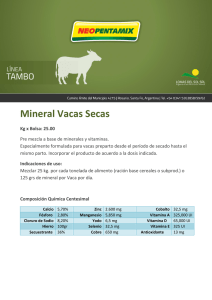
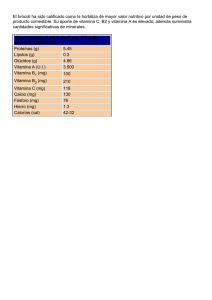
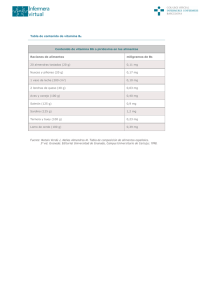
1.24.Vitamina D Olga Martínez Augustin Víctor Puerta Fernández María Dolores Suárez Ortega Capítulo 1.24. Vitamina D 1. Introducción 2. Absorción de la vitamina D 3. Fotobiogénesis 4. Metabolismo hepático y renal de la vitamina D 5. Inactivación y excreción de la vitamina D 6. Control de la síntesis y degradación de vitamina D 7. Fuentes de vitamina D 8. Requerimientos nutricionales de vitamina D 8.1. Ingesta máxima tolerada de vitamina D 9. Mecanismo de acción de la vitamina D 9.1. Receptores de vitamina D 9.1.1. Receptor de membrana del 24-R-calcitriol (VDRmem24,25) 9.1.2. Receptor de membrana del calcitriol (VDRmem1,25) 9.1.3. Receptor nuclear del calcitriol (VDRnuc1,25) 9.1.3.1. El gen del VDRnuc1,25 humano (hVDRnuc1,25) 9.1.3.2. Estructura proteica del VDR humano y su regulación mediante fosforilación 9.1.3.3. Factores implicados en la regulación transcripcional por calcitriol 10. Acciones de la vitamina D 10.1. Acciones clásicas de la vitamina D. Homeostasis mineral 10.1.1. Intestino 10.1.2. Hueso 10.1.3. Riñón 10.2. Acciones no clásicas de la vitamina D 10.2.1. Efectos de la vitamina D sobre la proliferación celular, la diferenciación celular y la apoptosis 10.2.1.1. Cáncer 10.2.2. Efectos de la vitamina D sobre el sistema inmune 10.2.2.1. Infecciones 10.2.2.2. Inflamación y enfermedades autoinmunes 10.2.2.3. Artritis reumatoide 10.2.2.4. Enfermedad inflamatoria intestinal 10.2.2.5. Esclerosis múltiple 10.2.3. Efectos de la vitamina D sobre el sistema renina-angiotensina 10.2.4. Efectos de la vitamina D sobre el sistema nervioso 11. Niveles normales y deficiencia de vitamina D 12. Enfermedades relacionadas con alteraciones en el metabolismo de la vitamina D o en la respuesta a vitamina D 12.1. Raquitismo resistente a vitamina D o hipofosfatemia familiar 12.2. Hipoparatiroidismo 12.3. Raquitismo tipo I dependiente de vitamina D 12.4. Raquitismo tipo II dependiente de vitamina D 13. Resumen 14. Bibliografía 15. Enlaces web Objetivos n Conocer y comprender el metabolismo de la vitamina D. n Saber las necesidades nutricionales de la vitamina D y los alimentos ricos en esta vitamina. n Conocer las formas moleculares activas de la vitamina D y relacionarlas con su mecanismo de acción. n Describir el efecto de la vitamina D sobre la homeostasis mineral, identificar los órganos diana y conocer los mecanismos de regulación. n Conocer las acciones de la vitamina D sobre la diferenciación y la proliferación celular. Relacionar estas acciones con la posible utilización de análogos de la vitamina D en el cáncer. n Estudiar las acciones de la vitamina D sobre el sistema inmune, el sistema nervioso y el sistema reninaangiotensina. Relacionar estas acciones con el efecto beneficioso de esta vitamina y de sus análogos en el tratamiento de distintas enfermedades. n Observar los efectos derivados de la deficiencia y del consumo excesivo de vitamina D. n Conocer las enfermedades relacionadas con alteraciones en el metabolismo de la vitamina D o en la respuesta a la vitamina D. Relacionar la utilización de distintas formas y análogos de esta vitamina en el tratamiento de estas enfermedades. 1. Introducción E xisten algunas referencias a enfermedades óseas parecidas al raquitismo atribuidas a Soranus de Éfeso, un médico que practicó la medicina en Roma durante los reinados de Adriano y Trajano. No obstante, hasta el siglo XVII, cuando el raquitismo era endémico en Europa, no aparecen más descripciones de esta enfermedad. Aunque la asociación entre el raquitismo y la deficiencia de vitamina D no se estableció hasta principios del siglo XX, ya en 1807 Bardsley escribió sobre el uso del aceite de hígado de bacalao en la prevención de la osteomalacia, y Palm en 1890 sugirió que la luz del sol poseía acción antirraquítica. El concepto de vitamina fue introducido por primera vez en 1911 por Funk; en 1913 McCollum y Davis describieron la existencia de un factor en el aceite de hígado de bacalao que era esencial para el crecimiento y que denominaron vitamina A. Unos años después, en Inglaterra (1919), Edward Mellanby et al. indujeron raquitismo a perros mediante manipulación dietética y observaron que la administración de aceite de hígado de bacalao era capaz de producir la curación, asumiendo que la vitamina A era capaz de prevenir y curar el raquitismo. Posteriormente, McCollum et al. (1922) demostraron que la destrucción de la vitamina A del aceite de hígado de bacalao mediante oxidación no eliminaba su actividad de prevención del raquitismo, por lo que dedujeron que este efecto debía deberse a un factor resistente al calor y la aireación al que denominaron vitamina D. Simultáneamente, Huldschinsky (1919) demostró mediante estudios clínicos que la exposición de niños a la luz solar o a luz ultravioleta era también capaz de prevenir o curar esta enfermedad. Por tanto, la cura del raquitismo parecía estar relacionada tanto con la exposición a la luz solar como con sustancias presentes en el aceite de hígado de bacalao. Steenbock et al. describieron, en los años 20 del siglo XX, que la irradiación de ciertos alimentos y productos biológicos podía inducir actividad antirraquítica en estos productos. Este descubrimiento proporcionó información crucial para el aislamiento y la identificación de la vitamina D2 y para la curación y eliminación del raquitismo como un problema médico importante, ya que llevó a la idea de que los alimentos podían ser fácilmente suplementados con esta vitamina y que la irradiación de ciertos alimentos como la leche o el pan podía hacer que éstos fueran útiles en el tratamiento del raquitismo. Actualmente se considera que la vitamina D es una vitamina y una hormona. Así, es un compuesto orgánico que actúa como micronutriente, y su ingestión es necesaria para la mayoría de las poblaciones urbanas; de aquí que se considere una vitamina. De hecho, como la vitamina D no es muy abundante en la dieta, es adicionada a distintos alimentos en varios países, y su deficiencia es frecuente en invierno en países en los que los periodos de tiempo sin sol son largos. A pesar de lo anterior, la suplementación con vitamina D es innecesaria en individuos que son capaces de completar sus requerimientos mediante la activación por irradiación (luz solar) del 7-deshidrocolesterol, un metabolito del colesterol que se 793 Capítulo 1.24. Vitamina D produce en el hígado y es exportado a la piel. La vitamina D producida en la piel por irradiación es a continuación metabolizada sucesivamente en el hígado y el riñón, produciéndose las formas activas que actúan sobre distintas dianas. Por tanto, los metabolitos 1α,25 dihidroxivitamina D3 (calcitriol), y 24R,25 dihidroxivitamina D3 (24-R-calcitriol) que se producen en condiciones normales en el riñón, independientemente de la dieta, y que actúan sobre los distintos órganos diana, pueden ser considerados hormonas, y la vitamina D una prohormona. El término genérico “vitamina D” agrupa a dos moléculas distintas: el ergocalciferol o vitamina D2 y el colecalciferol o vitamina D3. Según la nomenclatura moderna, las vitaminas D2 y D3 se denominan ercalciol y calciol, respectivamente. La vitamina D2 posee un doble enlace adicional en la cadena lateral y se produce mediante irradiación del ergosterol procedente de plantas, por lo que proviene necesariamente de la alimentación. Por su parte, la vitamina D3 es la principal fuente de vitamina D en la naturaleza, que puede ser producida de manera endógena mediante la irradiación del 7-deshidrocolesterol (un derivado del colesterol) o bien proceder de la alimentación. La vitamina D2 se utiliza en la suplementación de alimentos y se empleó en el pasado más ampliamente que la vitamina D3 en el tratamiento de enfermedades relacionadas con deficiencias de vitamina D, porque era más barata y porque se creía que ambas eran igual de potentes. Un gran número de estudios posteriores demostraron que la vitamina D2 es menos tóxica que la D3. No obstante, en este Capítulo se va a estudiar, preferentemente, la vitamina D3, ya que los mecanismos de acción, el metabolismo y, en líneas generales, las características de ambas vitaminas son muy parecidas. La Tabla 1 recoge la nomenclatura antigua y moderna de los compuestos relacionados con la vitamina D. 2. Absorción de la vitamina D La vitamina D ingerida en la dieta es generalmente absorbida con las grasas en el duodeno y el íleon, siendo necesaria la presencia de ácidos biliares para que se produzcan las correspondientes micelas. Como consecuencia, la inhibición de la absorción de grasas da lugar a una disminución en la absorción de vitamina D. Además, en pacientes con pancreatitis crónica, enfermedad celiaca u obstrucción biliar también se produce malabsorción de la vitamina D. Una vez absorbida por la mucosa intestinal, la vitamina D se incorpora a los quilomicrones y es exportada por vía linfática al hígado, donde se libera de éstos. La captación de la vitamina D en el hígado se realiza con el concurso de una proteína específica denominada DBP (D-Binding Protein: proteína de unión o fijadora de vitamina D), que es sintetizada en el propio hígado. Esta proteína es una α-globulina que actúa también como transportadora en la sangre de la vitamina D y de todos sus metabolitos; se han descrito al menos 37 metabolitos de la vitamina D3 -aunque sólo la 25-hidroxivitamina D3 [25(OH) vitamina D3], el calcitriol y el 24-R-calcitriol son activos- a los que la BDP se une con distinta afinidad. La DBP es también conocida como Tabla 1. NOMENCLATURA DE LOS COMPUESTOS RELACIONADOS CON LA VITAMINA D 794 Vitamina D3 Colecalciferol Calciol 25-hidroxivitamina D3 25-hidroxicolecalciferol Calcidiol 1α,25-dihidroxivitamina D3 1α,25-dihidroxicolecalciferol Calcitriol 24R,25-dihidroxivitamina D3 24R,25-dihidroxicolecalciferol 24-hidroxicalcidiol Vitamina D2 Ergocalciferol Ercalciol 25-hidroxivitamina D2 25-hidroxiergocalciferol Ercalcidiol 1α,25-dihidroxivitamina D2 1α,25-dihidroxiergocalciferol Ercalcitriol O. Martínez Augustin | V. Puerta Fernández | M.ª D. Suárez Ortega 1α-globulina o componente específico del grupo (Gc). Se puede considerar que, además de proporcionar un sistema de transporte para la vitamina D, la DBP constituye el lugar principal de almacenamiento de ésta [principalmente en forma 25(OH) vitamina D3 (ver apartado 4)]. La vitamina D puede ser también transportada en lipoproteínas plasmáticas. De hecho, el transporte en lipoproteínas o unido a la DBP depende de su origen: la vitamina D de origen endógeno se transporta unida a DBP, mientras que la de origen exógeno se transporta en quilomicrones y lipoproteínas. El medio de transporte determina entre otras cosas la velocidad con que la vitamina D es suministrada al hígado, siendo más rápida su captación cuando está unida a lipoproteínas. 3. Fotobiogénesis La fotobiogénesis es el proceso por el cual se obtiene vitamina D3 a partir del 7-deshidrocolesterol, un metabolito del colesterol producido en el hígado y exportado a la piel. En consecuencia, tanto los animales como el hombre pueden sintetizar vitamina D3, y simplemente con una exposición suficiente a la luz solar o a radiación ultravioleta-B (UV-B) se puede evitar la deficiencia de esta vitamina. Se calcula que la exposición de la cara y las manos a la luz solar durante 15 minutos 3 veces a la semana puede proporcionar cantidades adecuadas de vitamina D. La primera fase de la síntesis endógena de vitamina D3 se produce en los estratos germinativo y espinoso (capas basal y mucosa, respectivamente) de la piel, y consiste en la fotoconversión del 7-deshidrocolesterol (o provitamina D3) en previtamina D3 o precalciferol. En este proceso, la luz UV-B se absorbe por el anillo B del 7-deshidrocolesterol, produciéndose la ruptura del enlace 9,10. La tasa de fotoconversión depende tanto de la cantidad como de la calidad de la radiación que llega a estas capas de la epidermis. De hecho, las longitudes de onda requeridas son del orden de 290-315 nm aunque el máximo de conversión ocurre a 295 nm (Figura 1). Posteriormente, la previtamina D3 puede bien seguir transformándose en taquicolesterol y lumisterol mediante una nueva fotoconversión, o puede sufrir una isomerización química inducida por calor, obteniéndose la vitamina D3. El proceso de isomerización es un fenómeno que dura varios días (a la temperatura normal del cuerpo la isomerización del 50% de la previtamina D3 se produce en 28 horas y son necesarias 36 horas para que se transforme el 96% de la previtamina D3 en vitamina D3). Finalmente, la vitamina D producida en la epidermis llega al lecho dérmico capilar, desde donde es transportada al hígado unida a la DBP para iniciar su transformación metabólica. 