes, adaptarse al medio, saber lo que hay que hacer, … También se las denomina ocludens/ estancas/ estrechas. Se emplean solo para las uniones célula-célula. No existe el espacio intercelular. Esto crea barreras de permeabilidad→ supone que solo puedes pasar de un compartimento a otro si esa célula tiene un transportador para esa sustancia. Las uniones entre las células determinan la arquitectura corporal (forma, mensajes, disposición de las células, resistencia mecánica …). La orientación de los orgánulos también está definida por las uniones (ej.: núcleo de las células intestinales), también contribuye a la determinación de la forma de desplazarse (esto lo que significa es que en algunas células, las uniones se van a romper de forma más sencilla para que esta se pueda desplazar). También denominadas de comunicación/ GAP/ de hendidura (generan un acercamiento de las dos membranas en las zonas en las que hay unión)/ plasmodesmos. Solo se emplean entre uniones célula-célula. Ponen en contacto el citplasma de células adyacentes. También van a poner en contacto sus citoesqueletos. A veces, esas uniones también ayudan a que el estrés (peso) se transmita. Para soportar mayor estrés, se produce una transmisión mecánica (las proteínas de membrana se unen a la matriz y/o al citoesqueleto). Ya que estos últimos, están preparados para recibir ese estrés mecánico. A veces, cuando las células están unidas unas con otras muy juntas, ese estrés mecánico se lo pasa a la otra. UNIONES QUE FORMAN CANALES Normalmente las céluas unidas a otras células suelen tener los tres tipos de unión. TIPOS DE UNIONES INTERCELULARES 2) Uniones de anclaje para filamentos de actina: encargados de uniones célula-célula y célula-matriz. Los filamentos de actina siempre que los unamos a miosina pueden dar lugar a estructuras contráctiles. A cada una de estas uniones se le va a dar un nombre propio. 1) Uniones a filamentos de actina 1.1) Unión célula-célula (adherentes)→ son zónulas que reciben el nombre de desmosomas en banda o en cinturón. 1.2) Unión de célula-matriz (adeherentes)→ son máculas que reciben el nombre de contactos focales. 2) Uniones a filamentoss intermedios: 2.1) célula-célula: desmosomas (mácula). 2.2.) célula-matriz: hemidesmosoma (mácula). -Conectan el citoesqueleto de células adyacentes entre sí (de tal manera que una célula se entera de lo que le pasa a la otra, se transmite el estrés). Cuando hablamos de uniones se emplean los términos zónulas (ocupa una gran superficie de la membrana) y máculas (ocupan menos zonas de las células). No nos da una idea del tipo de unión. -Complejo de unión: situación en la que dos células están unidas por varios tipos de unión diferentes. En esa estructura los diversos tipos de unión están siempre colocados de una misma manera. Esta estructura se mantiene porque es útil. El complejo de unión señalado en el dibujo está estructurado de la siguiente manera (uniones estrechas, uniones adherens y desmosomas)→ separan el dominio apical del basal. Los dos más importantes están en el epitelio intestinal y en el músculo cardíaco (discos intercalares). Son distintos entre sí porque tienen diversas funciones. UNIONES DE ANCLAJE O DE ADHESIÓN -Reciben este nombre porque algunos de los filamentos intermedios del citoesqueleto o de actina (nunca los dos) se unen a ellas:1) Uniones de anclaje para filamentos intermedios: encargados de uniones célula-célula y célula-matriz. Nunca van a ser contráctiles, pero son muy resistentes al estrés mecánico. -Conectan el citoesqueleto con la matriz (es decir, con las proteínas de la matriz extracelular→ colágeno). Esto sirve para que las células de los tejidos no tengan que soportar ellas solas el estrés mecánico (=resistencia mecánica). Es decir, forman láminas resistentes (piel, músculo cardíaco). Aportan resistencia mecánica. Formación de este tipo de uniones Se forman mediante proteínas transmembranales de unión que sobresalen de la membrana. Para que esas dos células se mantengan unidas, son necesarias las proteínas de la placa densa. Estas últimas, están unidas los filamentos intermedios o de actina. Los filamentos intermedios pueden estar formados por distintas proteínas dependiendo de a qué tejido pertenece esa célula. Ej.: filamentos de queratina (epitelio), filamentos de desmina (musculares) y neurofilamentos (neuronas). PROTEÍNAS DE ADHESIÓN CELULAR Son fundamentales para las uniones de anclaje y se originan en la placa densa. Tienen distintas formas dependiendo de si las uniones son célula-célula (cadherinas) o célula-matriz (integrinas). Siempre es así, salvo en excepciones contadas. Hablamos de integrinas y cadherinas en plural porque pertenecen a una misma familia de proteínas (proteínas muy similares en forma y funcionamiento pero con secuencias de aa no idénticas→ a veces para distinguirlas se les añade un número o una letra). Todas tienen características comunes - Se originan en la placa densa. Atraviesan la membrana. Interaccionan con las de la célula adyacente o la de la matriz extracelular. Se unen al citoesqueleto celular. 2) Sin calcio se pliega sobre sí misma (no genera uniones). Las dos células están unidas muy fuertemente (estables y duraderas) debido a la gran superficie de unión. Sin embargo, como están formadas por enlaces débiles, no resulta tan difícil romperlas (se rompen de forma secuencial→ cremallera). Esto supone que para separar esas células no necesito gran cantidad de energía (las rompo poco a poco). Muchos de esos enlaces débiles se sitúan en paralelo. INTEGRINAS Poseen dos conformaciones posibles (activa e inactiva). Las uniones de las integrinas son uniones que duran menos tiempo (transitorias) que las anteriores porque son más fáciles de romper. Por eso, se emplean en la formación de hemidesmosomas= desplazamientos celulares. Ej.: ameba. Dependiendo del tipo de célula que seas, las cadherinas e integrinas de la membrana van a ser de un tipo u otro. Esto determina con quién te puedes unir y con quién no. CADHERINAS Participan en uniones adherentes o de adhesión y que necesitan estar unidas a Ca2+. Pueden adquirir 2 conformaciones: Se puede observar el tipo de cadherina que es, según las uniones que forme (mismo tipo de cadherina o compatibles). Estas uniones adherens permiten que todas las células del embrión se mantengan en fila formando el ectodermo. Cuando las células de en medio de la fila responden a una orden de contracción, las uniones se contraen, formando una invaginación. Finalmente, la estructura se cierra. Asimismo, existirá otra orden que comunique que las que se han cerrado tengan unas cadherinas diferentes a las que tienen las del ectodermo, así se separan porque no son compatibles. Creando el tubo neural por un lado y el ectodermo por otro. UNIONES ADHERENS 1) Con calcio, está extendida, lo que permite la formación de enlaces (mirar imagen). Las uniones también sirven para modelar la forma de las células y hacer que cambien todas a la vez, funcionando de manera síncrona. Esto es fundamental gracias a las uniones adherentes, especialmente aquellas en las que participa la actina. En algunos movimientos (corazón, movimientos peristálticos del intestino) es fundamental. UNIONES DE CÉLULAS CON MATRIZ EXTRACELULAR DESMOSOMAS (mácula) Se anclan filamentos intermedios y son uniones de célula con célula. Estas uniones no se emplean para coordinar movimientos, sino para que esas células soporten mejor el estrés mecánico. Por eso estas uniones son muy abundantes en los epitelios (epidermis→ barrera mecánica de protección) y en los músculos. Estas uniones generan un armazón estructural, de manera que da lugar a una estructura en la que el citoesqueleto de todas las células está comunicado. Se pueden repartir el estrés mecánico gracias a esas comunicaciones. Asimismo, parte del citoesqueleto de la célula está unido con la matriz extracelular. Entre células, las comunicaciones las llevan a cabo los desmosomas y la unión célula-matriz con los hemidesmosomas. INTEGRINAS MATRIZ EXTRACELULAR Siempre que algunas de esas proteínas tengan un fallo va a existir un riesgo que provoque la ruptura de las uniones. Ej.: piel→ epidermis formada por muchas células unidas unas con otras mediante desmosomas, debajo está situada la dermis que es un tejido conjuntivo al que las células se unen mediante hemidesmosomas. Debido al roce con un zapato, se rompen los hemidesmosomas (la epidermis se separa de la dermis) y para solucionarlo, el sistema inmune manda un líquido para arreglarlo (ampollas). Si la situación sigue así, no llegan nutrientes a la epidermis y esta muere. Esto es un riesgo en caso de que la zona dañada sea muy grande, ya que no hay control de electrolitos ni del líquido y las bacterias pueden entrar y salir cuando quieran. Hay diversos tipos de enfermedades bulllosas: 1) Pénfigo: enfermedades autoinmunes de origen genético que generan anticuerpos contra las cadherinas que las destruye. Permite que la célula responda los diferentes estímulos que llegan a ella. Aunque