4. Metabolismo hepático y renal de la vitamina D (Figura 1) La vitamina D3 que se concentra en el hígado es rápidamente hidroxilada en el carbono 25 por la enzima vitamina D3 25-hidroxilasa para obtener la 25(OH) vitamina D3. Esta reacción de hidroxilación se produce indistintamente sobre el calciferol (vitamina D3) y sobre el ergocalciferol (vitamina D2). La vitamina D3 25-hidroxilasa forma parte de un sistema enzimático dependiente de citocromo P-450, que se localiza principalmente en microsomas (aunque también se ha localizado un citocromo P-450 que cataliza esta actividad en mitocondrias hepáticas) y que requiere NADPH, oxígeno molecular e iones magnesio. Es interesante señalar que la vitamina D3 25hidroxilasa puede actuar también sobre la 1α-hidroxivitamina D2 y sobre la 1α-hidroxivitamina D3. Estos derivados de la vitamina D se usan frecuentemente en el tratamiento de distintas patologías renales. Una vez sintetizada, la 25(OH) vitamina D3 es enviada a la circulación sistémica, donde es la forma predominante de vitamina D3. De hecho, este metabolito es el que se determina cuando se estudian los niveles de vitamina D de un paciente. La 25(OH) vitamina D3 carece de actividad biológica, y ha de ser transportada al riñón, donde es nuevamente hidroxilada, para obtener los metabolitos activos: el calcitriol [1α,25(OH)2 vitamina D3] y el 24-R-calcitriol [24R,25(OH)2 vitamina D3]. Además de este destino metabólico, la 25(OH) vitamina D3 puede dar lugar a derivados más oxidados que son inactivos, o bien puede ser excretada por vía biliar, sufriendo un ciclo enterohepático. 795 Capítulo 1.24. Vitamina D Figura 1. Metabolismo de la vitamina D. La hidroxilación en riñón de la 25(OH) vitamina D3 es llevada a cabo por dos enzimas, la 25(OH) vitamina D3 1α-hidroxilasa y la vitamina D3 24-hidroxilasa, que se encuentran localizadas principalmente en las células del túbulo contorneado proximal. Ambas enzimas son muy parecidas a la vitamina D3 25-hidroxilasa, es decir, forman parte de un sistema enzimático en el que un citocromo P-450 cataliza la reacción de hidroxilación. No obstante, mientras que existe actividad 25(OH) vitamina D3 1α-hidroxilasa en mitocondrias y microsomas renales, sólo se ha detectado actividad vitamina D3 24-hidroxilasa en microsomas. 796 El calcitriol es producido principalmente por el riñón y, durante el embarazo, la placenta también secreta cantidades significativas de este metabolito. En los últimos años se ha descrito actividad 25(OH) vitamina D3 1α-hidroxilasa en varios tipos de células de la piel, del colon, de la próstata, etc. Aunque aún no se entiende bien la función de esta producción extrarrenal de calcitriol, se cree que que podría ser importante en el control del crecimiento y la diferenciación celular (ver apartado 11.2.1). De igual modo, además de en el riñón, existe actividad 25(OH) vitamina D3 24-hidroxilasa en la mucosa intestinal, el cartílago y otros tejidos que con- O. Martínez Augustin | V. Puerta Fernández | M.ª D. Suárez Ortega tienen receptores para el calcitriol. Posiblemente, esta actividad sea útil en la regulación de la actividad de esta forma de la vitamina D, ya que la 24-hidroxilación es la principal vía para su inactivación. El exceso de vitamina D3 se almacena en el tejido adiposo, al que llega transportada por la DBP, aunque, como ya se ha comentado anteriormente, el mayor depósito-almacén corporal de la vitamina D es el plasma. 5. Inactivación y excreción de la vitamina D La bilis es la principal vía de excreción de metabolitos de la vitamina D, aunque una cantidad muy pequeña puede ser excretada por la orina (menos del 5%). En la bilis sólo un 2-3% de la vitamina D está en forma de colecalciferol, 25(OH) vitamina D3 o calcitriol, siendo predominantes una serie de metabolitos hidroxilados y polares, y sus conjugados con ácido glucurónico. En la mayoría de los tejidos, la principal vía de inactivación del calcitriol se inicia con su 24-hidroxilación para después ser transformado mediante diversas oxidaciones, y en algunos casos conjugación con glucurónico, en compuestos más polares. Los compuestos que inducen hidroxilasas dependientes de citocromo P-450, como los barbitúricos y los anticonvulsivantes primidona y difenilhidantoína, provocan un incremento en la degradación de 25(OH) vitamina D3 y en la excreción de metabolitos de la vitamina D por vía biliar. Como resultado, el uso prolongado de anticonvulsivantes puede asociarse con el desarrollo de deficiencia de vitamina D. Además, los barbitúricos producen también la inducción de la 25-hidroxilasa incrementando, por tanto, la hidroxilación de la vitamina D3. 6. Control de la síntesis y degradación de vitamina D (Figura 2) Aunque el riñón produce las dos formas activas dihidroxiladas de la vitamina D3 (calcitriol y 24-Rcalcitriol), el calcitriol es el metabolito más activo; por tanto, no es de extrañar que su concentración circulante esté muy controlada. La enzima clave en la regulación es la 25(OH) vitamina D3 1α-hidroxilasa. La predominancia en la síntesis de una forma u otra de vitamina D3 viene determinada por los niveles circulantes de hormona paratiroidea (PTH) y por el estatus corporal de vitamina D. Así, cuando existe deficiencia de vitamina D o los niveles de calcio son bajos, se produce un incremento de la PTH que actúa: 1. Incrementando la transcripción de la 1α-hidroxilasa y, por tanto, su actividad, dando lugar a un incremento de la síntesis de calcitriol. 2. Inhibiendo la 24-hidroxilasa, disminuyendo por tanto la producción de 24-R-calcitriol y desplazando el equilibrio hacia la síntesis preferente de calcitriol. La inhibición de la 24-hidroxilasa se debe a que la PTH produce una disminución en el RNA mensajero de esta enzima al disminuir su vida media. Por el contrario, cuando el estatus de vitamina D es adecuado, los niveles de PTH son bajos y los de calcitriol altos, por lo que se produce un feedback negativo de este metabolito sobre la 25(OH) vitamina D3 1α-hidroxilasa, el cual reduce lógicamente los niveles de calcitriol. Además, el calcitriol es capaz de inducir la vitamina D3 24-hidroxilasa, lo que produce un incremento del 24-R-calcitriol, así como una inactivación del calcitriol mediante su transformación metabólica en 1α,24R,25(OH)3 vitamina D3. Se ha descrito que el calcitriol puede incrementar la expresión de la vitamina D3 24-hidroxilasa de dos formas diferentes: 1. Inducción rápida mediante la activación de factores de transcripción a través de la proteína kinasa C (PKC) y las proteína kinasas activadas por mitógenos (Mitogen Activated Protein Kinases, MAPK). 2. Inducción lenta mediante unión al receptor nuclear de vitamina D (VDRnuc1,25) (ver apartado 10.1). De hecho, la 24-hidroxilasa posee en su promotor dos elementos de respuesta al VDR. Este hecho es cierto para casi todos los tipos de células que se han estudiado excepto para los osteoblastos, en los que se ha descubierto que la PTH es capaz de aumentar la actividad de la 24-hidroxilasa. La regulación de la 25(OH) vitamina D3 1α-hidroxilasa por la PTH y por el calcitriol se produce 797 Capítulo 1.24. Vitamina D también a nivel de su expresión génica. De hecho, el promotor de esta enzima contiene tres sitios potenciales de unión de AMP cíclico (AMPc) y se ha demostrado que la PTH es capaz de incrementar la expresión de la enzima a través de un mecanismo dependiente de AMPc. Además, la PTH es capaz de activar a la proteína kinasa A y modificar la actividad de factores de transcripción fosforilándolos. Estos factores de transcripción inducirán la expresión de esta enzima. Por otra parte, el feed-back negativo del calcitriol sobre la 25(OH) vitamina D3 1α-hidroxilasa se produce mediante la unión del calcitriol al receptor nuclear de vitamina D (VDRnuc1,25) (ver más adelante), lo que produce la inhibición de la transcripción de la enzima. 7. Fuentes de vitamina D La principal fuente de vitamina D para la mayoría de los humanos es la endógena mediante exposición diaria a la luz del sol. Sin embargo, multitud de factores pueden disminuir e incluso suprimir la producción de vitamina D endógena. Así, la ingesta de calcio y fósforo, la edad, el sexo o la cantiFigura 2. Control de la síntesis y degradación de la vitamina D. dad de pigmentación de la piel pueden influir en la síntesis de vitamina D. Además, el grado de exposición al sol, luz solar son factores que pueden contribuir a que que varía con la aplicación tópica de pantallas sola piel encuentre dificultades para la síntesis de vilares, la contaminación atmosférica, la tendencia a tamina D3. vivir en ciudades cuyos elevados edificios pueden De los factores comentados, el que tiene más imobstaculizar la radiación del sol, la tendencia a haportancia general en la producción de vitamina D3 bitar en interiores o vivir en regiones geográficas es el ángulo de incidencia de la radiación solar. Por del mundo que no reciben suficiente cantidad de tanto, la latitud, la estación del año y la hora del día 798 O. Martínez Augustin | V. Puerta Fernández | M.ª D. Suárez Ortega afectan a la síntesis de vitamina D3. Así, cuanto más lejos se esté del ecuador, menor es la porción del año en la que la radiación solar es suficiente para la fotoconversión del 7-deshidrocolesterol. Esto es debido a que las ondas UV-B procedentes del sol son absorbidas cuando pasan a través de la atmósfera; a mayor latitud el ángulo de los rayos del sol es mayor y por tanto el camino a través de la atmósfera es más largo y llegará menos UV-B a la superficie de la tierra. Por ejemplo, en Vigo (42,14 ºN) el ángulo de incidencia del sol es tan oblicuo en invierno que muy pocos fotones de radiación UV-B con la energía necesaria llegan a la superficie de la tierra. Por tanto, en Vigo durante los meses de noviembre a febrero se sintetizará muy poca vitamina D3 en la piel. Por el contrario, en Algeciras el ángulo es menos oblicuo a lo largo del año, por lo que la producción de vitamina D3 ocurrirá durante casi todo el año. Como regla general, en latitudes por encima de 40 grados al norte y al sur del ecuador la producción de vitamina D3 en la piel está significativamente disminuida o es inexistente durante el invierno. La vitamina D3 es particularmente abundante en productos animales, concretamente en los pescados marinos grasos, como los arenques, el salmón o las sardinas. También se encuentra en aceites de hígado de pescado como el de hígado de bacalao. Además, los huevos, la carne bovina, la mantequilla y los aceites vegetales contienen pequeñas cantidades de vitamina D3 (Tabla 2), mientras que las plantas, las frutas y los frutos secos son muy pobres en esta vitamina. En todos los casos, la vitamina D presente en los alimentos es estable y no es destruida por el calor ni por procesos tecnológicos. En general, cuando las vitaminas son adicionadas a alimentos de uso general, como la leche, el pan o la margarina, con objeto de asegurar una ingesta adecuada, se habla de fortificación. Por el contrario, cuando las vitaminas se añaden para restaurar las pérdidas ocurridas durante el procesado, se habla de enriquecimiento vitamínico. Varios países, incluyendo Estados Unidos, Canadá y algunos países de Europa, fortifican con vitamina D alimentos como la leche, la margarina, los cereales, algunos panes y pastas o el zumo de naranja. En España, gran parte de la leche desnatada o semidesnatada se enriquece con vitaminas A y D. La adición de complejos multivitamínicos es obligatoria para algunos productos utilizados durante largo tiempo como nutriente único por al- gunas poblaciones; tal es el caso de las fórmulas lácteas, los cereales infantiles y los productos para nutrición enteral y parenteral de uso predominantemente hospitalario, además de la dietas de bajo valor energético para reducción de peso. La Tabla 3 recoge las recomendaciones de la Unión Europea para la suplementación con vitamina D3 de distintos preparados. Estas recomendaciones son de obligado cumplimiento para todos los Estados miembros. En general la vitamina D se adiciona a los alimentos en forma de ergocalciferol (vitamina D2) obtenido mediante radiación del ergosterol. Éste, a su vez, se obtiene por vía microbiológica en biorreactores. 8. Requerimientos nutricionales de vitamina D Debido a que la principal fuente de vitamina D es la derivada de la síntesis endógena en la piel, la determinación de los requerimientos o del aporte recomendado de vitamina D es difícil, ya que cuando la radiación solar es suficiente la vitamina D exógena es innecesaria. Además, los requerimientos de vitamina D varían en función de los parámetros antes comentados (ingesta de calcio y fósforo, edad, sexo, grado de exposición al sol, cantidad de pigmentación de la piel, etc.). La Organización Mundial de la Salud define la unidad internacional (UI) de vitamina D3 como la actividad vitamínica de 0,025 mg de la preparación de referencia internacional de vitamina D cristalizada. Es decir, 1 UI de vitamina D3 equivale a 0,025 mg, lo que, a su vez, equivale a 65 pmol. Cuando se descubrió el metabolismo de la vitamina D3 se recomendó que una unidad de calcitriol fuera considerada como el equivalente molar de una unidad de vitamina D3. Por tanto, una unidad de 1α,25(OH)2 vitamina D3 equivale a 65 pmoles. En 1997 el Comité de Alimentos y Nutrición del Instituto de Medicina (Institute of Medicine, IOM) de la Academia Nacional de Ciencias Americana instituyó las siguientes ingestas diarias recomendadas de vitamina D. 1. De 0 a 1 año. En la leche materna hay niveles relativamente bajos de vitamina D, excepto cuando se suplementa la dieta de la madre con 799 Capítulo 1.24. Vitamina D Tabla 2. CONTENIDO EN VITAMINA D DE DISTINTOS ALIMENTOS (μg/100 g) Cereales Semillas, harinas, almidones 0 Leche y productos lácteos Leche de vaca 0,01-0,03 Leche humana 0,04 Leche en polvo 0,21 Nata 0,1-0,28 Queso 0,03-0,5 Yogur Trazas-0,04 Huevos Completos 1,75 Yema 4,94 Grasas y aceites Mantequilla 1,00-3,00 Aceite de hígado de bacalao 330,00 Carne y productos cárnicos Ternera, vaca, cerdo, cordero Trazas Pollo, gallina Trazas Hígado 0,2-1,1 Pescado Pescado blanco Trazas Pescado graso Trazas-27,00 Salmón 16,00 Arenque 27,00 Anguila 20,00 Crustáceos o moluscos Trazas Vegetales 0 grandes dosis de vitamina D, por lo que los niños alimentados al pecho son propensos a desarrollar deficiencia de vitamina D. Dado que las fórmulas infantiles están normalmente suplementadas con vitamina D, los niños alimentados de esta forma suelen recibir cantidades adecuadas de esta vitamina. El IOM recomienda como ingesta adecuada diaria para niños 200-400 UI/día, aunque se ha demostrado que ingestas de 100 UI/día son suficientes para prevenir la aparición de raquitismo en niños que se exponen frecuentemente al sol. 2. De 1 a 18 años. Los niños y adolescentes necesitan la vitamina D para el desarrollo óptimo 800 del esqueleto y la mineralización ósea. La mayoría de los niños reciben suficiente radiación solar, lo que ayuda a asegurar niveles adecuados de vitamina D. No obstante, los niños con alta pigmentación cutánea y aquellos que reciben poca luz solar son grupos de riesgo de deficiencia en vitamina D. La IOM aconseja una ingesta diaria de 200 UI/día para niños que están adecuadamente expuestos a la luz solar, y de 400 UI/día para el resto. 