también sirve para un aumento de la resistencia mecánica. INTEGRINAS: ESTRUCTURA Hay más de 24 tipos y están formadas por heterodímeros. Las integrinas se unen con las proteínas de la matriz: laminina, fibronectina, colágeno, … La parte de fuera de la integrina se va a unir con el medio mediante enlaces débiles. También nos vamos a encontrar con uniones con el citoesqueleto. Las integrinas, como las cadherinas, también tienen dos conformaciones diferentes (plegada sobre sí misma→ NO sitios de unión; y estirada→ sitios de unión). Para activar o desactivar las integrinas se emplean reguladores alostéricos. Este mensajero puede ser intracelular o extracelular, se van a unir a sitios de la proteína que no sean los sitios de unión (son alostéricos, no competitivos). La integrina lo que permite es la unión indirecta de los filamentos intermedios y la lamina basal. Asimismo, hay otro tipo de transmisión de señales a parte de la química que es la mecánica (le pasa el peso a otras proteínas→ dependiendo de la dirección en la que va el peso, la célula puede cambiar de forma (si el peso va de la lámina basal hacia el citoesqueleto de la célula) (ej.: creación de pseudópodos). En algunas ocasiones, las respuestas químicas originan una señal mecánica o viceversa. Además es bidireccional (puede ser de la matriz a la célula o al revés). HEMIDESMOSOMA Encargados de la unión de la célula (filamentos intermedios→ placa) + (ej.: colágeno XVII→ atraviesa membranas debido a que ayuda a las integrinas, es decir, se une también a las proteínas de la matriz extracelular) a la lámina basal. UNIONES CÉLULAS-MATRIZ DE ACTINA/ CONTACTO FOCAL Son uniones más pequeñas que los desmosomas, se denominan contactos focales. Por lo tanto las uniones son más débiles y se forman y se rompen sin apenas emplear energía. Este es el motivo por el que este tipo de uniones se emplean cuando se da desplazamiento. La actina que forma estas uniones también puede estar unida a miosina (denominados fibras de estrés= empujando a la membrana para que esa célula se adhiera a la superficie por la que se mueve). *Excepción: uniones mio-tendinosas→ van a estar formadas por un GRAN “contacto focal”. El gran tamaño de esta unión está justificado por la necesidad de que estas uniones sean resistentes. UNIONES ESTRECHAS Son abundantes en los epitelios pero también están presentes en los tejidos musculares. De este complejo va a depender la posición de la zona apical y la zona basal de los epitelios. Así, crean dos dominios. Es decir, polarizan la célula. Dividen la membrana en zonas con distinta composición química (diversos tipos de transportadores), distinta estructura (microvellosidades,…) y distinta función. Esto es debido a que una zona tiene que absorber y otra ha de secretar. Asimismo, el polo apical es una superficie libre, la membrana no está unida a nada. Sin embargo en la zona basolateral va a haber distintos tipos de uniones. Las uniones estancas sirven para crear regiones que no permiten la difusión libre de los solutos. Entre ellas no hay espacio posible, generan barreras. Divide a la célula en compartimentos que no están conectados de ninguna manera. Por eso se emplea la transcitosis. Es selectiva, porque solo se van a poder introducir en la célula solutos para los que exista un transportador. De estas células depende lo que desechamos y lo que aprovechamos. Las uniones de este tipo aparecen en distintos tipos de tejidos: -Células absortivas (renales, intestinales)→ captación de nutrientes y selección. Estas uniones polarizan a la célula→ la dividen en zonas de absorción y en zonas de secreción. Elimina los espacios intercelulares. La única manera de llegar al otro lado, es mediante los transportadores: 1) Transportadores de la superficie apical: ACTIVO (secundario) 2) Transportadores de la superficie basolateral: DIF. FACILITADA (permeasas). Ej.: transporte de Glucosa acompañada de sodio. Así la introduzco en contra de su gradiente. La impermeabilidad no es absoluta, esto es debido a que no siempre interesa que el sistema sea tan estricto. Estos estudios se llevan a cabo mediante colorantes. Ej.: Glucosa se tiene que introducir en el intestino muy rápido por los movimientos peristálticos. Esto supone que las proteínas cambian su conformación para dejar pasar a la glucosa. No es necesario gasto de E química, pasa a favor de gradiente. En cuanto el gradiente ha desaparecido, las proteínas se vuelven a cerrar y vuelve a llevarse a cabo la transcitosis, ya que las aperturas son temporales. Por esos huecos, solamente pueden pasar monómeros e iones. Cunado se abren esos huecos pequeños, se denomina transporte paracelular (ninguna célula participa de forma activa sin necesidad de estructuras celulares). Ej.: el vibrion cholerae, mantiene esos huecos siempre abiertos= PÉRDIDAS EXCESIVAS. PROTEÍNAS QUE FORMAN UNIONES ESTRECHAS -Las proteínas que forman esas uniones sobresalen muy poco de la membrana, por eso es casi una unión entre membranas. Se disponen en forma de hileras paralelas. Cada hilera está comunicada con las demás mediante el mismo tipo de uniones. Es decir, son zónulas. Hay varias filas para hacer más difícil que se rompan todas las uniones. Las proteínas que intervienen en esas uniones son las claudinas y las ocludinas, tienen una forma muy parecida. Son proteínas transmembranales de unión y son multipaso (atraviesas cuatro veces la membrana). Ambas son familias de proteínas, por ende, cada célula lleva un tipo diferente de las proteínas para que las células puedan llevar a cabo un reconocimiento (siempre se realizan uniones con las proteínas que son compatibles). Cada una va a permitir el paso de cosas distintas Ocludinas Aquí las proteínas que intervienen son también de dos tipos: conexinas e inexinas (unión entre conexinas y conexones). CANALES El diámetro no es más de 1,5nm (menos de 1000 Daltons). Por eso, pasan por ahí iones y moléculas pequeñas (monosacáridos, nucleótidos, inositol-3-P, AMPC, vitaminas y Ca2+). La coordinación se puede llevar a cabo mediante señales eléctricas o metabólicas (glucólisis, acumulación de lípidos, …) Tricelulina La proteína que mejor se conoce es la conexina: es multipaso (4 veces) y también pertenece a una familia. Son necesarias seis conexinas para formar un conexón, 2 conexones suponen un canal acuoso. Dependiendo de la familia de la que sea la conexina estos conexones pueden ser: homoméricos o heteroméricos. Los canales, debido al mismo motivo, pueden ser: homotípicos (suelen ser dos células del mismo tejido) o heterotípicos (se hace entre células que emplean conexinas distintas pero compatibles). Dependiendo del diámetro de los canales, los solutos que los atraviesen van a ser distintos. A las uniones formadas por la claudina y la ocludina a veces se les añade tricelulina. Esto sucede cuando hay que unir más de dos células. Se denominan canales porque el diámetro es pequeño y pueden estar abiertos o cerrados. Esto supone que necesitan una regulación de la apertura o el cierre. No están en todas las uniones estrechas, son prescindibles. Claudinas Hay hasta 24 tipos, lo que da lugar a muchos tipos de combinaciones. Hay varios tipos según su longitud. Cuanto más largas sean, más impermeables. También se pueden denominar uniones de hendidura porque forman un valle. UNIONES GAP/ DE COMUNICACIÓN/ DE HENDIDURA UNIONES GAP. FUNCIONES. La mayor parte de los tejidos las poseen, ya que son importantes para la comunicación intercelular. Es una comunicación directa. Crean una especie de túnel y canal que permite el paso de moléculas disueltas al citosol de la otra célula. Es una comunicación directa (citosol-citosol), esto garantiza que la señal llegue a su destino y que la señal esté protegida, es el transporte más rápido (más que la sinapsis) y es muy segura. Lo que provoca que ambas células hagan las cosas de manera síncrona. Además, supone un ahorro en “órdenes”. -En el músculo cardíaco o liso son fundamentales ya que transmiten el potencial de membrana y permiten que el movimiento se lleve a cabo de manera síncrona. (Células excitables eléctricamente). Ej.: músculo formado por 10.000 células, les doy una señal mediante una neurona motora (solo es necesario una porque gracias a las uniones GAP se pueden comunicar). Esto también supone que sea más fácil de coordinar. Al igual que las uniones de anclaje, hay un espacio intercelular (2-4nm→ permite el paso de iones). -En las células que no son excitables eléctricamente, su función la llevan a cabo mediante segundos mensajeros químicos como el calcio, los protones, el AMPc y el inositol. -Normalización del pH: mediante las uniones, si me estoy volviendo muy ácido, le voy mandando los protones a mis vecinas. Así, todas las células se igualan y otros mecanismos reguladores, terminan de normalizarlo. -Coordinación de actividades (ejemplo de antes). -Desarrollo de los folículos ováricos: los ovocitos están rodeados de células denominnadas células de la granulosa (dan órdenes al ovocito). Están unidos mediante este tipo de uniones. -Embriogénesis: cuando se genera el tubo neural, hay órdenes que se dan mediante uniones GAP. Sin embargo, cuando quiero separar el ectodermo, cierro los canales entre el resto de células y el ectodermo y, así, se produce la contracción de un grupo específico de células. REGULACIÓN DE LA PERMEABILIDAD -No pueden estar siempre abiertos. Esto se puede llevar a cabo mediante cambios en las conformaciones de las proteínas que se hacen muy deprisa y no necesitan ni enzimas ni energía. -Se cierran cuando el calcio es alto y el pH es muy bajo. Estos son señales de que el metabolismo no está bien regulado. Si no se puede arreglar mediante mecanismos de reparación, la célula entraría en apoptosis. Esto supone que la que tiene el calcio demasiado alto, enviaría el exceso de calcio a sus vecinas y todas entrarían en apoptosis. EL CITOESQUELETO El citosol es solo el líquido y el citoplasma lo es todo. Es importante comprender que en la célula TODO está en movimiento, y estos movimientos son azarosos. El citoesqueleto está formado por proteínas que son alargadas y estrechas (filamentos). Compuestos principalmente por microtúbulos, microfilamentos y filamentos intermedios. Estos van acompañados de proteínas accesorias que les permiten llevar a cabo su función. No todos los filamentos del citoesqueleto tienen ni el mismo grosor ni el tamaño, tampoco están distribuidos por la misma zona de la célula y no todos se organizan de la misma manera (se cruzan, paralelos, …). Esto nos permite averiguar el tipo y la función. Los ribosomas interaccionan con los filamentos del citoesqueleto. FUNCIONES DEL CITOESQUELETO -Mantiene la forma y tamaño de la célula→ Determina su forma y sus cambios. Es indispensable para que las neuronas tengan axones y dendritas, las intestinales tengan microvellosidades, haya cilios y flagelos y para la creación de pseudópodos e invaginaciones. -Fundamental para la organización del interior de la célula. Hay orgánulos que ocupan el sitio que ocupan debido a ello. Ej.: Aparato de Golgi→ cerca del núcleo. Tanto el Golgi como el RE como el núcleo, suelen tener una posición específica. Los que se mueven, también necesitan al citoesqueleto para moverse. -Interacciones mecánicas. Ej.: síntesis de proteína→ ARNm llega a su destino. Son carreteras. -Movimientos coordinados: es necesaria la coordinación de todo el citoesqueleto para que una célula pueda moverse. -Ayudan a sujetar el volumen celular para que la membrana pueda aguantar la presión de todo el líquido que hay en el interior. COMPONENTES DEL CITOESQUELETO -Microfilamentos/ filamentos de actina: formados por muchas unidades de una proteína globular denominada actina. Esas moléculas de actina se unen formando dos filamentos largos que se unen el uno al otro formando una especie de cuerda. Suelen estar por debajo de la membrana y por eso se dice que forman el córtex celular. Algunos se unen a proteínas de membrana. -Filamentos intermedios (más gruesos que los filamentos de actina pero más finos que los microtúbulos): están formados por proteínas fibrosas (hélice alfa). Los filamentos intermedios están formados por un tipo de proteína dependiendo de la célula a la que pertenecen. Suelen tener sus extremos cerca de la membrana. Recorren la célula de membrana a membrana hasta llegar a todas las partes de la célula. -Microtúbulos: son los de mayor diámetro (forman cilindros huecos). Están formados por la tubulina (heterodímero: alfa y beta). Todos se reparten radialmente por toda la célula desde el núcleo. Teniendo cada uno de ellos una distribución diferente. Los filamentos del citoesqueleto aparecen y desaparecen constantemente. Ej.: si no se hace nada, un microtúbulo desaparece cada 10 min, cuando uno muere otro llega. Esto supone que al nuevo lo puedo colocar donde quiera→ la célula cambia de forma de una manera muy rápida, lo que supone que te puedes adaptar a los cambios muy rápidamente. Ej.: función del huso mitótico en la división celular. Hago y deshago el huso mitótico. ORGANIZACIÓN DEL CITOESQUELETO EN DISTINTOS TIPOS DE CÉLULAS Los tres compuestos son igual de abundantes en el citoesqueleto. En las células normales (no polarizadas), las divisiones del citoesqueleto son muy parecidas. Puedo adivinar la localización de una membrana gracias a los filamentos de actina (debajo de la membrana). Sin embargo, en las células polarizadas, la distribución tiene que ser distinta: Intestinales -Los filamentos de actina: están debajo de la membrana y van a estar dentro de cada microvellosidad. -Filamentos intermedios: van de un desmosoma a un hemidesmosoma. Cruzan la célula unidos a la membrana. -Microtúbulos: se encuentran dispuestos verticalmente debido a las transcitosis (marcan el camino de las sustancias que entran y salen de la célula). Si se unen 13, forman las paredes del cilindro hueco (=microtúbulo). Cuando estos microtúbulos crecen se pueden polimerizar y cuando menguan, se despolimerizan. Los microtúbulos poseen polaridad. Las uniones para formar el heterodímero podría ser (cabeza-cabeza= hendidura con hendidura), (cola-cola= parte redonda con parte redonda) y (cabeza-cola= mix→ esto supone que un extremo tiene una forma y el otro, otra diferente). Esta última sería la disposición de los heterodímeros. Por eso se dice que tienen polaridad estructural. Neuronales -Filamentos intermedios: se encuentran en el axón, paralelos unos a otros. Las neuronas no se anclan a otras células. -Filamentos de actina: ayudan al mantenimiento de las formas de las dendritas. Se encuentran debajo de la membrana, formando el córtex. -Microtúbulos: dispuestos de manera paralela a lo largo del axón. También llegan a los extremos de las dendritas. Salen desde el soma. Marcan la salida de los neurotransmisores. Como son carreteras, han de tener un sentido. Este viene marcado por sus extremos. Células en división→ la actina es fundamental para la cinocitosis y los filamentos intermedios se están creando. MICROTÚBULOS Si se ven al microscopio aumentado, se ven como si fueran rectángulos. DIAPO: sitio verde al lado del núcleo es en el que empiezan los microtúbulos. Al ser cilindros huecos, son los más rígidos. Como se encuentran cerca del final de la membrana participan en la forma de la célula, participan en ciertos tipos de desplazamientos celulares (siempre que se hagan con cilios o flagelos), formación de estructuras como los centriolos, son imprescindibles para formar el huso mitótico (división celular). DIAPO: Imagen III→ representa la función de organización. Estructura: cilindro hueco (que sea hueco le aporta rigidez)→ (24nm). Compuestos por tubulina (heterodímero alfa-beta→ cuando se polimeriza o se alarga lo hace por la unión de los heterodímeros). Hay un extremo alfa-terminal y beta-terminal. En las uniones laterales, se unen alfa con alfa y beta con beta. La unión de heterodímeros se realiza mediante enlaces débiles (como son muchos=resistencia. Es muy fácil quitar y poner heterodímeros de un protofilamentos, pero no del microtúbulo). Para formar un heterodímero necesito la unión de las cadenas alfabeta de forma discontinua. A estas uniones se las denomina protofilamentos. Para que un microtúbulo se pueda alargar, la subunidad alfa y beta han de estar unidas a una molécula de GTP. Para que los heterodímeros se puedan separa de ahí, hay que liberar energía (=hidrólisis de GTP). Ambos tienen GTP, para separarse del filamento, tienen que llevar a cabo pequeños cambios conformacionales. Sin embargo, solo la beta lo va a poder hacer (sin mucha complicación), ya que no estaría unida a nada. La que lo va a poder hacer de manera más sencilla es la beta-terminal, aunque las beta intermedias también lo van a poder hacer (van a hacerlo de forma más lenta). Los extremos crecen y se alargan de forma distinta (extremo beta va a ser más rápido= extremo más; el extremo alfa va a ser extremo menos). A esto se le denomina polaridad funcional. De esta manera, tanto la forma como el comportamiento, nos indican el sentido de la molécula. del anillo), esto supone que cada vez que pasa una alfa-beta la atrapa, atrapa a las moléculas de alfa-tubulina. En condiciones normales van a estar continuamente formando y deshaciéndose si no hacemos nada. La vida media de un microtúbulo son diez minutos (ventajas para cambios de forma rápidos de la célula). Controla el número de microtúbulos, la localización, la orientación y la polimerización. Los extremos más suelen estar cerca de la membrana, y el extremo menos está más cerca del núcleo. Está formado por microtúbulos. Son estructuras cilíndricas cuya estructura está formada por 9 tripletes de microtúbulos. Los centriolos están es posición perpendicular el uno respecto al otro. Cuando se replican para que la célula se divida, emplean una división semiconservativa. Cuando los heterodímeros hidrolizan el GTP (subunidad beta), empiezan a disminuir su tamaño y, por eso, adquieren forma de palmera. Los heterodímeros son reutilizables. Si las condiciones del medio lo permiten, se