3. De 19 a 50 años. La IOM aconseja una ingesta diaria adecuada de 200 UI/día. Se considera que una ingesta de 400 UI/día sería también razonable y no causaría ningún efecto adverso. O. Martínez Augustin | V. Puerta Fernández | M.ª D. Suárez Ortega Tabla 3. RECOMENDACIONES DE LA UNIÓN EUROPEA PARA LA SUPLEMENTACIÓN CON VITAMINA D DE DISTINTOS PREPARADOS (μg/100 kcal) Mínimo Máximo Preparados para lactantes 1 2,5 Preparados de continuación para lactantes 1 3 Alimentos de uso médico especiales para lactantes 1 2,5 Alimentos de uso médico especiales para niños y adultos 0,5 3 4. De 51 a 70 años. La ingesta diaria recomendada para este grupo de edad es de 400 UI/ día. Esta ingesta puede ser incrementada hasta 600 UI/día en individuos que no están expuestos a la luz solar. 5. Mayores de 71 años. La edad disminuye la capacidad de producir vitamina D3 porque disminuye la concentración de su precursor, el 7-deshidrocolesterol; además, se calcula que la capacidad de producir vitamina D3 en la piel de una persona de 65 años es 3-4 veces menor que la capacidad de la piel de un adulto joven saludable. A estos hechos hay que añadir que la edad disminuye la capacidad renal de hidroxilación de la 25(OH) vitamina D3. Los ancianos de países industrializados son, por tanto, un grupo de riesgo de deficiencia de vitamina D. En función de lo anteriormente expuesto, el Comité de Alimentos y Nutrición triplicó en 1997 la ingesta adecuada establecida en 1989 para individuos mayores de 70 años (de 200 UI a 600 UI). 6. Embarazo y lactancia. Aunque cabría pensar que el embarazo y la lactancia deberían incrementar los requerimientos de vitamina D, no existen datos suficientes en la literatura que avalen esta hipótesis. Por tanto, se considera la ingesta adecuada durante la lactancia y el embarazo de 200 UI/día. No obstante, podría ser razonable incrementar esta cantidad hasta 400 UI/día, que es la cantidad proporcionada en los suplementos dietéticos prenatales. 8.1. Ingesta máxima tolerada de vitamina D Aunque el consumo excesivo de vitamina D en la dieta es poco probable, a menos que se con- suman grandes cantidades de hígado de bacalao, no es descartable que se produzcan ingestas altas procedentes de suplementos vitamínicos. El Comité de Alimentos y Nutrición del Instituto de Medicina Americano considera que una ingesta de 1.000 UI/día para niños menores de 12 meses y de 2.000 UI/día para niños mayores de 1 año, adultos, embarazadas o madres lactantes puede ser la ingesta máxima tolerada. Dosis excesivas de vitamina D (1.000 UI/día en niños y 50.000 UI/día en adultos, más de 25 veces las dosis usuales en adultos) producen hipercalcemia y, como consecuencia de ésta, el calcio se deposita en los tejidos blandos como el riñón y el cerebro, con producción de hipertensión arterial e insuficiencia renal. La administración continuada de dosis tóxicas puede incluso producir la muerte. Los síntomas de sobredosis moderada de vitamina D pueden incluir náuseas, vómitos, anorexia, fatiga, poliuria y cefaleas. Además, el exceso de vitamina D en sangre puede producir cambios en el estado mental como confusión. 9. Mecanismo de acción de la vitamina D 9.1. Receptores de vitamina D (Figura 3) Los metabolitos de la vitamina D (el calcitriol y el 24-R-calcitriol) producidos en el túbulo proximal del riñón se unen a la DBP para ser transportados hasta los órganos diana de la vitamina D. Estos órganos diana están definidos por la presencia de tres receptores distintos. Así, el calcitriol [1α,25(OH)2 vitamina D3] se puede unir al 801 Capítulo 1.24. Vitamina D Figura 3. Receptores de la vitamina D. receptor nuclear de vitamina D (Vitamin D Receptor, VDRnuc) y al receptor de membrana de vitamina D (VDRmem1,25). Por su parte, el 24R-calcitriol [24R,25(OH)2 vitamina D3] se une a otro receptor de membrana denominado VDRmem24,25. En general, la función del receptor nuclear está relacionada con la respuesta genómica a nivel transcripcional, mientras que los receptores de membrana median las llamadas respuestas biológicas rápidas (no transcripcionales) de la vitamina D, que implican la estimulación de cascadas de transducción de señal. De hecho, estos receptores 802 de membrana pertenecen a un grupo de proteínas o complejos de unión a esteroides asociados a membranas que median respuestas biológicas rápidas [Membrane Associated, Rapid Response Steroid (MARRS) binding proteins or complexes]. Una de las características más importantes del calciferol es su capacidad de generar una gran variedad de conformaciones al ser una molécula muy flexible. Así, la unión del calcitriol al receptor de membrana o al receptor nuclear depende de la capacidad de este ligando de generar diferentes formas que satisfacen específicamente los requerimientos de cada receptor. O. Martínez Augustin | V. Puerta Fernández | M.ª D. Suárez Ortega 9.1.1. Receptor de membrana del 24-R-calcitriol (VDRmem24,25) La distribución tisular del receptor VDRmem24,25 es muy específica, ya que se limita a las células del hueso y el cartílago, y posiblemente a las células secretoras de la hormona paratiroidea. Los efectos biológicos del 24-R-calcitriol han sido estudiados principalmente en animales y en células en cultivo. Los efectos más sobresalientes incluyen, a nivel de glándulas paratiroideas, la inhibición de la secreción de la hormona PTH y la regresión de glándulas paratiroideas hipertrofiadas en animales hipocalcémicos deficientes en vitamina D. En relación con el hueso, se ha descrito en pollos que la administración local de 24-R-calcitriol puede curar las lesiones producidas por el raquitismo y que esta forma de vitamina D3 interviene en la curación de fracturas. De hecho, varios días después de producirse una fractura se ha detectado que la actividad 24R-hidroxilasa renal se encuentra incrementada en modelos de fracturas óseas que utilizan pájaros y pollos. Además, en varios modelos animales se ha observado que esta vitamina incrementa la masa ósea e incrementa la resistencia del hueso en conejos. Por último, se ha descrito que el 24-R-calcitriol produce la activación de condrocitos de la zona de descanso en cultivo (ver apartado 11.1.2). Las vías de señalización desde el receptor VDRmem24,25 están comenzando a ser estudiadas y ya se ha descrito que en la activación de estas células de la zona de descanso del cartílago por el 24-R-calcitriol interviene la activación de la proteína kinasa D (PKD) asociada al VDRmem24,25. La activación de esta proteína kinasa produce la inactivación de la PKC y de MAPK, produciendo efectos a nivel genómico y no genómico. 9.1.2. Receptor de membrana del calcitriol (VDRmem1,25) La idea de que algunas de las acciones del calcitriol debían estar mediadas por un receptor de membrana fue propuesta originalmente en 1984. Posteriormente, en 1994 se aisló una proteína de unos 60 kDa a la que se unía este metabolito. Desde entonces, varios laboratorios han mos- trado pruebas de la existencia de este receptor en distintos tipos de células, aunque hasta la fecha la proteína correspondiente al VDRmem1,25 no ha sido aislada, por lo que debe ser denominada putativa. Al contrario que el VDRmem24,25 el VDRmem1,25 es un receptor ampliamente distribuido en gran variedad de tejidos donde media gran cantidad de respuestas biológicas denominadas rápidas, mediante la estimulación de una gran variedad de sistemas de señalización celular. Los principales efectos de la respuesta rápida inducida por el calcitriol son la estimulación de la absorción de calcio tanto a nivel intestinal como en osteoblastos además de la inhibición de la proliferación celular y la inducción de la diferenciación en distintos tipos de células como osteoblastos, keratinocitos y colonocitos. El calcitriol actúa vía VDRmem1,25, produciendo un incremento rápido (en minutos) en la absorción de calcio y en la concentración intracelular del mismo, mediante el incremento de su entrada a través de canales de calcio y mediante la liberación del calcio intracelular. Estos mecanismos están relacionados, ya que la entrada de calcio produce la hidrólisis de lípidos de membrana mediada por proteínas G, fosfolipasa C y fosfolipasa D, produciendo la liberación de segundos mensajeros (IP3 y/o DAG), que a su vez producen la liberación del calcio de los depósitos intracelulares y la activación de cascadas de señalización intracelular mediadas por PKC y MAPK (ver Capítulo 1.5). El incremento de la entrada de calcio podría estar relacionado con un incremento de las actividades de la adenilato ciclasa y de la proteína kinasa A (Figura 4). Por otra parte, el calciferol es capaz de activar la PKC por unión directa a la enzima y mediante la unión a sus receptores de membrana, que a su vez se encuentran asociados a fosfolipasa y PKC (Figura 4); en respuesta al calcitriol se producen cambios rápidos en la liberación de ácido araquidónico y en su reincorporación a los fosfolípidos de la membrana, además de incrementos en la síntesis de prostaglandinas E1 y E2. Las MAPK pertenecen a la familia de proteínas con actividad serina/treonina kinasa que contienen serina o treonina y que pueden ser activadas, mediante fosforilación de un residuo de treonina, por mitógenos o agentes promotores de la 803 Capítulo 1.24. Vitamina D Figura 4. Mecanismo de acción del receptor de membrana del calcitriol. diferenciación celular (ver Capítulo 1.32). Estas kinasas forman parte de las vías de transducción de señal mediadas por varios segundos mensajeros, entre ellos la PKC, que regulan muchas funciones celulares mediante la fosforilación de kinasas citoplasmáticas y la regulación de factores nucleares de transcripción como el receptor del factor de crecimiento epidérmico u oncogenes como c-myc y c-jun. El calcitriol produce la activación de MAPK. Recientemente se ha postulado que estas kinasas podrían jugar un papel importante en la transducción de señal mediada por el calcitriol desde la membrana al núcleo. De este modo, el calcitriol mediante su acción rápida podría modular las repuestas genómicas que tienen como efecto final la inhibición de la proliferación celular y la inducción de la diferenciación. 804 9.1.3. Receptor nuclear del calcitriol (VDRnuc1,25) El receptor nuclear de vitamina D (vitamin D receptor, VDRnuc1,25) pertenece a la superfamilia de factores de transcripción activados por ligandos, con los que comparte mecanismo de acción y estructura (ver Capítulo 1.7). A esta superfamilia de receptores pertenecen también los receptores nucleares de estrógenos, andrógenos, mineralocorticoides, glucocorticoides, hormonas tiroideas y vitamina A. La activación de estos factores de transcripción se produce mediante la unión de ligandos específicos que, por su pequeño tamaño y por su naturaleza lipofílica, son capaces de difundir a través de la membrana celular. Una vez unido al ligando, cambia la conformación del receptor que O. Martínez Augustin | V. Puerta Fernández | M.ª D. Suárez Ortega el ligando del RXR es el ácido todo-trans-retinoico (Figura 5). El elemento de respuesta al que se une el dímero VDRRXR se denomina elemento de respuesta a vitamina D (Vitamin D-Responsive Element, VDRE) y está constituido por dos repeticiones directas imperfectas de una secuencia de hexanucleótidos separadas por tres nucleótidos. Por tanto, la secuencia consenso del VDRE sería la siguiente: 5´-GGGTCA-NNN-GGTTCA-3´. Obviamente, los promotores de los genes modulados por esta vitamina poseen VDRE que contienen variaciones en esta secuencia consenso (Figura 5). 