puede volver a formar el microtúbulo. Los microtúbulos se forman en los centrosomas (COMT). Son imprescindibles para la formación de los microtúbulos, aunque todavía no se sabe cómo. Dependiendo de la célula los COMT pueden estar dispuestos de diversas maneras. Siempre están orientados de la misma manera (extremo más→ memb. Citoplasmática). Ej.: mitocondria que va hacia la membrana= va hacia más. En las células que van a empezar a dividirse, los COMT se replican y cada uno de ellos está en un polo del huso mitótico. Extremo más y menos igual que antes. En las células que tienen cilios o flagelos: van a tener en la base un corpúsculo basal que es un único centriolo (COMT). Los microtúbulos van a crear un esqueleto interno para soportar la membrana del cilio o del flagelo. Es importante que los microtúbulos sean muy dinámicos (huso mitótico). En el caso de los flagelos me interesa que no se muevan tanto, sino que duren más tiempo. Por eso, se convierte a los microtúbulos en estructuras más estables, para ello se requiere la ayuda de las proteínas accesorias o asociadas a los microtúbulos. Las células ciliadas (en nuestro caso), no se mueven ellas, mueven lo que hay fuera (corrientes). También fijan los orgánulos en la célula. Ejemplo: aparato de Golgi, núcleo y RE. Aquellos orgánulos que no tienen un sitio fijo, se mueven mediante movimientos dirigidos por los microtúbulos (carreteras). CENTRIOLOS Cuando terminan de duplicarse, se mueven hacia los extremos de la célula, y empiezan a formar el huso mitótico. Los centriolos no son los corpúsculos basales de los cilios y los flagelos. Para saber qué van a formar los microtúbulos, nos centramos en la localización→ cerca de la membrana= cilio o flagelos. Cerca del núcleo→ huso mitótico. PROTEÍNAS ACCESORIAS DE LOS MT (MAP/ PAM) Todos los cambios de comportamiento los llevan a cabo mediante señales muy estrictas. Hay 100 tipos diferentes que cambian el comportamiento de los microtúbulos de distintas maneras. Ej.: formación invaginaciones o evaginaciones si microtúbulos cerca de la membrana. Proteínas que determinan el dinamismo de los MT: -Proteínas que favorecen la nucleación (formación de nuevos microtúbulos) y su estabilización: 1) Unión a extremos (CAPs)→ tapan extremos, no se pueden unir. 2) Estabilización de los laterales→ protección frente a la catanina. -Proteínas que desestabilizan pueden 1) Unirse a los extremos y se despolimeriza→ desensamblaje 2) Cortes (catanina)→ fragmentación. Es importante en la anafase FORMACIÓN DE MICORTÚBULOS -Proteínas que forman enlaces cruzados o estructuras complejas El COMT además de tener el par de centriolos, también tienen el material pericentriolar (albergan heterodímeros para crear los microtúbulos, así como el GTP). La gamma-tubulina está representada como anillos (alrededor de 50→mirar diapositivas). Estos anillos son lugares de nucleación (unión de heterodímeros para formar un microtúbulo). Para esto, la gamma-tubulina forma anillos en los cuales hay 13 moléculas de gamma-tubulina (laterales 1) Obliga a los microtúbulos a permanecer paralelos (paralelos formando un cilindro→ haces; paralelos formando hojas→ láminas) 2) Obliga a los MT a cruzarse→ redes -Proteínas motoras: transporta cosas de gran tamaño a un sitio concreto, para ello andan sobre microtúbulos. SUSTANCIAS QUE AFECTAN A LOS MT - Taxol= se une y estabiliza los microtúbulos. Nocodazole= se une a los heterodímeros para evitar que se alarguen los MT. Esto es fundamental en la quimioterapia, ya que las células tumorales se dividen sin control. No solo afectan a las células tumorales, sino que afectan a todas las células en división del organismo (médula ósea, pelo, …) Ej.: caveolas o depresiones recubiertas para hacer endocitosis y pinocitosis. Si se produce exocitosis pueden ser pseudópodos (la forma del pseudópodo varía según lo que envuelvan→ importantes para la fagocitosis y para los movimientos ameboides), microvellosidades (proyecciones digitiformes), filopodios (prolongaciones con forma de espina→ empleados para el estudio del terreno de las células, es decir, la detección de la orientación de las señales. Ej.: importantes en las neuronas), lamelipodios (forma de abanico→ importantes en los movimientos ameboides), … Todo lo relacionado con estrucutras contráctiles (sarcómeros). Siempre tienen que estar asociados a la miosina para que sean estructuras contráctiles. FILAMENTOS DE ACTINA PROTEÍNAS ACCESORIAS DE LOS MF Las moléculas de actina se unen para formar dos protofilamentos y estos se enrollan, formando una especie de cuerda. Son mucho más flexibles que un microtúbulo, aunque son menos resistentes. La mayoría de los filamentos de actina están debajo de la membrana citoplasmática, por eso se dice que forman el córtex celular. Son los más directamente relacionados con las proteínas de la membrana. Tienen un papel fundamental en la forma de la célula, son los responsables de los cambios de forma (implicados en la formación de las microvellosidades→ más membrana= más transportadores). Algunos de esos cambios de forma son fundamentales para que la célula se pueda desplazar mediante mov. ameboides. Si la actina se junta con la miosina, se forma una estructura contráctil (son indispensables para los movimientos ameboides). También son fundamentales para la citocinesis (anillo contráctil= actina). En las fibras musculares, hay grandes cantidades de actina y miosina. Los MF funcionan gracias a proteínas accesorias. Unión de actina más miosina= zona más oscura. Solo actina= zona más clara. Dependiendo de cómo se encuentre la actina en una célula puede ser: -Actina G: 1 cadena polipeptídica globular con un sitio de unión al ATP/ ADP. - - Proteínas de nucleación: 1) Formina (ayuda a la formación de MF). 2) Cap Z (más estable)→ fundamental en los sarcómeros. 3) Tropomiosina→ estabiliza a los filamentos mediante el lateral→ sarcómeros. Proteínas desestabilizadoras Proteínas que les obligan a formar enlaces cruzados: 1) Filamina→ formación de redes. 2) Espectrina→ les obligan a unirse a la membrana. Imp.: eritrocitos. PROTEÍNAS ACCESORIAS PARA FORMAR ESTRUCTURAS COMPLEJAS Filamentos de actina unidos directamente a las proteínas de la membrana o mediante las proteínas accesorias. Esto supone que las uniones de filamentos de actina con la membrana, aporta integridad mecánica. Los cambios de forma dependen de la finalidad (ej.: microvellosidades, filopodios, pseudópodos)→ estas estructuras están formadas por proteínas que crean haces de fibras se denominan Bundling proteins (distinta separación en función de la célula). -Actina F: forma el filamento de actina. Forman una hélice dextrógira de gran flexibilidad. Para formar los protofilamentos de actina, todos los monómeros tienen que tener ATP. Siguen teniendo polaridad estructural y funcional (al igual que los MT). Al tener polaridad, estas estructuras indican también orientación (=carreteras). Requieren de interacción con proteínas accesorias para transportar las cargas. El extremo más es el que está más cerca de la membrana y el extremo menos es el que está más lejos. Para formar un lamelipolios, necesito Gel-forming proteins, crean redes (dependiendo de la apertura de la bisagra los agujeros van a tener una mayor o menor tamaño). Todo lo que tenga que ver con la forma de la célula está relacionado con los filamentos de actina. Lo que permite el mantenimiento o el cambio de forma (dependiendo de las necesidades celulares). *Para que una fibra sea contráctil, los filamentos tienen que estar dispuestos de forma antiparalela y, con el sufiente espacio para que quepa la miosina. La variación del estado del citosol se “determina” en la zona del córtex: cuanto más densas sean las uniones de la gel-forming proteins, va a ser más gel. Las células que no están en división son más bien sol. BUNDLING PROTEINS Estas proteínas mantienen a las microvellosidades estables y permite que sean paralelos y estén situados de forma vertical. Cada proteína va a estar unida a dos filamentos de actina diferentes, dependiendo de las proteínas que se empleen, la forma que se adoptará para mantener esa forma celular será distinta. En función de la proteína empleada, la distancia que aporten los microfilamentos entre, por ejemplo, microvellosidades va a ser mayor o menor. En las microvellosidades me interesa que todos las uniones entre los filamentos de actina sean más bien cortas para que se pueda disminuir la distancia. En las microvellosidades, se emplean la fimbrina y la villina→ no dejan apenas espacio entre los filamentos de actina, la miosina II no cabe. En estructuras con capacidad contráctil: hay una mayor separación entre los filamentos de actina, para poder introducir la miosina II. Quien une los filamentos de actina a la membrana es la miosina I. GEL-FORMING PROTEINS Cuando quiero formar estructuras encargadas del movimiento, se emplean proteínas con enlaces cruzados que forman redes. Las proteínas tienen dos sitios de unión a la actina, obligando el cruce de los filamentos de actina. Es decir, cada una de las proteínas verdes (mirar diapo) se va a encargar de cruzar los sitios de unión. A este tipo de proteínas se las denomina las Gel-forming proteins. Ejemplos de estas proteínas son filamina (lamelipodios) y espectrina (eritrocito). Esta consistencia más viscosa, facilita el movimiento de las membranas. APLICACIONES MÉDICAS Ejemplo diapo A: no se pueden desplazar porque, aunque lo intenten, no pueden formar lamelipodios. Dato curioso: la seta de los gnomos es venenosa porque afecta a los filamentos de actina y se dejan de llevar a cabo funciones vitales básicas. FILAMENTOS INTERMEDIOS Son las estructuras más estables y menos dinámicas del citoesqueleto pero, son muy resistentes a las fuerzas mecánicas. Son buenos para que las células transmitan el estrés mecánico a sus células vecinas o a la matriz extracelular. Están formados por proteínas fibrosas no globulares, cuya estructura secuandaria es la hélicealfa. Dependiendo de la célula que sea el filamento intermedio va a ser distinto, pero todos tienen estructura secundaria similar. Siempre se unen a desmosomas o a hemidesmosomas, están distribuidas de tal manera que sus extremos siempre están anclados a la pared (placa densa). Comunican el citoesqueleto con la matriz extracelular. Por esto, son muy abundantes en los tejidos que están sometidos a gran presión: epitelios (queratina), músculo (desmina), tejido conjuntivo, es decir, fibroblastos (vimentina) y neuronas (neurofilamentos)→ aunque los axones sean pequeños, aguantan esta presión por los filamentos. No todas las proteínas forman el mismo número de enlaces cruzados, ya que depende de la resistencia que tenga que tener la célula. Cuantas más uniones, más fuerte es. En el núcleo también existen filamentos intermedios, pero siempre están debajo de la membrana nuclear, que forman estructuras planas sobre la cual se apoya la envuelta nuclear. Como esta unión de filamentos intermedios es tan fina, se la llama lámina nuclear.Están proteínas, que forman las láminas nucleares, se denominan láminas (solo aparecen dentro de los núcleos de las células). Los filamentos intermedios duran más que los anteriores y son más estables (no se alargan o contraen). Estos filamentos, tampoco tienen polaridad. Se forman de otra manera. En las células que se mueven, se puede apreciar en qué dirección se mueven por la zona más ancha de la célula. Estos movimientos, que son ameboides, son necesarios, pero a veces no me interesan (ej.: células tumorales→ metástasis). Evitando que las células tumorales se muevan, puede cesar esa metástasis. Para ello se proveen medicamentos como la faloidina (estabiliza filamentos, no avanza), citocalasina (estabiliza extremos +), swinolida (cortes de actina), latrunculina (impide su polimerización). Los extremos, tienen estructura globular. El resto del filamento se une de una forma paralela (extremo amino con su análogo y veceversa)= formación de dímeros. Este dímero se asocia con otro dímero (se unen de una manera solapada). En un lado sobresale el amino terminal (caperuzas) y está más metido el carboxilo y en el otro dímero al revés. Por eso se dice que este tetrámero es antiparalelo. Se unen dos tetrámero, formando un octámero (se encajan por las partes más y menos salidas→ puzzle). Este proceso sigue hasta retorcerse y cerrarse, formando una cuerda, de tal manera que desde el principio va a adquirir su longitud final. Los enlaces son muchos más que en los microfilamentos y los microtúbulos. Como afectan a los filamentos de actina, también afectan a loa división celular (anillo contráctil). El número de tetrámeros que emplees para formar la pared, varía según el tipo de filamento que sea. DIAPO (FILAMENTOS INTERMEDIOS) Las zonas en las que los filamentos verdes se alargan, son los desmosomas. Es una célula epitelial y, por ello, tiene un gran número de desmosomas para poder repartirse mejor el estrés mecánico. -Esclerosis Lateral Amiotrófica (ELA)= Enfermedad de Lou Gehrig: Los filamentos intermedios forman axones permitiendo el paso de neurotransmisores (en las placas motoras, por ejemplo: se libera acetilcolina). Para que la conexión entre axones vaya bien, es necesario que los filamentos estén paralelos. Lo que provoca esta enfermedad es que no deja pasar los neurotransmisores (desorganización de los filamentos intermedios→ se cruzan). Esto provoca que la neurona que tuviera que recibir señales, se atrofian. LÁMINA NUCLEAR Son filamentos intermedios que forman una red plana en forma de hoja que forman una red plana debajo de la envuelta nuclear. FILAMENTOS INTERMEDIOS: DISPOSICIÓN Aunque los enlaces sean débiles, hay una gran cantidad y, por ello, no son fáciles de romper. En este caso también existen porteínas accesorias pero su función es hacerlos aún más resistentes. También emplean gran cantidad de proteínas que les ayuden a formar uniones cruzadas para formar haces o redes. Como no son dinámicos, no pueden generar movimiento pero, cuando la célula se mueva, los filamentos intermedios se tienen que reorganizar. Para ello, existen proteínas que ayudan a los filamentos intermedios a unirse a microtúbulos y a filamentos de actina. Cuando se habla de que forman redes resistentes, se hablan de redes de comunicación (desmosomas). Es decir, son redes que interconectan, permitiendo el paso de tensiones de un lado a otro. ALTERACIONES DE FILAMENTOS INTERMEDIOS -Si una mutación afecta a una de las queratinas encargadas de la formación de filamentos intermedios, van a provocar que los filamentos intermedios sean menos resistentes de lo que deberían ser. Esto provoca la separación de los desmosomas y los hemidesmosomas, generando la separación de la dermis y la epidermis→ Epidermolisis bullosa. También se puede ver dañado el tracto digestivo, ya que está rodeado de epitelio, lo que genera que sea muy doloroso tragar, … Esta enfermedad es de origen genético. Todo el núcleo tiene la lámina nuclear menos el complejo del poro (zonas menos ordenadas). Esto permite que por el poro se transporten diversas sustancias. Esta lámina nuclear proporciona resistencia mecánica a la envuelta nuclear. Ya que los movimientos de la célula también los sufre el núcleo. Si quito la lámina nuclear, la envuelta nuclear se desintegra, porque no es capaz de asumir el estrés mecánico a la que la envuelta se enfrenta solo por existir (por eso regula el desensamblaje y el ensamblaje de la envuelta en cada ciclo celular). Cuando empieza el proceso de división celular, un factor de crecimiento activa a las kinasas, que actúan fosforilando a los filamentos intermedios para generar la desaparición de la envuelta. Esto supone que en las uniones entre los tetrámeros se va a ver “interrumpida” por los grupos PO-4. Ya que son cargas opuestas enfrentadas. Cuando llego a la telofase, las quinasas se inhiben y se activan unas fosfatasas, que hacen desaparecer todos esos grupos fosfato. Así, la lámina nuclear se vuelve a formar y la envuelta nuclear se ensambla de nuevo. MOVIMIENTOS CELULARES Pueden ser: movimientos internos, desplazamientos o creación de corrientes. En estos movimientos van a estar implicadas, entre otras, las proteínas motoras. Estas proteínas ayudan al citoesqueleto a realizar parte de los movimientos anteriormente mencionados, junto con los microtúbulos y los microfilamentos. Estas proteínas pertenecen a la familia de las miosinas (movimiento relacionado con los microfilamentos→ su papel es producir los movimientos en sustrato sólido (ameboides) y formar cualquier tipo de estructura contráctil); dineínas y quinesinas (colaboran con los microtúbulos y van a estar implicadas en desplazamientos gracias a flagelos, la formación de corrientes (depende de los cilios) y con los movimientos intracelulares). En el caso de las dineínas hay dos tipos diferentes que dependiendo de la función van a estar solo en el citosol (dineínas citoplasmáticas) o las presentes en cilios y flagelos (dineínas ciliares). PROTEÍNAS MOTORAS: GENERALIDADES Todas las proteínas motoras se tienen que unir mediante enlaces a los filamentos del citoesqueleto (por ello existe la especificidad). Todas van a hidrolizar ATP, para tener la energía necesaria para los movimientos. Todas van a tener sitios de unión para el ATP y van a tener actividad enzimática que les permite romper el ATP. Van a poder transportar: macromoléculas, orgánulos, vesículas (exocitosis y secreción), … Lo que supone que van a tener dos sitios de unión. El funcionamiento es el desplazamiento de la carga sobre el citoesqueleto (dirección= extremo más y menos→ las proteínas solo se pueden mover en una dirección concreta: dineínas siempre van hacia menos, las quinesinas van hacia más). Cuando necesito producir tensiones, lo que se necesita es que una proteína ande sobre un filamento del citoesqueleto y su carga sobre otro. De esa manera, se producen deslizamientos de unos filamentos sobre otros (si contráctil= actina; si cilio o flagelo= microtúbulo). Se dice que todas tienen una cabeza (proteínas globulares→ capacidad motora, ya que genera el movimiento uniéndose al ATP e hidrolizándolo, lo que genera un cambio de conformación, también define si una carga se mueve mediante un microtúbulo o un microfilamento y en qué dirección se mueve) y una cola (proteína con forma de hélice alfa: la cola determina la función biológica, ya que indica a qué carga te puedes unir). La forma de la proteína suele asemejarse a la de miosina II. Miosina II: dos cadenas pesadas y una cola. La zona donde se juntan las cabeza, se denomina bisagra (la longitud de la bisagra varía de unas a otras. Cuanto más larga sea= más velocidad (la miosina es de las más rápidas) y es la encargada de cambiar de forma. A su vez, está unida a cadenas ligeras. Estas cadenas están donde se producen los cambios conformacionales porque tienen actividad reguladora (casi siempre se unen a Ca 2+). Pies= cabezas y bisagras= piernas. Se gasta un ATP por cada paso. FUNCIONES Dependen de tu carga, pero lo más habitual es mover las cargas de un lado a otro del citoplasma. - Ayudan a los filamentos del citoesqueleto a reorganizarse de distintas formas para formar un lamelipodio, un filopodio, cilios, …. Encargados de comportamientos celulares complejos (división celular→ supone muchas variables coordinadas. En estos cambios, las MAP, van a estar implicadas). SUPERFAMILIA DE MIOSINA Se desplazan sobre microfilamentos (actina), aunque no todas hacen exactamente lo mismo, y por ello van a actuar en procesos diferentes. Esta familia está directamente implicada en la contracción muscular. Su zona de unión a los filamentos de actina está hecha para encajar, se une con los microfilamentos mediante enlaces débiles. La dirección de la miosina siempre es el extremo más (no hace falta recorrer el microtúbulo entero). Al ser una familia grande, se distinguen poniendo un número romano después de su nombre. Las más importantes son la miosina II, I y V. La miosina II está formada por cadenas polipeptídicas con un extremo globular (se une al ATP, lo hidroliza y produce el movimiento): La miosina I solo tiene una cabeza pesada. MIOSINA V Es especialista en unirse a cosas grandes que hay que mover dentro de la célula de un sitio a otro (Ej.: mover mitocondria allá donde sea necesario ATP, un ejemplo podría ser un flagelo). Puede unirse a cargas con membrana (vesículas) o a cargas que no la tengan (ribosomas, …). MIOSINA I Unión mediante uno de sus extremos a las membranas de la célula (citoplasmática o de orgánulos) y con el otro a un microfilamento. Puede unirse a la membrana citoplasmática por un lado y al microfilamento por otro. De tal manera que como la unión de la membrana y la miosina I es fija, el microfilamento se tiene que desplazar. Dando lugar a protusiones. Por ende, esta MAP nos sirve para formar protusiones y llevar a cabo desplazamientos. Para llevar a cabo estos movimientos es importante que TODO el citoesqueleto se coordine. Quienes dirigen el movimiento son los filamentos de actina del córtex, el resto solo sigue sus órdenes. Cuando una célula se está moviendo, va a haber una parte de la célula de mayor tamaño ( en la dirección que se desplaza→ donde miosina I+ filamentos de actina) y una zona más pequeña donde se generan movimientos contráctiles para empujar a todo el citoplasma (miosina II+actina). Por eso, a la zona que avanza de forma más lenta, se la denomina zona retrasada. MIOSINA II Cuando forma estructuras contráctiles (células musculares y no musculares→ contactos focales con fibras de estrrés para movimientos ameboides, anillo contráctil, …). Las cabezas de la miosina II se pueden unir a dos filamentos a la vez, de tal manera que se desplazan el uno sobre el otro, ya que las miosinas no se pueden separar. Esto produce la contracción de la célula. Esto también se produce debido a que los dos filamentos se encuentran en una posición antiparalela. Todo lo que “brilla” son filamentos de actina, pero los verdes son los filamentos que están creciendo. MOVIMIENTOS EN SUSTRATO SÓLIDO (MOV. AMEBOIDES) En los movimientos ameboides va a haber tres fases: Estos movimientos los llevan a cabo todas nuestras células menos los espermatozoides. Para que estos movimientos ameboides se produzcan necesitamos la colaboración del citoesqueleto (filamentos de actina y las MAP son especialmente relevantes), que permite que se lleven a cabo las protusiones. Durante el desarrollo embrionario es fundamental que hay células que se muevan de donde están para ir a otro sitio y generar una nueva estructura. Ej..: gastrulación. Los macrófagos y los neutrófilos circulan por la sangre hasta que reciben una orden para salir de allí y atacar a la célula. Esto lo hace mediante mov. ameboides. Los fibroblastos suelen permanecer en un sitio fijo, pero, cuando se produce una lesión, van a esa zona para reparar los tejidos dañados, también mediante mov. ameboides. Cuando una neurona se está formando y ha des establecer sinapsis con una célula más o menos cercana. La célula genera señales como las citoquinas y la neurona, genera filopodios para detectar de dónde proviene la señal. Estos filopodios crecen en la dirección de la señal, generando el crecimiento del axón. El desarrollo de los filopodios se lleva a cabo mediante movimientos ameboides. También se llevan a cabo en las metástasis→ varias células se separan del tumor primario hasta llegar a un vaso sanguíneo y expandirse en otros lugares. Esto también es debido a los mov. ameboides. También son relevantes los contactos focales. 1) Protusión: es necesario generar evaginaciones de distinto tipo según las necesidades celulares. Para crear esas evaginaciones la actina se polimeriza y se produje un empuje hacia delante. 2) Adhesión: es importante que la membrana se una al sustrato sobre el que se moviliza. Esto se hace mediante contactos focales, ya que son pequeños, débiles y se pueden formar sin gasto de ATP. La célula no levita. 3) Tracción: La parte de atrás avanza gracias a las uniones entre miosina II y los microfilamentos (=filamentos de actina). Para ello, he de romper los contactos focales que se habían establecido en la parte posterior. La velocidad a la que formas las uniones adelante, tiene que ser la misma a la que rompes las uniones por detrás, porque sino, se podría producir la ruptura de la célula. Normalmente las células se mueven en una dirección específica que responda a las señales que le llegan. Ej.: neutrófilos→ llega una señal emitida por un tripéptido (formil-Met-A-B). De tal manera que le está llegando la información de que hay una bacteria. Por ello, va a formar lamelipodios hacia ese tripéptido para poder fagocitar a la bacteria. Cuando la señal cambia de dirección, los lamelipodios hacen lo propio. DESPLAZAMIENTO SOBRE MICROTÚBULOS Se emplean otro tipo de proteínas motoras, ya que tiene que existir la compatibilidad entre las proteínas y el citoesqueleto. Por ello, en los microtúbulos se emplean las dineínas (se mueven hacia menos) y las quinesinas (van hacia más). Tienen una estructura muy similar a la miosina II. Tienen dos cadenas pesadas y un tallo. En el caso de la dineína es un poco menos similar porque es más pequeña. El movimiento se produce en todas de una forma similar→ hay una proteína motora que se une al microtúbulo mediante las cabezas y mediante el tallo se unen a la carga. Las cabezas forman uniones débiles con el filamento del citoesqueleto. Como funcionan como si fueran pies, una cabeza va a estar más adelantada que la otra. Lo que significa que la de adelante va a estar unida a ADP y la de atrás a ATP. Cuando la de atrás hidrolice el ATP, va a romper los enlaces que la mantenían unida al microtúbulo se van a romper y, va a avanzar (cambio conformacional), además de formar nuevos enlaces. Para volver a conseguir el ATP, una proteína intercambiadora de nucleótidos va a transformar el ADP en ATP. Así, el movimiento puede continuar. En las células no polarizadas: Unirte a una quinesina significa que vas a moverte desde una zona más cercana al núcleo a una más cercana a la membrana citoplasmática. Esos movimientos se van a denominar movimientos centrífugos. En la dineína va a ocurrir lo contrario, en este caso los movimientos se denominan centrípetos. En las células polarizadas: El extremo menos está lo más cercano posible al soma y el extremo más, lo más lejos posible. Cuando se unen a una quinesina= movimiento anterógrado. Cuando se unen a una dineína= movimientos retrógrados Ej.: neurotransmisores se secretan y luego regresan para ser reciclados. Empleo de quinesinas y dineínas, respectivamente. QUINESINAS Se unen a los microtúbulos y avanzan en dirección más. Por lo que van a participar en movimientos centrífugos o anterógrafos. La quinesina 1 tiene un extremo amino-terminal y los extremos carboxilo-terminales en el final de su cola. La quinesina 5, va a ser similar a la miosina II ya que también tienen una disposición antiparalela. Esta proteína es fundamental para la formación del huso mitótico. Ya que es fundamental que