9.1.3.1. El gen del VDRnuc1,25 humano (hVDRnuc1,25) Es bien conocido que mutaciones específicas en el DNA del hVDRnuc1,25 causan raquitismo resistente a vitamina D, una enfermedad autosómica recesiva poco frecuente, también conocida como raquitismo tipo II dependiente de vitamina D (ver apartado 13.4). Se han descriFigura 5. Mecanismo de acción del receptor nuclear del calcitriol. to distintas mutaciones en la secuencia de DNA del hVDRse dimeriza y se une en el núcleo a secuencias esnuc1,25 que son responsables de esta enfermedad pecíficas, denominadas elementos de respuesta, siy que afectan tanto a la unión del receptor a su lituadas típicamente en los promotores cuya transgando como a la localización nuclear del complejo cripción van a modular (Figura 5). hormona-receptor, a la unión del receptor a su seLa dimerización del VDRnuc1,25 se produce con cuencia diana o a la unión del receptor a un coactiel receptor X del ácido retinoico (Retinoic X Revador. De hecho, recientemente se ha demostrado ceptor, RXR) perteneciente también a la superfaen pacientes con mutaciones en el VDR que afecmilia de factores de transcripción con respuesta a taban al dominio de unión al ligando que el trataligandos y es necesaria para que la unión del VDRmiento con análogos del calcitriol puede en parte nuc1,25 a su elemento de respuesta sea de alta afio totalmente restaurar la respuesta del VDR mutanidad. Como se ha indicado anteriormente, el lido, sugiriendo la posibilidad de tratar a pacientes gando del VDRnuc1,25 es el calcitriol, mientras que seleccionados con estos análogos. 805 Capítulo 1.24. Vitamina D Figura 6. Polimorfismos funcionales del gen humano del receptor nuclear del calcitriol. Además, se ha descrito que existen otras variaciones en los alelos de este gen que se producen con mucha más frecuencia y podrían influir en el metabolismo del calcio y el fósforo y predisponer genéticamente al padecimiento de ciertas enfermedades, especialmente enfermedades óseas como la osteoporosis. Estudios llevados a cabo al principio de la década de los 90 del siglo pasado demostraron la relación génetica entre el receptor VDR y la densidad mineral ósea. En estos estudios se observó que los niveles séricos de osteocalcina se correlacionan positivamente con la densidad ósea y dependen de variaciones alélicas en el gen del VDR. La síntesis de osteocalcina, la proteína de tipo no colágeno más abundante en el hueso, es inducida por unión del complejo calcitriolhVDRnuc1,25 a una secuencia VDRE presente en el promotor de este gen. Además, en 1992 un estudio describió, en un grupo de gemelos australianos monocigotos y heterocigotos, que la existencia de una variación polimórfica en la región 3’ del gen hVDRnuc1,25 podía suponer variaciones de hasta 806 el 75% en la densidad mineral ósea. Las mutaciones descritas en este trabajo inicial eran mutaciones en un intrón o mutaciones silenciosas, por lo que actualmente se piensa que los polimorfismos descritos pueden servir como marcadores de otro polimorfismo funcional en el gen del hVDR o de un gen próximo. Hasta ahora se han descrito tres posibles polimorfismos funcionales en el gen del hVDR (Figura 6). El primero se encuentra situado en la región 3’ no traducida del RNA mensajero de este receptor y consiste en la existencia de una región poli-A situada aproximadamente 1 kb corriente arriba de la señal de poliadenilación. Se han descrito variaciones alélicas múltiples (alrededor de 12) en la longitud de esta secuencia que podrían afectar a la estabilidad o eficiencia de traducción de este RNA mensajero. En función de su longitud estas variaciones han sido clasificadas como largas (L) y cortas (S), y varios estudios han demostrado que el alelo L del hVDR es más activo en fibroblastos humanos. O. Martínez Augustin | V. Puerta Fernández | M.ª D. Suárez Ortega Un segundo polimorfismo se ha localizado en el sitio de inicio de la traducción del gen del hVDRnuc1,25. En este caso, la presencia de una sustitución (T por C) en el exón 2 elimina el sitio de inicio de la traducción (el primer ATG), por lo que se utiliza el segundo situado a 9 pares de bases. Por tanto, aparecen dos proteínas distintas de VDRnuc1,25 que diferen en 3 aminoácidos y que han sido denominadas M1 y M4, según contengan 427 o 424 aminoácidos respectivamente. Se ha demostrado que la isoforma M4 se traduce más activamente que la M1, probablemente porque el mRNA interaccione mejor con la proteína TFIIB del complejo basal de inicio de la traducción. Este polimorfismo puede ser detectado mediante la técnica de polimorfismo de longitud de fragmentos de restricción (Restriction Fragment Length Polymorphysm, RFLP), ya que la presencia de un sitio de corte para la enzima de restricción Fok-I indica que es la isoforma M1 la que está siendo expresada, mientras que este sitio de corte se pierde cuando es la isoforma M4 la que se expresa. Por último, se ha detectado un tercer polimorfismo con posible relevancia funcional entre dos exones que codifican parte de la región 5’ no traducida del gen del hVDRnuc1,25. En este caso, se produce una sustitución de G por A en el elemento de unión de un factor de transcripción denominado Cdx-2. Se ha observado que la presencia del nucleótido de adenina en esta posición está relacionada con una mayor actividad del VDRnuc1,25. En relación con esta observación, se ha descrito una menor densidad mineral ósea en mujeres japonesas, postmenopáusicas y homocigotas GG, coincidente con una menor capacidad de transactivación del alelo G. Estos hechos podrían explicarse teniendo en cuenta que la presencia del nucleótido de adenina hace que la homología con el sitio de unión a DNA del Cdx-2 sea mayor. En resumen, podría decirse que está claro que existen múltiples variaciones polimórficas en el gen del VDRnuc1,25 que tienen distintos tipos de consecuencias. Así, las alteraciones descritas en el promotor y el extremo 5’ pueden afectar a los niveles de expresión del gen, además de a la eficiencia de la traducción, mientras que los polimorfismos descritos en el extremo 3’ no traducido pueden afectar a la estabilidad del RNA mensajero y/o a la eficiencia de la traducción de la proteína. La combinación de estas diferencias genotípicas daría lugar a variacio- nes en los niveles de la proteína del VDRnuc1,25 y/ o en su función, dependiendo del individuo, del tipo de célula, del estado de desarrollo y del estado de activación. No obstante, es necesario describir mejor la función de estos polimorfismos y su asociación con funciones biológicas y con la etiología de enfermedades como la osteoporosis. Además, es muy probable que en los próximos años se describan más polimorfismos funcionales en este gen. Además de los polimorfismos y de las mutaciones descritos previamente, la variación genética en la expresión del VDRnuc1,25 y su expresión específica en distintos tejidos puede ser explicada teniendo en cuenta que este gen tiene un promotor muy extenso capaz de generar múltiples tránscritos mediante procesos alternativos de corte y empalme (splicing alternativo) a nivel del extremo 5’. Dos de estos tránscritos contienen sitios alternativos de inicio de la traducción, por lo que potencialmente podrían dar lugar a la presencia de aminoácidos adicionales en la proteína del hVDRnuc1,25. No obstante, será necesario definir en un futuro la funcionalidad de estas isoformas así como su distribución tisular. 9.1.3.2. Estructura proteica del VDR humano y su regulación mediante fosforilación En cuanto a su estructura, el VDRnuc1,25 humano (hVDRnuc1,25), al igual que los demás receptores de esta superfamilia, posee en su estructura primaria diferentes dominios que reflejan a nivel estructural las acciones del receptor en respuesta a su ligando. Así, este receptor posee un dominio de unión a DNA en su extremo amino terminal, un dominio de unión al ligando en su extremo carboxilo terminal y un dominio de heterodimerización con el RXR que se encuentra disperso en subregiones del dominio de unión a DNA y del dominio de unión al ligando (Figura 7). El dominio de unión a DNA es el más conservado entre las distintas especies en la superfamilia de receptores nucleares e incluye dos motivos del tipo dedo de zinc, concretamente del tipo cys4. En este dominio se encuentran además tres agrupaciones de residuos básicos implicados en la localización nuclear del VDRnuc1,25. Por su parte, el dominio de unión al ligando es un dominio multifuncional, ya que está implicado no sólo en la unión al ligando y en la dimerización, sino también en la 807 Capítulo 1.24. Vitamina D Figura 7. Estructura proteica y regulación mediante fosforilación del VDR nuclear humano. activación de la transcripción (transactivación). Concretamente, este dominio contiene un motivo denominado función de transactivación dependiente del ligando o función de activación 2 (Activating Function 2, AF2), formado por un residuo de glutámico (E420) flanqueado por residuos hidrofóbicos, que es útil en la interacción con coactivadores que son proteínas que regulan la transcripción (ver apartado 10.1.3.3). Las funciones del hVDRnuc1,25 (localización nuclear, unión al DNA y activación de la transcripción) se modulan intracelularmente por fosforilación/ desfosforilación de serinas. En este sentido, el hVDRnuc1,25 es muy parecido a los receptores nucleares de glucocorticoides, de estrógenos, de la hormona tiroidea y del ácido retinoico, que también son regulados de esta manera. De las serinas presentes en este receptor, la serina-208 es la más fosforilada en respuesta al calcitriol, y se ha demostrado que la fosforilación de este residuo mantiene al hVDRnuc1,25 en un estado conformacional más activo para interaccionar con coactivadores o componentes del aparato basal de la transcripción. Por tanto, la fosforilación en ser-208 estimula la transactivación. Aparentemente, es la enzima proteína kinasa CKII la que cataliza la reacción de fosforilación de este residuo. 808 La ser-51 presente en el dominio de unión a DNA es el segundo residuo de serina más fosforilado del hVDRnuc1,25 e influye no sólo en la transactivación, sino también en la localización nuclear del factor de transcripción y en su unión al DNA. Se ha descrito que su fosforilación está catalizada por la proteína kinasa C-β (PKC-β). Esta fosforilación no depende de la presencia del ligando, y disminuye la capacidad de unión al VDRE. Aunque la fosforilación de estos dos residuos (ser-208 y ser-51) supone el 90% del total de las fosforilaciones del hVDRnuc1,25, se ha descrito que la fosforilación producida por proteína kinasa dependiente de AMP cíclico (proteína kinasa A, PKA) entre los aminoácidos 134 y 201 podría estar relacionada con la regulación de la transactivación estimulada por el calcitriol. La existencia de diferentes sitios en el hVDRnuc1,25 regulados mediante fosforilación, y el hecho de que estén regulados independientemente y por distintas cascadas de kinasas iniciadas en la superficie celular, indica que las funciones nucleares del calcitriol están finamente reguladas no sólo por la unión de este ligando al hVDR, sino también por hormonas y factores de crecimiento y por el estado celular de crecimiento y diferenciación. O. Martínez Augustin | V. Puerta Fernández | M.ª D. Suárez Ortega 9.1.3.3. Factores implicados en la regulación transcripcional por calcitriol La regulación de la transcripción por factores de transcripción se lleva a cabo mediante la unión de los mismos al promotor del gen regulando la tasa de transcripción de la RNA polimerasa II (ver Capítulo 1.7). Para que el factor de transcripción (que frecuentemente se une a regiones del promotor lejanas al punto de inicio de la transcripción) pueda alterar la tasa de transcripción de la RNA polimerasa, es necesaria la existencia de una serie de coactivadores que actúan de nexo entre el factor de transcripción y el aparato basal de la transcripción. Estos coactivadores pueden activar o inhibir la transcripción mediante varios mecanismos que incluyen: 1. La remodelación de la cromatina, que como ya se ha dicho hace que sea más accesible el gen para el aparato basal de la transcripción. 2. El reclutamiento de proteínas de la maquinaria basal de la transcripción. 3. El mantenimiento de la estabilidad del complejo basal de inicio de la tanscripción. De hecho, recientemente se ha descrito la secuencia de acontecimientos que llevan al inicio de la transcripción: en primer lugar es necesaria la unión de coactivadores que remodelan la cromatina, para a continuación comenzar a reclutar componentes del aparato basal que incluyen factores de transcripción generales (TFIIA, TFIIB, TFIID, TFIIE,TFIIH) y la RNA polimerasa II. Parece ser que se necesitan ambos tipos de coactivadores (los que remodelan la cromatina y los que reclutan la polimerasa II) o factores con ambas actividades para que la tanscripción sea eficiente. La unión al VDRnuc1,25 produce, además de la heterodimerización con el RXR, el reclutamiento de varias proteínas coactivadoras entre las que se han incluido hasta la fecha proteínas de la familia de coactivadores de receptores esteroideos (Steroid Receptor Coactivators, SRC), el complejo de proteínas que interaccionan con el VDR (VDR Interacting Protein Complex) o DRIP/TRAP, el complejo CBP/P300 y las proteínas coactivadoras P160, SMAD3 o NcoA-62. El SRC se une al dominio AF-2 y a otros puntos de VDRnuc1,25, mientras que otros coactivadores como el NcoA-62 no necesitan el dominio AF-2 para unirse al VDRnuc1,25. Se ha descrito que el hecho de que distintos coactivadores se unan a distintas secuencias del VDRnuc1,25 puede hacer que se produzca un efecto sinérgico con el fin de incrementar la transcripción activada por el ligando (el calcitriol). 