los microtúbulos se solapen para formar ese huso mitótico. La quinesina 14 es la única que va hacia menos, ya que tiene las cabezas en el sentido contrario. Esto es debido a que su estructura está invertida. En el caso de la quinesina 13, las cabezas polares están en el centro. A esta quinesina se le llama factor catástrofe ya que consigue la despolimerización de los microtúbulos. Las quinesinas se denominan una superfamilia con dominio motor común, porque tienen una cabeza con una composición química muy similar. Ya que la composición variable es la de la cola. FUNCIONES QUINOSINAS -Transporte de vesículas membranosas, orgánulos, macromoléculas, enzimas (esto lo haría principalmente la quinesina I). -Uniones a otros microtúbulos (esto produce deslizamientos, fundamental para: la formación del huso mitótico y la segregación de las cromátidas). En la segregación de las cromátidas, los microtúbulos se despolimerizan y se acortan, para favorecer esta separación y el huso mitótico se alarga. DINEÍNAS Todas ellas se desplazan hacia el extremo menos, pero tienen una estructura diferente. Poseen dos o tres cadenas pesadas (cabezas). En el caso de las citoplasmáticas son 2 cadenas pesadas y son homodímeros. Sin embargo, en las dineínas ciliares van a estar presentes tres cabezas y las cadenas polipeptídicas van a estar formadas por heterotrímeros y heterodímeros. FUNCIONES -Dineínas citoplasmáticas: su función es el tráfico de sustancias de un sitio a otro del citosol. Esas sustancias pueden ser orgánulos con o sin membranas, macromoléculas, …. También se van a encargar de determinar la localización del Aparato de Golgi. -Dineínas ciliares: andan sobre un microtúbulo y su carga es otro microtúbulo, eso es lo que va a permitir el movimiento de los cilios y los flagelos de las células eucariotas. TRANSPORTE INTRACELULAR DE VESÍCULAS MEMBRANOSAS En un corte transversal observaríamos una estructura diferente: Hay orgánulos como por ejemplo el Aparato de Golgi, que se heredan. En la diapositiva A. Se pueden ver dos células en las que el Aparato de Golgi está teñido de verde. Como podemos ver, en una de ellas el Golgi se encuentra a ambos lados del núcleo. Es decir, las dos células hijas reciben algo de Golgi, ya que cada una se lleva un parte. Esto es posible gracias a que, al replicarse los centriolos, va a haber dos extremos menos y esto lo toma de referencia el aparato de Golgi para desplazarse. Esto va a ser gracias a las dineínas citoplasmáticas. -En la zona central habría dos microtúbulos (dos cilindros huecos) que no están unidos entre sí. Sin embargo, en la imagen B, el Aparato de Golgi no sabe a dónde ir. -Alrededor, me encuentro con 9 parejas de microtúbulos, (A= 13 protofilamentos, B=10/11 protofilamentos). Están adosados para compensar la falta de esos protofilamentos por parte de B. Las uniones entre las distintas parejas siempre van a ser A-B. De cada par periférico salen estructuras que conectan a A con la vaina. A estas estructuras se las denomina fibras radiales (para que los cambios conformacionales sean informados a toda la estructura). También van a existir puentes de nexina, que comunican a un par periférico con otro. Desde A salen los brazos de dineína (interno y externo), que se acercan a B, pero solo se unen al par vecino si hay que moverse. En el caso del retículo endoplasmático, se van a encargar de su desplazamiento, las quinesinas. Ya que estas se encargan de disponer las nuevas cisternas más cerca de la membrana. Si quitásemos las quinesinas pasaría lo mismo que en la imagen B. En el caso de las dineínas ciliares, se encuentran exclusivamente en los cilios y los flagelos de las células eucariotas. El interior de los cilios y los flagelos va a estar formados por proteínas accesorias que les ayudan a ser más estables (formación de CAPs) y otras que permiten su movimiento. Los cilios y los flagelos tienen la misma estructura interna pero los cilios son más finos y cortos que los flagelos. Asimismo, la proporción en la que se encuentran en la célula es distinta, ya que suele haber menos flagelos en las células flageladas que cilios en las células ciliadas. -A su alrededor, se dispone la vaina central→ envuelve a ambos y los conecta (aunque no están unidos físicamente, hay proteínas que van de uno a otro y que les informan de los cambios del microtúbulo adyacente). Por eso se dice que el axonema tiene una estructura 9+2. Se habla de proteínas accesorias a intervalos regulares porque están establecidas a lo largo de toda la estructura de forma regular. SI hiciera un corte longitudinal del corte transversal vería la siguiente imagen: Donde se observan los microtúbulos periféricos y los centrales. La función es permitir el movimiento en un medio acuoso, pero en nuestras células los cilios no son un medio de locomoción. En nuestro caso, nuestras células emplean los cilios para generar corrientes o mover sustancias externas. En nuestro organismo, las células ciliadas son las que están polarizadas (en la zona apical van a estar presentes los cilios). Los cilios han de moverse de forma coordinada para lograr su objetivo. Cuando se observa a cilios y flagelos mediante un microscopio electrónico se ven igual y poseen tres zonas (corte longitudinal): -La que sobresale: tallo o axonema. -La que está a la altura de la superficie celular: zona de transición. -La que está dentro del citosol: corpúsculo basal. Se distinguen de las microvellosidades porque dentro de las microvellosidades me voy a encontrar con una gran cantidad de filamentos de actina que llenan todo el interior de la microvellosidad. En la zona de la membrana, en la base del flagelo, se encuentra la zona de transición. Es el paso del tallo o axonema a la placa basal. Se llama así porque está en la base del tallo y porque es plana (es densa a los electrones→ Está compuesta por proteínas acumuladas en esa zona (anillos de gamma-tubulina, tubulina, GTP y GDP, …). Con el corte longitudinal, se observa la interrupción del par central. Si lo observo desde un corte transversal en vez de una estructura 9+2, vería una estructura 9+0. Por eso puedo distinguir la zona en la que estoy del cilio o del flagelo independientemente del tipo de corte. Si sigo profundizando, me encontraré debajo de la superficie de la membrana y observaría el corpúsculo basal. Con el corte longitudinal observaré que se va a formar 9 tripletes periféricos. En un corte transversal podré ver la estructura de los tripletes en los que ya no hay brazos de dineína porque NO se mueve. Sí que contiene fibras de nexina. Tanto B como C van a ser incompletos, solo van a tener 10 o 13 protofilamentos. La estructura en esta zona también es 9+0. Dentro del corpúsculo basal voy a tener dos zonas diferenciadas: 1) Región distal: el centro es el citosol (distal del núcleo de la célula). 2) Región proximal: proteínas que van al centro de la estructura y que conforman las láminas radiales, porque tienen forma de cinta. Lo que hay en su interior es un “agujero” causado por la imposibilidad de las láminas radiales para llegar al interior. Se dice que tiene estructura de rueda de carro. De los cilios y flagelos parten unas proteínas hacia el interior de la célula que se denominan raíces ciliares, que se dirigen a una zona próxima al centro para coordinar los movimientos. De esta forma, todos los cilios y flagelos se moverían al mismo tiempo. CORPÚSCULO BASAL -9 tripletes periféricos unidos mediante puentes de nexina entre A y C. Dependiendo del corte podré observar mejor o peor los dos microtúbulos internos. Las dineínas de los cilios se van a unir a un microtúbulo del par vecino, y van a empezar a andar sobre B. Cuando no hay movimiento esos brazos de dineína salen de A, pero no llegan a tocar a B. La dineína no es la única que une los dos pares, también está la nexina, que es fija. Lo que ocurre es que no hay un deslizamiento en línea recta, sino que, debido a la nexina, el movimiento se curva. La dirección en la que se curvan decide la dirección del movimiento. Este movimiento supone un gran gasto de energía. No solo por la cantidad de ATP que gasta la energía, sino porque cuando uno se mueve, lo hacen todos a la vez. En los cilios, el movimiento se produce gracias a las dobleces de los cilios, pero para poder avanzar, estos han de volver a su forma original. Cuando los tallos se doblan en la dirección del movimiento→ bateo eficaz. Cunado los tallos vuelven a su forma→ bateo de recuperación. Como los cilios son cortos, el movimiento es ondulante. Pero el flagelo al ser más largo emite ondas para la propulsión de la célula en una determinada dirección. SÍNDROME DE KARTAGENER Sin la dineína, este movimiento no se podría llevar a cabo. En este síndrome, la dineína se puede producir y no funcionar o no producirse. En los varones esto puede causar esterilidad. Estas personas suelen tener infecciones respiratorias, ya que los cilios se encargan de desplazar el moco y que no se acumulen bacterias. Cuando la dineína no funciona, las bacterias no se eliminan y se generan infecciones. También se puede originar el situs inversos. Esto se origina debido a que se producen alteraciones en el movimiento en la gastrulación, suponiendo que las estructuras que tengan que estén a la derecha estén a la izquierda y viceversa. LOS RIBOSOMAS Sintetizan proteínas tanto en las células procariotas y eucariotas debido a que su forma es muy similar. Aunque las proteínas que sintetizan son ligeramente distintas en eucariotas y procariotas (forma, composición, …). -Forma y función similares: subunidad grande y subunidad pequeña. Cada subunidad tiene una velocidad de sedimentación. Cuando bajan separadas no suman la misma velocidad que cuando bajan juntas. Por eso, las dos piezas por separado no tienen la misma forma que cuando están juntas. En cuanto a su forma, son muy parecidas, pero no idénticas ya que al tener una composición química diferente, va a propiciar ligeros cambios conformacionales. ARN En las células hay muchos tipos de ADN. Algunos están libres en el citosol, otros están en el núcleo, …. En un principio hay dos formas de clasificación: 1) ARN codificantes: ARNmensajero, se encuentran en el núcleo y salen al citosol para que los lean los ribosomas. 2) ARN no codificantes: ARNribosomal (forman parte de los ribosomas, uno de ellos cataliza la síntesis de proteínas que sintetiza los enlaces peptídicos); ARNtransferencia (coge aas y los lleva al ribosoma); ARNsn (hebras de pequeño tamaño que se encargan del splicing→ indican a la enzima dónde empieza el intrón y dónde acaba y que se encuentra en el núcleo); snoRNA (pequeños y nucleolares, están en el nucleolo. Es la maduración postranscripcional de los ARN ribosomales); los scaRNA, miRNA y siRNA regulan la expresión génica. Otros se encargan de la inactivación del cromosoma X (en el caso de las mujeres), por lo que solo usamos uno de los dos cromosomas X y la síntesis de la telomerasa. PROCARIOTAS Y EUCARIOTAS -Distinta velocidad de sedimentación (70s y 80s). La s viene de Svedberg que es una unidad que mide la velocidad a la que caen las cosas cuando las sometes a un proceso centrífugo). Para ello introduzco en un tubo de ensayo una gelatina, en la que introduzco ribosomas de células procariotas y células eucariotas. Después de la centrifugación, los ribosomas de las eucariotas bajan más rápido que los eucariotas. Esta rapidez viene de que son más grandes, pesan más y son más densos. -Distinto tamaño. -Distinto peso. -Distinta densidad. -Distinta composición química: están formados por ARNr ambos, pero no son del mismo tipo. Así como sus proteínas (también son distintas). ARNr Dentro de los ribosomas de las procariotas hay dos tipos de ARNr, dependiendo de la unidad a la que corresponda. Dentro de las eucariotas hay tres tipos de ARNr. Dos de ellos se encuentran en la subunidad mayor. Esta comparación también se puede hacer con el ARN, porque se puede medir su velocidad de sedimentación. Un ribosoma está formado solo por ARNr (2/3) y proteínas (1/3). Ese ARNr, es el más abundante de todos los tipos de ARN que tenemos en las células. Estos ARNr son un tanto peculiares en comparación a los ARNm: -El 70% de la molécula está dispuesta en doble hélice. Ya que antes de llegar al final se producen gran cantidad de giros, llevados a cabo por bases nitrogenadas metiladas (poseen un grupo químico que no sea capaz de formar puentes de hidrógeno. La secuencia de nucleótidos de un lado del giro, son complementarios con respecto al que está enfrente, de tal manera que se generan uniones entre pares de bases, produciendo la doble hélice. A su vez, está estructura se pliega sobre sí misma. Estas estructuras se van a disponer en las zonas más internas de las subunidades de los ribosomas. Esto es debido a los anillos de las bases nitrogenadas, lo que les confiere gran apolaridad. La estructura del ribosoma es muy compacta, la parte más interna (core) es donde se encuentran los ARN y en la superficie estarían las proteínas ribosomales (hidrófobas→ lo que permite a los ribosomas estar en el citosol). Para formar un ribosoma, se lleva a cabo un autoensamblaje ribosomal (muchas copias en un lugar pequeño de la célula en el que tiene lugar la unión entre las subunidades. También es cooperativo, cuantas más se unan, más fáciles son las uniones). Es importante que la célula pueda diferenciar los distintos tipos de ARN. Lo que supone que por ejemplo el ARNm es lineal, esto es peligroso porque hay exonucleasas que se encargan de romper los enlaces de los extremos del ARN y las endonucleasas que se encargan de partir trozos de ARN. Ambas son ribonucleasas. En los ribosomas y en sus subunidades, para que se lleve a cabo la síntesis de proteínas son fundamentales los sitios de unión al ARNm y al ARNt. Esas uniones entre los ribosomas y el mensajero y transferentes se llevan a cabo mediante puentes de hidrógeno entre bases complementarias. Van a ser muy eficaces frente a las exonucleasas gracias a la formación de CAPs en los extremos, en los que forma enlaces 5´-5´. Porque la enzima solo sabe romper enlaces 3´-5´. En la subunidad grande hay tres sitios de unión al ARNt: En el extremo 3´se sitúa una cola de poliA, lo que supone la unión de 300-500 adeninas. -Sitio A o aminoacilo: al que se une un transferente que trae un aa del citosol para añadirlo a una cadena polipeptídica que está sintetizando. En el caso del resto de ARN la forma va a ser distinta y no van a tener esas estructuras protectoras. -Sitio P o peptidilo: se une a ARNt para que se forme una cadena larga mediante enlaces peptídicos. Los ARNr van a tener funciones estructurales y catalíticas (hay un ARNr que es una ribozima→ cataliza la creación del enlace peptídico). El ARN que lleva a cabo está función es el de la subunidad grande (28S en eucariotas y 23S en procariotas). -Sitio E o exit: sitio por donde sale el ARNt porque ya ha depositado su carga. Es decir, si asignáramos el trabajo según la subunidad, podríamos decir que la subunidad pequeña se encarga de detectar al ARNm y unirse a él, saber dónde está el codón de iniciación y poder traducir codón a codón (sin saltarse espacios, …) y la subunidad grande sería la encargada de la generación del enlace peptídico. En la subunidad pequeña: Esos tres lugares no pueden estar ocupados a la vez (no hay espacio), o están A y P o P y E. -Un sitio de unión para el mensajero: hendidura. MORFOLOGÍA -La subunidad grande posee una cresta, una protuberancia central y el tallo. Finalmente, entre las tres protuberancias hay un hueco al que denomino valle. -La subunidad pequeña se divide en cabeza, hendidura y plataforma. En la hendidura se enhebra el mensajero. Estas subunidades son bastante grandes por lo que pueden ser observadas por un microscopio electrónico. La parte gris es ARNr y la parte naranja son las proteínas ribosomales. En la imagen de la subunidad grande C, se puede ver un agujero que es por el que sales los ARNt, es decir, el sitio E. UNIÓN SUBUNIDADES REGIONES FUNCIONALES La unión de las subunidades se realiza mediante enlaces débiles (se unen de esta forma porque las subunidades solo se unen cuando trabajan en la síntesis de proteínas). En el interior de la estructura todo se mantiene gracias a puentes de hidrógeno, aunque también participan las uniones hidrófobas entre bases modificadas. Los sitios de unión de cada subunidad definen la función de cada sitio. Las responsabilidades son distintas en cada subunidad: -Subunidad pequeña: detecta dónde está el CAP 5´, recorre el mensajero y leer la secuencia hasta el codón de iniciación (no está al principio→ es AUG pero antes de ese AUG hay una secuencia de nucleótido que le indica que se está acercando) y pararse en ese momento para realizar el ensamblaje con la subunidad grande. -Subunidad grande: se une a los ARNt y los coloca en el sitio adecuado para que se produzca la síntesis. También contiene una ribozima encargada de sintetizar el enlace peptídico→ peptidil-transferasa (transfiere un péptido de un sitio a otro). Para que este proceso se realice necesito energía que proviene de: -Gasto de 1 molécula de ATP por cada aa que uno al ARNt. -Gasto de GTP en el ribosoma para formar el enlace peptídico. Todas las células tienen ribosomas. Dentro de nuestras células pierden los ribosomas a medida que se diferencian. Un ejemplo son los espermatozoides y los eritrocitos. Aunque en las fases iniciales del desarrollo todas van a tener ribosomas. Según el destino de las proteínas -Síntesis en el citosol: proteínas que van a vivir en el citosol. -Síntesis en el RER: proteínas de orgánulos y estructuras extracelulares. Si un ribosoma está unido a la membrana del retículo es porque está sintetizando una proteína. -Síntesis mitocondrias: (protorribosomas, más parecidos a procariotas). -Síntesis en cloroplastos. Estos ribosomas pueden estar aislados o pueden formar complejos denominados polisomas (todos leen y sintetizan a la vez la proteína). Los ribosomas cumplen mejor su función cuando forman polisomas que cuando están aislados.