10. Acciones de la vitamina D (Figura 8) 10.1. Acciones clásicas de la vitamina D. Homeostasis mineral La vitamina D3 participa de manera activa en el mantenimiento de la concentración circulante de calcio actuando sobre la absorción intestinal de calcio, sobre la síntesis y degradación del hueso y sobre la excreción renal de calcio. Como se ha comentado anteriormente, una disminución de la concentración sérica de calcio estimula la liberación de PTH, que a su vez estimula la síntesis de calcitriol (Figura 2). Además, el calcitriol, junto con la PTH, estimula la reabsorción renal de calcio y la movilización de calcio del hueso (resorción ósea). Los efectos intestinales de la PTH están mediados por el calcitriol, mientras que ambas moléculas tienen actividad directa sobre el riñón y el hueso. El incremento de los niveles séricos de calcio produce la inhibición de la secreción de la PTH, disminuyendo como consecuencia la biosíntesis de calcitriol y la movilización de calcio. Además, cuando los niveles plasmáticos de calcio suben por encima de lo normal, las células C del tiroides secretan la hormona calcitonina que bloquea la movilización de calcio del hueso y posiblemente estimula la excreción de calcio y fósforo en el riñón, volviendo a la normalidad los niveles de calcio. La PTH actúa en cuestión de minutos, mientras que la estimulación del calcitriol requiere muchas horas. Por tanto, la regulación a corto plazo del control del calcio circulante depende de la acción de la PTH en el riñón y el hueso, con ayuda del calcitriol existente, mientras que una hipocalcemia prolongada provocará un incremento del calcitriol estimulando la absorción de calcio intestinal. A continuación, se discutira más en profundidad la acción del calcitriol sobre sus tres órganos diana tradicionales: el intestino, el hueso y el riñón. 809 Capítulo 1.24. Vitamina D Figura 8. Acciones de la vitamina D. 10.1.1. Intestino Es bien conocido que la administración de calcio intravenoso o el tratamiento a largo plazo con dosis orales de calcio producen la curación de niños que sufren raquitismo severo. Por otra parte, las anormalidades óseas producidas por deficiencias graves de vitamina D son prácticamente normalizadas mediante la infusión prolongada de calcio. Además, los ratones en los que el gen VDRnuc1,25 ha sido eliminado (ratones knockout para VDR) nacen normales fenotípicamente y sólo desarrollan hiperparatiroidismo, hipocalcemia, osteomalacia y raquitismo una vez son destetados. No obstante, la alimentación de estos ratones con una dieta con altos niveles de calcio, fósforo y lactosa normaliza los niveles de calcio y de hormona paratiroidea, a la vez que previene la aparición de raquitismo y osteomalacia. Estos hechos sugieren que el principal efecto del calcitriol sobre la 810 homeostasis del calcio se produce sobre la absorción intestinal de calcio. En general, la absorción neta de calcio es el resultado del balance entre la absorción intestinal de calcio vía transcelular (saturable) y vía paracelular (no saturable), y la secreción de calcio por diferentes órganos en el intestino (secreción gástrica, biliar, pancreática e intestinal). A su vez, el transporte neto intestinal de calcio está determinado por la disponibilidad de este ión en la dieta, por su solubilidad en el intestino y por la capacidad neta de absorberlo a través del intestino. El transporte paracelular de calcio es un proceso pasivo no saturable que depende de la concentración luminal de calcio y de la integridad de las tight junctions. Por el contrario, el transporte transcelular de calcio es un proceso saturable mucho más regulado, que es funcionalmente importante en condiciones de baja ingesta o deficiencia de calcio. En este caso, es necesario captar el máximo O. Martínez Augustin | V. Puerta Fernández | M.ª D. Suárez Ortega En las células intestinales se dan dos respuestas distintas a la administración de calcitriol. Por una parte, se produce una respuesta rápida que incrementa la absorción de calcio. Esta respuesta se produce gracias al reclutamiento hasta la membrana apical del enterocito de canales de calcio presintetizados. Hasta la fecha se han descrito dos canales epiteliales de calcio denominados ECaC1 (Epithelial Calcium Channel) y ECaC2 (en el intestino predomina éste, mientras que en el riñón predomina el EcaC1), además de un canal transportador de calcio denominado CaT1 (Calcium Transport channel), que median la entrada de Figura 9. Mecanismo de acción de la vitamina D a nivel intestinal. calcio en el enterocito y son sensibles a la acción del calcitriol. posible de calcio del lumen intestinal; la absorción Por otra parte, se produce una respuesta lenta paracelular no será suficiente y se necesitará la consistente en la síntesis de más canales de calcio, ayuda de la absorción transcelular. Por el contrario, de calbindina (encargada del transporte de calcio cuando la ingesta de calcio sea alta (alta concentraen el enterocito) y de transportadores de calcio ción luminal de calcio), la vía transcelular no será que se encargan de la extrusión del calcio desde la importante, ya que se absorberá suficiente calcio membrana basolateral del enterocito. De hecho, se por la vía paracelular. ha demostrado que los ratones a los que se elimina El transporte transcelular se da principalmente el gen del VDRnuc1,25 presentan una expresión de en el duodeno y en la porción proximal del yeyuno los canales ECaC2 y ECaC1 muy disminuida (del y consta de tres fases: orden del 90%) en comparación con ratones nor1. La entrada de calcio a través de la membramales (wild type). na del borde en cepillo mediante canales de calcio Entre otras funciones, la calbindina es una proespecíficos. teína que se une directamente al calcio y actúa co2. El transporte intracelular. mo su transportador intracelular, de modo que ha3. La extrusión del calcio hacia el torrente sance llegar el calcio a la membrana basolateral desde guíneo en la cara basolateral. donde mediante transporte activo sale fuera de la El calcitriol es el principal factor que controla la célula. Así, la calbindina se encarga de mantener los absorción intestinal de calcio, actuando sobre las niveles de calcio intracelular bajos, contribuyendo tres fases. Otros factores como la hormona paraa incrementar la captación de calcio por los canatiroidea (PTH), glucocorticoides, estrógenos, facles del borde en cepillo del enterocito. Existen dos tores relacionados con el embarazo, hormona de subclases de calbindina denominadas en función crecimiento o factor de crecimiento similar a la inde su peso molecular: calbindina-D9k (9.000 Da) sulina (Insulin-like Growth Factor, IGF) pueden tamy calbindina-D28k(28.000 Da). La calbindina-D9k bién influir en el transporte de calcio. No obstanse expresa en el intestino de mamíferos y en el rite, actualmente no está claro hasta qué punto su ñón de ratón, mientras que la calbindina-D28k se efecto es directo o está mediado por la vitamina expresa en el intestino de aves y en el riñón y el D. La Figura 9 muestra la regulación del transpáncreas de aves y mamíferos, así como en el ceporte transcelular de calcio en el intestino por la rebro de mamíferos. El calcitriol induce la producvitamina D. ción de mRNA de calbindina mediante su unión al 811 Capítulo 1.24. Vitamina D Figura 10. Acciones de la vitamina D a nivel óseo. VDRnuc1,25. La relación entre el calcitriol y la producción de calbindina es tal que la medida de calbindina es una medida directa de la deficiencia o suficiencia de vitamina D. La regulación de la absorción de calcio por la vitamina D se produce en función del calcio ingerido de la siguiente manera. Cuando la ingesta de calcio es alta, la concentración luminal de calcio lo es también, por lo que la necesidad de evitar una entrada excesiva de calcio, que podría, entre otras cosas, ser tóxica para el enterocito, se convierte en esencial. Se produce entonces una disminución del calcitriol circulante (hay que tener en cuenta que la cantidad calcitriol circulante varía de forma inversamente proporcional a la concentración plasmática de calcio), lo que llevará al cierre de los canales de calcio. Además, al disminuir los niveles plasmáticos de calcitriol se produce menos calbindina y como resultado disminuye el transporte de calcio transcelular. La extrusión de calcio a través de la membrana basolateral del epitelio intestinal es un proceso activo (en contra del gradiente electroquímico) mediado por dos transportadores de calcio: la bomba de calcio de membrana plasmática (Plasma Membrane CAlcium pump, PMCA), que es una ATPasa, y un intercambiador Na+/Ca2+ denominado NCX. Este último es responsable del 20% del calcio expulsado a través de la membrana basolateral, y se ha demostrado que el calcitriol no afecta a su acti- 812 vidad. Por el contrario, el calcitriol aumenta los niveles de RNA mensajero y la síntesis proteica de la PMCA. Así, se ha demostrado que los pollos adaptados a una dieta baja en calcio y fósforo presentan un incremento de la síntesis de proteína y de RNA mensajero de la PMCA en comparación con el grupo control, alimentado normalmente. 10.1.2. Hueso (Figura 10) En general, la incorporación de calcio al hueso es un fenómeno que depende de la concentración de calcio circulante, la cual, a su vez, depende fundamentalmente de la absorción intestinal. Consecuentemente, se ha descrito que el principal efecto del calcitriol sobre el metabolismo óseo del calcio se produce a nivel de su absorción intestinal, suministrando suficiente calcio disponible para ser incorporado al hueso. No obstante, el calcitriol tiene efectos directos sobre el hueso que afectan tanto a la formación como a la resorción ósea. El sistema esquelético está constituido por elementos celulares y por la matriz extracelular. Dentro de los elementos celulares es interesante destacar la presencia de células osteoprogenitoras indiferenciadas, con una gran capacidad proliferativa que van perdiendo conforme maduran y se diferencian, dando lugar a varias líneas celulares (ver O. Martínez Augustin | V. Puerta Fernández | M.ª D. Suárez Ortega Capítulo 1.27). De estas líneas interesa resaltar la línea condroprogenitora, cuya célula final es el condroblasto, y la línea osteoprogenitora que origina el osteoblasto maduro, del que, a su vez, deriva el osteocito. Los osteoblastos son células cuya función principal es la secreción de una matriz extracelular en la que se depositarán los iones minerales, regulando así el proceso de calcificación. Por su parte, los osteocitos son las células más abundantes en el hueso maduro y crean una superficie de intercambio entre el hueso y el líquido extracelular a través del cual puede moverse el calcio y el fosfato desde el gran reservorio óseo para mantener la homeostasis plasmática. El calcitriol estimula directamente, mediante su unión al receptor VDRnuc1,25, la diferenciación de osteoblastos y la producción de proteínas de unión a calcio óseo, como la osteocalcina y la osteopontina. De hecho, existen nuevos análogos del calcitriol que tienen acción selectiva anabólica sobre los osteoblastos y, como resultado, producen un incremento en la formación de hueso. Por otra parte, tanto el calcitriol como el 24-Rcalcitriol podrían estimular la formación de hueso promoviendo la diferenciación de condrocitos. En el proceso de formación del hueso en el feto se produce inicialmente un esqueleto formado por cartílago. A continuación, este cartílago se calcifica, produciéndose la invasión vascular del cartílago calcificado y la formación de hueso utilizando como molde el cartílago. Como última etapa, el cartílago es reemplazado por la médula ósea. Con el fin de permitir el crecimiento del hueso en la etapa postnatal, es necesario mantener cartílago. Así, se conservan unas zonas denominadas placas de crecimiento en las que se mantienen condrocitos capaces de producir cartílago, de proliferar, diferenciarse y finalmente sufrir el proceso de calcificación para formar el hueso. Este proceso produce como resultado el crecimiento longitudinal del hueso. No obstante, los condrocitos permanecen en una zona denominada zona de descanso, donde sólo proliferan y se diferencian cuando reciben las señales adecuadas. En el proceso de vascularización de las placas de crecimiento y la formación posterior de hueso intervienen el calcitriol y el 24-R-calcitriol. Ambos son necesarios para el crecimiento óptimo y la diferenciación de las placas de crecimiento. De he- cho, cuando aparece raquitismo (causado por deficiencia de vitamina D), las placas de crecimiento no se mineralizan y las zonas hipertróficas crecen. Las zonas hipertróficas son aquellas que se van formando al crecer el hueso, en las que hay condrocitos maduros no proliferativos, productores de matriz, pero en las que aún no se ha producido la mineralización de la matriz y por tanto no constituyen hueso propiamente dicho. Las zonas hipertróficas crecen porque las placas de crecimiento no se calcifican y el cartílago no se elimina por los condroclastos. Por tanto, no tiene lugar la invasión vascular ni la formación subsiguiente de hueso y se acaba produciendo el arqueamiento característico de las piernas asociado al raquitismo en niños. Además de los tipos celulares mencionados anteriormente, en el hueso se encuentran unas células denominadas osteoclastos, que derivan de las células progenitoras de los granulocitos y los macrófagos. Estas células se transforman en precursores de osteoclastos que son distribuidos al hueso por vía sanguínea. En el hueso, los osteoclastos se encuentran en cavidades denominadas “lagunas de Howship”, donde ejercen su acción erosiva que forma parte del proceso de remodelado óseo: destrucción de la matriz ósea en varios puntos y sustitución por hueso nuevamente formado. Del equilibrio de estos procesos que se producen durante toda la vida depende la masa ósea del individuo. El osteoclasto no permanece activo de forma continua, sino que se activa cuando existe una demanda de calcio (en este caso, el esqueleto actúa como reserva de calcio) y retorna a la inactividad una vez satisfecha. Dado que el osteoclasto no presenta receptores para la PTH, es necesario que esta hormona actúe indirectamente a través del calcitriol. Así, cuando se produce una disminución de los niveles plasmáticos de calcio, se incrementan los niveles de PTH y como consecuencia se incrementa la síntesis de calcitriol. La actuación de este metabolito de la vitamina D sobre los osteoblastos hace que se produzcan citokinas y factores de crecimiento que estimulan la actividad y la formación de los osteoclastos. Además, el calcitriol actúa directamente incrementando la formación de nuevos osteoclastos y su diferenciación. En este efecto del calcitriol está implicado el VDRnuc1,25. En definitiva, el calcitriol incrementa la actividad y el número de osteoclastos, produciendo la resorción del hueso y la liberación del calcio óseo. 813 Capítulo 1.24. Vitamina D 10.1.3. Riñón En el riñón el calcio es inicialmente filtrado de forma masiva (hasta 10 g/día) en el túbulo proximal e igualmente absorbido a continuación en el túbulo contorneado proximal. No obstante, a nivel renal el sitio clave para la regulación hormonal del calcio es el túbulo distal. Es, concretamente, en esta parte donde se produce la reabsorción selectiva de calcio, controlándose así las pérdidas urinarias netas. Este proceso está regulado a nivel molecular por mecanismos similares a los implicados en el transporte activo de calcio en el intestino, aunque en el intestino la absorción de calcio está más regulada en las regiones proximales (duodeno y yeyuno) que en las distales, donde la absorción de calcio es menos saturable. El calcitriol afecta al transporte de calcio a través de la membrana celular, ejeciendo su efecto sobre la entrada a través de la membrana apical (incrementa los niveles de RNA mensajero del transportador EcaC1), sobre la difusión a través del citosol mediante la calbindina (incrementa los niveles de esta proteína por el mismo mecanismo descrito en intestino), y sobre la extrusión activa de calcio a través de la membrana basolateral. 10.2. Acciones no clásicas de la vitamina D Además de las acciones clásicas de la vitamina D sobre la absorción y el uso del calcio y el fosfato, se ha descrito que esta vitamina es capaz de afectar a la proliferación y a la diferenciación celular y que tiene efectos sobre la respuesta inmune y del sistema nervioso (Figura 8). Estas acciones no clásicas de la vitamina D han hecho que se asocien parámetros como la ingesta de vitamina D, las concentraciones plasmáticas de 25(OH) vitamina D3 o variaciones alélicas en el gen del VDR con la incidencia de múltiples enfermedades como ciertos tipos de cáncer, infecciones, enfermedades autoinmunes, enfermedades inflamatorias, la diabetes tipo I, la hipertensión o enfermedades cardiovasculares. En consecuencia, la obtención de análogos sintéticos de las moléculas activas de la vitamina D se ha convertido en los últimos años en un campo de investigación muy activo. A continuación, se discutirán las acciones no clásicas de la vitamina D y su papel en el tratamiento de ciertas enfermedades. 814 10.2.1. Efectos de la vitamina D sobre la proliferación celular, la diferenciación celular y la apoptosis La vitamina D es antiproliferativa, promueve la maduración celular, e induce tanto la diferenciación como la apoptosis en diferentes líneas celulares, incluyendo líneas cancerosas. El papel de la vitamina D en estas células puede ejercerse mediante su unión al VDRnuc1,25. De hecho, se ha demostrado la presencia de VDRnuc1,25 en células de la glándula mamaria, el colon, la próstata y el sistema nervioso central. Por otra parte, se ha descrito que estas células también expresan 25(OH) vitamina D3 1α-hidroxilasa, lo que podría indicar que pueden producir localmente calcitriol con el fin de regular su diferenciación, proliferación y muerte celular programada. Por tanto, en los últimos años se ha hecho patente que la vitamina D juega un papel importante en el crecimiento y en la diferenciación celular, y que podría proteger frente al inicio y la progresión del cáncer. 10.2.1.1. Cáncer Hay evidencias de que la exposición a la luz solar se asocia con una baja tasa de mortalidad por cáncer de mama, colon y próstata, mientras que la distribución geográfica del raquitismo coincide con la de muertes por cáncer. Además, y más directamente, en varios estudios se han correlacionado bajas ingestas de vitamina D y niveles séricos bajos de 25(OH) vitamina D3 con un incremento en el riesgo de padecer cáncer de mama, de próstata, de colon y colorrectal. La agresividad del cáncer puede también estar relacionada con los niveles séricos de vitamina D; de hecho, las concentraciones séricas bajas de vitamina D se han relacionado con una mayor agresividad del cáncer de próstata y de mama. A nivel molecular, varios estudios genéticos han identificado alelos específicos del VDR que se correlacionan con un incremento del riesgo de cáncer de mama esporádico y con la agresividad de las metástasis. Estudios similares han relacionado alelos específicos con el cáncer de próstata y colon. Los efectos específicos del calcitriol sobre las células de cáncer de mama, leucemia mieloide y tumores del sistema nervioso central han sido estudiados principalmente en modelos animales y celulares. Entre ellos, se incluyen la inhibición del crecimiento O. Martínez Augustin | V. Puerta Fernández | M.ª D. Suárez Ortega Figura 11. Acciones de la vitamina D sobre la inmunidad. mediante la modulación de la maquinaria del ciclo celular, y la consiguiente detención de las células en la fase Go/G1, y la disrupción de la función mitocondrial con el fin de inducir muerte celular por apoptosis. Además, se ha demostrado que el calcitriol puede suprimir la tumorogénesis mamaria. En cuanto a los estudios clínicos, se ha descrito la efectividad del calcitriol en el tratamiento de glioblastomas en un estudio clínico de fase II. Otro estudio ha demostrado que la proliferación de las células del colon de pacientes con riesgo de neoplasia cólica puede ser reducida con la administración de grandes dosis de 25(OH) vitamina D3. 10.2.2. Efectos de la vitamina D sobre el sistema inmune 10.2.2.1. Infecciones Los macrófagos representan la primera línea de defensa inespecífica del sistema inmune. El calcitriol es capaz de inducir la diferenciación de monocitos a macrófagos y de incrementar la tasa de fagocitosis y actividad de estos últimos mediante la inducción de la producción de enzimas lisosómi- cas. Este efecto está mediado por un incremento en la expresión de receptores de superficie-Fc específicos y por un incremento de la respiración celular. Además, es interesante reseñar que los macrófagos poseen actividad 1α-hidroxilasa y pueden por tanto sintetizar calcitriol a partir de 25(OH) vitamina D3. La actividad de esta enzima está incrementada en macrófagos activados, lo que produce un incremento en la concentración de calcitriol en estas células (Figura 11). Existen datos epidemiológicos que señalan una correlación entre la deficiencia de vitamina D y un mayor riesgo de infección. Estos estudios indican que la incidencia y la prevalencia de enfermedades respiratorias en niños con raquitismo nutricional son mayores. 10.2.2.2. Inflamación y enfermedades autoinmunes En los últimos años se ha demostrado que el calcitriol tiene efectos moduladores sobre el sistema inmune específico, los cuales están siendo estudiados activamente. De hecho, tanto el calcitriol como varios análogos estructurales del mismo han demostrado tener efectos beneficiosos en el tratamiento de enfermedades autoinmunes producidas en modelos animales, como la diabetes, la artritis o la nefritis, y en modelos de transplantes. Las citokinas derivadas de los macrófagos producen la diferenciación de los linfocitos T-colaboradores en reposo (Th) hasta células Th0. Posteriormente, y gracias a la influencia de factores adicionales como citokinas exógenas y moléculas coestimuladores producidas por células presentadoras de antígenos (macrófagos y células dendríticas), las células Th0 se diferencian de las células Th1 o Th2. Ambos tipos de células secretan un perfil específico de citokinas que están implicadas en la proliferación y en la diferenciación de células T y B. El calcitriol puede regular la respuesta inmune tanto en 815 Capítulo 1.24. Vitamina D órganos linfoides secundarios como en tejidos diana mediante varios mecanismos (Figura 11): a) El calcitriol inhibe la diferenciación y la maduración de células dendríticas, que son células presentadoras de antígenos, cruciales en la inducción de la respuesta inmune mediada por células T. b) El calcitriol inhibe el desarrollo de células Th1 en tanto que induce el desarrollo de células CD4+CD25+ y de células Th2. Estos dos últimos tipos célulares son capaces de inhibir a las células Th1. El calcitriol inhibe la producción de IL-12 y estimula la producción de IL-10, a la vez que disminuye la expresión de las moléculas coestimuladoras (CD40, CD80, CD86) en células dendríticas (presentadoras de antígenos). Como consecuencia se inhibe el desarrollo de células Th1. c) El calcitriol actúa inhibiendo la síntesis del mRNA de citokinas producidas por macrófagos y células presentadoras de antígenos, como son la interleukina (IL)-1, la IL-6, la IL-12 y el factor de necrosis tumoral (TNF-α). Además, el calcitriol estimula la secreción de prostaglandina E2 (PGE2) de carácter antiinflamatorio, mientras que inhibe la producción del factor estimulador de colonias de granulocitos-macrófagos (GM-CSF), responsable de la producción de nuevos monocitos. d) El calcitriol puede disminuir la actividad presentadora de antígenos de los macrófagos a los linfocitos mediante la disminución de la expresión en la superficie celular de moléculas del complejo principal de histocompatibilidad de clase II (MHC-II). e) El calcitriol actúa directamente sobre las células T, inhibiendo la secreción de IL-2 (esencial para la expansión clonal de los linfocitos) e interferón-γ (IFN-γ) por células Th1. 10.2.2.3. Artritis reumatoide La artritis reumatoide se caracteriza por la infiltración de macrófagos, linfocitos T y células plasmáticas en el sinovio, produciendo un estado de inflamación crónica, caracterizado por la producción de citokinas como la IL-6 y el TNF-α. Entre otros signos, los pacientes que sufren artritis reumatoide poseen altos niveles de proteína C reactiva, un marcador bioquímico de inflamación. Varios estudios han indicado que existe una correlación entre el padecimiento de artritis reumatoide y la gravedad de la enfermedad y los nive- 816 les bajos de vitamina D. Además, estudios clínicos han indicado que la administración de 2 µg/día de vitamina D a pacientes afectados de artritis reumatoide es capaz de aliviar el dolor y de producir una disminución significativa de los niveles séricos de proteína C reactiva. 10.2.2.4. Enfermedad inflamatoria intestinal La enfermedad inflamatoria intestinal es otro tipo de enfermedad inflamatoria que ha sido relacionada con niveles bajos de vitamina D. De hecho se ha observado que los pacientes afectados por esta enfermedad poseen niveles más bajos de vitamina D y que la cantidad de vitamina D disponible puede ser un factor importante en el desarrollo de la enfermedad. En estudios recientes, se ha descrito que el receptor de la vitamina D podría tener un papel crucial en la regulación de la inflamación en el tracto gastrointestinal y concretamente en la enfermedad inflamatoria intestinal. En estos estudios se ha demostrado que la eliminación del gen del VDRnuc1,25 en diversos modelos de enfermedad intestinal en ratones produce la aceleración del desarrollo de la enfermedad inflamatoria intestinal y además un incremento en su gravedad y en la mortalidad. 10.2.2.5. Esclerosis múltiple La esclerosis múltiple es una enfermedad en la que se produce una desmielinización del sistema nervioso central que parece estar causada por procesos autoinmunes mediados por los linfocitos T. La enfermedad se manifiesta generalmente entre los 20 y los 40 años. Varios hechos indican que un estado inadecuado de vitamina D es un factor patogénico importante en el desarrollo de esta enfermedad. Así, la prevalencia de esta enfermedad es prácticamente nula en zonas cercanas al ecuador y manifiesta un gradiente de prevalencia norte-sur. Por otra parte, varios estudios indican que en gran parte de los pacientes afectados los niveles de vitamina D son insuficientes. Un estudio con enfermos de esclerosis múltiple ha demostrado que la suplementación con calcio, magnesio y vitamina D (125 µg/día) durante 1-2 años fue capaz de reducir la incidencia de recaídas con respecto a las cifras esperadas. O. Martínez Augustin | V. Puerta Fernández | M.ª D. Suárez Ortega Varios estudios han demostrado que los efectos beneficiosos de la vitamina D en la esclerosis múltiple pueden ser debidos a la inhibición de las células Th1 (inflamatorias), a la inhibición de la producción de citokinas inflamatorias por macrófagos activados, a un incremento de la producción de citokinas antiinflamatorias y a la acción antiproliferativa en linfocitos mediante la expresión del VDRnuc1,25. En línea con estas observaciones se ha descrito recientemente que la suplementación con vitamina D es capaz de reducir los niveles de mRNA de IL-2 en células mononucleares de sangre periférica de pacientes con esclerosis múltiple. Otro mecanismo posible es la inhibición de la expresión de iNOS en el sistema nervioso central. 10.2.3. Efectos de la vitamina D sobre el sistema renina-angiotensina El sistema renina-angiotensina juega un papel esencial en la regulación de la presión sanguínea. La renina se produce y secreta predominantemente en el aparato yuxtaglomerular. Su principal función es la de cortar el angiotensinógeno para obtener un decapéptido denominado angiotensina I, el cual se transforma posteriormente en un octapéptido denominado angiotensina II por la acción de la enzima convertidora de angiotensina. La angiotensina II es, mediante su acción sobre diversos órganos, el efector principal del sistema renina angiotensina, mientras que la regulación principal se produce a nivel de la síntesis y secreción y por tanto de la actividad de la renina. Varios estudios clínicos y epidemiológicos han sugerido una relación inversa entre la vitamina D, la presión sanguínea y la actividad de la renina plasmática. Efectivamente, se ha demostrado tanto en pacientes normotensos como hipertensos que los niveles séricos de vitamina D están inversamente asociados con la presión sanguínea y la actividad de la renina plasmática. Además, en estudios clínicos se ha descrito que la vitamina D reduce la presión sanguínea en ancianos hipertensos. De hecho, se ha demostrado que el tratamiento calcitriol reduce la actividad de la renina plasmática, los niveles de angiotensina II, la presión sanguínea y la hipertrofia del miocardio. Aunque no se conoce el mecanismo de acción del calcitriol sobre el sistema renina-angiotensina, sí hay datos que indican que su efecto está mediado por el VDRnuc1,25 y recientemente se ha descrito que el calcitriol es un potente inhibidor de la expresión génica de la renina. 10.2.4. Efectos de la vitamina D sobre el sistema nervioso El calcitriol puede sintetizarse y degradarse en el cerebro, ya que se ha demostrado la existencia de vitamina D3 25-hidroxilasa, 25-dihidroxivitamina D3 1α-hidroxilasa y vitamina D3 24-hidroxilasa en el cerebro. Además, tanto en el cerebro como en la médula espinal existe VDRmem1,25 por lo que tanto las células de la glia, como las neuronas y los astrocitos podrían ser una diana importante para el calcitriol. La vitamina D podría tener efectos neuroprotectores en el sistema nervioso. Los mecanismos descritos que avalan este efecto son los siguientes: a) Se ha demostrado que el calcitriol induce la muerte y/o rediferenciación de células del glioma. Por tanto, su síntesis por células activadas de la microglia y por neuronas, las cuales expresan 25-dihidroxivitamina D3 1α-hidroxilasa podría constituir una respuesta antitumoral del sistema nervioso central. Esta respuesta podría estar regulada por astrocitos mediante la expresión de vitamina D3 24-hidroxilasa. b) Se ha demostrado que en astrocitos el calcitriol induce la síntesis de varias neutrofinas [γ-glutamil transferasa, neutrofina 3 (NF3), factor neutrófico derivado de la línea celular glial (GDNF) y factor de crecimiento de nervios (NGF)], que podrían ejercer efectos neuroprotectores. c) El calcitriol inhibe la síntesis de iNOS. Los niveles elevados de óxido nítrico son tóxicos tanto para las neuronas como para los oligodendrocitos. Además, el óxido nítrico puede reaccionar con otras moléculas, produciendo radicales libres que resultan nocivos. Por otra parte, la γ-glutamil transferasa, cuya síntesis por los astrocitos es inducida por el calcitriol, podría prevenir la formación de radicales reactivos del nitrógenos y del oxígeno. El VDRmem1,25 presente en el sistema nervioso, y por tanto la vitamina D, podría tener un papel importante en la regulación de la neuro-ontogénesis. Los hechos que avalan esta hipótesis son los siguientes: 817 Capítulo 1.24. Vitamina D a) La expresión de VDRnuc1,25 está regulada por el desarrollo en los tejidos del sistema nervioso. b) El VDR se ha localizado en el neuroepitelio durante la neurogénesis y, más tarde, en la zona subventricular del cerebro en áreas que son capaces de mantener la generación de células madre durante la vida. c) El gen del VDR se expresa específicamente en las neuronas ganglionares de la raíz dorsal de roedores, lo cual indicaría que la vitamina D ejerce una función en el desarrollo del sistema nervioso periférico. Se ha demostrado que la vitamina D puede estar relacionada con diversas enfermedades que afectan al sistema nervioso, como la esclerosis múltiple, la isquemia cerebral e incluso el Alzheimer. 11. Niveles normales y deficiencia de vitamina D Como ya se ha comentado anteriormente, y aunque carece de actividad, la concentración sérica de 25(OH) vitamina D3 es el parámetro utilizado para el estudio de los niveles de vitamina D, siendo la concentración normal de este metabolito en suero de 25-50 ng/ml. La revisión de estudios publicados entre 1990 y 1999 indica que en España entre un 47,1% y un 94,2% de los individuos tienen ingestas de vitamina D inferiores a las recomendadas. No obstante, el riesgo de carencia se resuelve por la posibilidad de sintetizar la vitamina en el organismo, aunque el aporte es claramente deficitario en personas con escasa exposición a la luz solar y en embarazadas. La ingesta media de vitamina D en embarazadas en España es de 3,1 ± 1,2 µg/día, muy inferior a la ingesta de referencia. De hecho, más del 90% de las embarazadas estudiadas presentan aportes de vitamina D inferiores a los recomendados. Concentraciones séricas de 3 ng/ml de 25(OH) vitamina D3 se asocian con signos clínicos de deficiencia. No obstante, se interpreta que existe déficit de vitamina D cuando la concentración de 25(OH) vitamina D3 en suero es menor o igual a 12 ng/ml. Cuando esta deficiencia es continua durante meses se produce raquitismo u osteomala- 818 cia. Ambos términos designan el mismo trastorno, aunque el término raquitismo es utilizado cuando se produce en niños, mientras que el de osteomalacia se utiliza para adultos. Los cambios bioquímicos característicos que se producen cuando hay deficiencia de vitamina D incluyen niveles plasmáticos bajos de calcio y fósforo inorgánico, mientras que la fosfatasa alcalina se encuentra incrementada en el plasma. Inicialmente la deficiencia produce una disminución de la absorción de calcio y un hiperparatiroidismo secundario, secretándose PTH como respuesta a los bajos niveles de calcio. Con el calcitriol remanente se moviliza el calcio óseo, restaurándose los niveles de calcio séricos a valores normales. No obstante, la PTH también causa fosfaturia e hipofosfatemia, lo que da lugar a fallos en la mineralización ósea y eventualmente a la aparición de los sígnos clínicos óseos característicos del raquitismo y la osteomalacia. En ambos casos, el diagnóstico se realiza mediante la determinación de las concentraciones plasmáticas de calcio, fósforo, fosfatasa alcalina, PTH y 25(OH) vitamina D3, además del diagnóstico radiográfico de las deformaciones óseas. Los signos clínicos óseos característicos del raquitismo incluyen tumefacciones a nivel de las epífisis de los huesos largos e incurvaciones producidas por ablandamiento de los huesos. El crecimiento óseo se produce a través de la creación de nuevo cartílago y de zonas hipertróficas en las placas de crecimiento de los extremos de los huesos. Cuando el fosfato cálcico se deposita en el cartílago se crea una estructura dura. En el caso de que exista deficiencia de vitamina D3, no hay calcio disponible para la mineralización del hueso y como resultado se produce hueso blando. Otros síntomas típicos del raquitismo a nivel óseo incluyen la aparición de protuberancias óseas en las costillas y las rodillas, denominados rosario costral, la aparición de craneomalacia o el aplastamiento anteroposterior del tórax. Éstos pueden aparecer acompañados de hipotonía muscular y retraso motor e incluso de convulsiones que se pueden dar ocasionalmente en niños con raquitismo por los bajos niveles de calcio en sangre. El raquitismo se cura rápidamente con la administración durante 1 mes de 4.000 UI de vitamina D por vía oral. Durante este periodo de tiempo, se deben monitorizar los niveles de 25(OH) vita- O. Martínez Augustin | V. Puerta Fernández | M.ª D. Suárez Ortega mina D3, con el fin de asegurar que los niveles se han normalizado. Además de la administración de vitamina D, se aconseja que estos niños reciban luz solar o radiación de una lámpara de luz ultravioleta durante al menos 20 minutos al día. En el adulto, la deficiencia en vitamina D provoca defectos en la mineralización del hueso que se manifiestan con la aparición de dolores en la zona dorsolumbar, cintura pélvica y huesos. En el control radiológico, se aprecia un aspecto borroso y algodonoso de la sustancia ósea con una transparencia anormalmente aumentada. La osteomalacia se trata con la ingestión diaria de 2.500 UI de vitamina D durante al menos 3 meses. Se aconseja también la exposición diaria al sol o en su defecto el uso de lámparas de luz ultravioleta. Los niveles plasmáticos de 25(OH) vitamina D3 deben ser evaluados al final de este periodo para asegurar la eficacia de la terapia. Dado que la vitamina D es tóxica en dosis superiores o iguales a las 50.000 UI al día en adultos y 1.000 UI al día en niños, en ambos tratamientos se debe tener cuidado por si aparecen síntomas de toxicidad producidos por un exceso de vitamina D. La prognosis de la osteomalacia y el raquitismo es excelente. El tratamiento con vitamina D produce la normalización de la mineralización ósea y la corrección de los niveles plasmáticos de calcio. Además, las anormalidades óseas en niños generalmente desaparecen en un periodo de 3-9 meses, aunque en casos graves pueden persistir de por vida. Como se ha comentado anteriormente, los grupos de riesgo en el padecimiento de raquitismo u osteomalacia son las personas oscuras de piel, aquellas que viven en países en los que los inviernos son largos, las mujeres de países islámicos, los enfermos que tienen impedida su movilidad, los ancianos y los niños. En definitiva, aquellos grupos de población en los que la exposición al sol no es suficiente. Además, la deficiencia de vitamina D puede aparecer en pacientes con alteraciones del funcionamiento renal y hepático o de la absorción intestinal que pueden interferir con los mecanismos de absorción, transporte o metabolismo de la vitamina D; por ejemplo, síndromes de malabsorción y esteatorrea producidos por enfermedades como la enfermedad celiaca, la enfermedad inflamatoria intestinal, una pancreatitis crónica o una insuficiencia hepática. Además, determinadas situaciones quirúrgicas como resecciones gástricas o bypass yeyuno-ileal pueden producir esta deficiencia. Los pacientes que sufren fallo renal crónico presentan frecuentemente baja absorción de calcio, hipocalciemia, hiperparatiroidismo secundario y ostrodistrofia. Estos pacientes sufren frecuentemente osteítis fibrosa y/u osteomalacia. Estos signos se producen como consecuencia de una disminución en la excreción de fósforo que produce la consiguiente inhibición de los niveles de PTH y la disminución de la actividad de la 25(OH) vitamina D3 1α-hidroxilasa renal y por tanto de la síntesis de calcitriol. A estos pacientes se les suele administrar 1α-hidroxivitamina D3 o 1α-hidroxivitamina D2. Ambas formas de vitamina D pueden ser metabolizadas por la 25-hidroxilasa renal para obtener el calcitriol. Por otro lado, el uso prolongado de fármacos anticomiciales, como se ha explicado anteriormente, altera el metabolismo del calcio estimulando la acción de hidroxilasas dependientes de citocromo P-450, lo que acelera la degradación del calcitriol, acelerando procesos de raquitismo u osteomalacia. 12. Enfermedades relacionadas con alteraciones en el metabolismo de la vitamina D o en la respuesta a vitamina D 12.1. Raquitismo resistente a vitamina D o hipofosfatemia familiar La hipofosfatemia familiar es una enfermedad hereditaria ligada al cromosoma X cuya disfunción principal consiste en la pérdida de fosfato que se produce a nivel del túbulo renal, lo que conlleva una disminución de los niveles séricos de fosfato y el incremento de fosfatasa alcalina plasmática. Además, la absorción intestinal de calcio y fosfato se encuentra disminuida y se observan concentraciones elevadas de PTH en los sujetos afectados. Se ha descrito asimismo que el metabolismo de la vitamina D se encuentra alterado, aunque ésta no es la causa de la hipofosfatemia. 819 Capítulo 1.24. Vitamina D Las causas moleculares de esta enfermedad están siendo objeto de estudio. De hecho, se ha identificado un gen denominado PHEX (Phosphate regulating gene with Homologies to Endopeptidases, on the X chromosome: gen regulador de fosfato con homología con endopeptidasas, en el cromosoma X), que podría ser el responsable de la enfermedad, ya que se han detectado múltiples mutaciones en este gen en pacientes con hipofosfatemia familiar. El gen PHEX codifica una proteína de 749 aminoácidos que posee homología con metalopeptidasas de membrana con afinidad por el zinc. Estas endopeptidasas están en general implicadas en la degradación o activación de una gran variedad de hormonas peptídicas. Se cree que la proteína codificada por el gen PHEX podría estar implicada en la activación de una serie de hormonas que se han denominado fosfatoninas, las cuales estarían relacionadas con la absorción de fosfato, hipótesis esta que ha sido formulada recientemente por algunos investigadores y está adquiriendo cada vez más relevancia. La hipofosfatemia familiar normalmente se manifiesta en la infancia y en la pubertad durante la fase rápida de crecimiento, y su signo más evidente es un retraso del mismo. Aunque la administración individual de vitamina D o de fósforo inorgánico no restaura el crecimiento, la administración conjunta de ambos ha dado buenos resultados, posiblemente porque la administración de fosfato contrarresta su pérdida renal, mientras que la vitamina D en forma de calcitriol o de vitamina D2 incrementa la absorción de calcio y previene el hiperparatiroidismo secundario a la enfermedad. También se ha descrito que la suplementación con 24-R-calcitriol es capaz de disminuir los niveles de hormona paratiroidea y de mejorar los síntomas de raquitismo y osteomalacia en los enfermos afectados. Por otra parte, en algunos estudios el tratamiento con hormona de crecimiento ha demostrado ser efectivo, mejorando el crecimiento lineal y produciendo una disminución transitoria en la excreción de fosfato urinario. 12.2. Hipoparatiroidismo El hipoparatiroidismo aparece generalmente como resultado de la extirpación quirúrgica de las gándulas paratiroides. Como ya se ha comentado 820 anteriormente, la hormona paratiroidea estimula la síntesis de calcitriol, por lo que el hipoparatiroidismo se acompaña de una disminución en la síntesis de esta vitamina. En consecuencia, los pacientes afectados de esta enfermedad producen cantidades inadecuadas de vitamina D en respuesta a la hipocalcemia. La administración de 25(OH) vitamina D3, 1α (OH) vitamina D3, calcitriol, dihidrotaquisterol o vitamina D2 son efectivas en el tratamiento de estos pacientes, siempre que la ingesta de calcio sea adecuada. La 25(OH)vitamina D3 es la molécula más efectiva de las utilizadas. El hipoparatiroidismo cursa con convulsiones y espasmos tetánicos, por lo que los pacientes que padecen esta enfermedad son frecuentemente sometidos a terapia con anticonvulsivantes antes de que se detecte la verdadera patología. Como se ha indicado el uso prolongado de anticonvulsivantes acelera la degradación metabólica de la vitamina D, lo que podría acentuar la sintomatología de la enfermedad. 12.3. Raquitismo tipo I dependiente de vitamina D El raquitismo tipo I dependiente de vitamina D es una enfermedad hereditaria con carácter autosómico recesivo que se caracteriza por la existencia de niveles anormalmente bajos en sangre de calcio y fósforo, además de la aparición de retraso en el crecimiento, anormalidades óseas características del raquitismo y miopatía. Cuando se comparan los signos de esta enfermedad con los de la hipocalcemia familiar, se observa que la aparición de los síntomas es más temprana en el raquitismo dependiente de vitamina D y que responde mejor a la administración de la vitamina. Además, la miopatía es característica de esta enfermedad. La causa del raquitismo tipo I es la deficiencia en la actividad 25(OH) vitamina D3 1α-hidroxilasa, lo que da lugar a la existencia de niveles séricos anormalmente bajos de calcitriol y, entre otros efectos, la consecuente disminución en la absorción intestinal de calcio. Se ha demostrado que existen múltiples mutaciones en el gen que codifica esta enzima responsables de la enfermedad. La administración diaria de calcitriol es suficiente para contrarrestar los síntomas de la enfermedad. O. Martínez Augustin | V. Puerta Fernández | M.ª D. Suárez Ortega 12.4. Raquitismo tipo II dependiente de vitamina D El raquitismo tipo II dependiente de vitamina D es una enfermedad autosómica recesiva poco frecuente. Esta enfermedad se caracteriza por la aparición de raquitismo u osteomalacia con hipocalcemia e hiperparatiroidismo secundario. A diferencia del raquitismo tipo I, esta enfermedad está asociada a valores séricos normales de calcitriol. Es bien conocido que esta enfermedad se debe a mutaciones específicas en el DNA del hVDRnuc1,25, dando lugar a una forma de resistencia periférica a las acciones del calcitriol. Los niveles séricos de 25(OH) vitamina DhVDR3 son normales, mientras que los de calcitriol están elevados y los del 24-R-calcitriol son indetectables. Los signos clínicos y radiológicos más característicos de esta enfermedad coinciden con los del raquitismo, pudiendo estar o no asociada a la aparición de alopecia. De esta forma se distinguen dos tipos de esta enfermedad, el tipo IIA (con alopecia) y el tipo IIB (sin alopecia). El tratamiento clásico de esta enfermedad requiere dosis elevadas de calcio y vitaminas D2 y D3 por vía parenteral. Como se ha indicado anteriormente, se han descrito distintas mutaciones en la secuencia de DNA del hVDRnuc1,25 que son responsables de esta enfermedad y que afectan tanto a la unión del receptor a su ligando, como a la localización nuclear del complejo hormona-receptor, a la unión del receptor a su secuencia diana o a la unión del receptor a un coactivador. Otras proteínas distintas del hVDRnuc1,25 pueden ser también responsables de raquitismo. Así, es interesante resaltar que recientemente se ha descrito la existencia de casos de raquitismo con alopecia en pacientes con VDRnuc1,25 normal. Se ha demostrado que estos pacientes sobreexpresaban una proteína nuclear llamada proteína de unión al elemento de respuesta de la vitamina D (Vitamin D Response Element Binding Protein, VDRE-BP) que compite con el dímero VDR-RXR por la unión a la VDRE, actuando como reguladora. 821 Capítulo 1.24. Vitamina D 13. Resumen El raquitismo y la osteomalacia son producidos por la deficiencia prolongada de vitamina D. Ambos transtornos se conocen desde la Antigüedad, ya que tuvieron una incidencia bastante alta en siglos pasados y aún son prevalentes en ciertos grupos de población. Los ancianos, los niños y aquellas personas que por cualquier causa no reciben suficiente luz solar, son grupos de riesgo de deficiencia de vitamina D. La deficiencia continua de vitamina D produce la aparición de raquitismo y osteomalacia. Ambos términos engloban el mismo trastorno, aunque el término raquitismo es utilizado cuando se produce en niños, mientras que el de osteomalacia se utiliza para adultos. Los signos más característicos del raquitismo afectan al esqueleto y consisten en la aparición de deformaciones óseas. La administración de dosis altas de vitamina D por vía oral durante unos meses es suficiente para curar ambos trastornos. En los últimos años, la vitamina D y el desarrollo de análogos de esta vitamina están cobrando gran importancia, ya que cada vez es mayor el número de procesos fisiológicos en cuya regulación interviene esta vitamina. Además, y como consecuencia de lo anterior, la incidencia de diversas enfermedades como el cáncer, la esclerosis múltiple, la hipertensión o la enfermedad inflamatoria intestinal se ha relacionado con niveles bajos de vitamina D, e incluso se ha demostrado que la administración de esta vitamina puede ser beneficiosa en el tratamiento de dichas enfermedades. Por sus características, actualmente se considera que la vitamina D es una vitamina y una hormona. Así, es un compuesto orgánico que actúa como micronutriente y su ingestión es necesaria para la mayoría de las poblaciones urbanas; de aquí que se considere una vitamina. No obstante, la suplementación con vitamina D es innecesaria en individuos que son capaces de completar sus requerimientos mediante síntesis endógena de vitamina D y de metabolitos activos de ésta (el calcitriol y el 24-R-calcitriol). Estos metabolitos actúan sobre distintos órganos diana, por lo que pueden ser considerados hormonas, y la vitamina D una prohormona. 822 La síntesis endógena de vitamina D incluye la activación por irradiación (luz solar) del 7-deshidrocolesterol, un metabolito del colesterol que se produce en el hígado y es exportado a la piel. La vitamina D producida en la piel es a continuación metabolizada sucesivamente en el hígado (por la 25-hidroxilasa) y el riñón (por la 1α-hidroxilasa y por la 24R-hidroxilasa), produciéndose en condiciones normales las formas activas que actúan sobre distintos órganos diana. La enzima clave en la regulación de la vitamina D es la 1α-hidroxilasa renal. Esta enzima se regula en función del calcio circulante, interviniendo en la regulación la hormona paratiroidea (PTH) y el propio metabolito de la enzima. Así, una disminución de la concentración sérica de calcio estimula la liberación de PTH, que a su vez estimula la síntesis del calcitriol, mientras que el calcitriol produce mediante feed-back negativo la inhibición de la enzima y la estimulación de la 24R-hidroxilasa, lo que a su vez favorece la formación de 24,1α,25(OH)2 vitamina D3, que es el principal metabolito inactivo del calcitriol. Los órganos diana de la vitamina D están definidos por la presencia de tres receptores distintos a través de los cuales la vitamina D ejerce su acción. Así, el metabolito principal de la vitamina D (calcitriol) se puede unir al receptor nuclear de vitamina D (Vitamin D Receptor, VDRnuc1,25) y al receptor de membrana de vitamina D (VDRmem1,25). Por su parte, el 24-R-calcitriol se une a otro receptor de membrana denominado VDRmem24,25. En general, la función del receptor nuclear está relacionada con la respuesta genómica a nivel transcripcional, mientras que los receptores de membrana median las llamadas respuestas biológicas rápidas (no transcripcionales) de la vitamina D, que implican la estimulación de cascadas de transducción de señal. El receptor nuclear de la vitamina D ha sido objeto de intensos estudios en los últimos años. Su importancia radica en que se encuentra ampliamente distribuido en distintos tipos de células y además media las acciones de la vitamina D desconocidas hasta hace unos años, sobre órganos diana también desconocidos hasta hace poco tiempo. Además, mutaciones en este receptor son responsables de la aparición de raquitismo tipo II dependiente de vitamina D. O. Martínez Augustin | V. Puerta Fernández | M.ªD. Suárez Ortega La vitamina D3 participa de manera activa en la regulación de la homeostasis mineral, concretamente en el mantenimiento de la concentración circulante de calcio. Así, los metabolitos activos de la vitamina D incrementan la absorción intestinal de calcio a la vez que disminuyen su excreción renal. Además, actúan sobre el hueso estimulando la movilización del calcio óseo (resorción ósea). Recientemente, se ha descrito que la vitamina D actúa sobre múltiples dianas, regulando procesos como la diferenciación celular, la proliferación celular o la apoptosis. Se ha descrito que esta vitamina induce la diferenciación celular y la apoptosis, mientras que es capaz de inhibir la proliferación celular. Por otra parte, la vitamina D es capaz de actúar sobre el sistema inmune, el sistema nervioso o el sistema renina-angiotensina. De hecho, como se ha comentado anteriormente, se ha relacionado la presencia de niveles bajos de vitamina D con la mayor incidencia de enfermedades como el cáncer, la enfermedad inflamatoria intestinal, la esclerosis múltiple, la hipertensión o la artritis reumatoide. 823 Capítulo 1.24. Vitamina D 14. Bibliografía Revisión en la que se recoge en detalle y de manera comprensible la información disponible sobre el receptor nuclear de la vitamina D. Li YC. Vitamin D regulation of the renin-angiotensin system. J Cell Biochem 2003; 88: 327-31. Artículo que revisa las acciones de la vitamina D sobre el sistema renina-angiotensina y sus implicaciones en la regulación de la presión arterial. Aranceta J, Serra Majem L, Ortega R, Entrala A, Gil A. Libro blanco. Las vitaminas en la alimentación de los españoles. Estudio eVe. Editorial Médica Panamericana. Madrid, 2000. ISBN: 84-7903-578-1. Información detallada sobre la ingesta de vitaminas en la población española según grupos de edad y sexo. Además, se definen los grupos de riesgo de deficiencia en vitaminas de la población española y se recogen los criterios de suplementación de vitaminas en España. Bender DA. Vitamin D. En: Bender DA (eds.). Nutritional Biochemistry of the Vitamins, 2nd ed. Cambridge University Press. Cambridge, UK, 2003; Chapter 3: 77-108. Capítulo que recoge información actualizada sobre la vitamina D. Bronner F. Mechanisms of intestinal calcium absorption. J Cell Biochem 2003; 88: 387-93. Artículo que revisa los conocimientos en los mecanismos que regulan la absorción intestinal de calcio. Garcion E, Barbot-Wion N, Monetero-Menei CN, Berger F, Wion D. New clues about vitamin D functions in the nervous system. Trends in Molec Med 2002; 13: 100-5. Artículo que revisa los efectos neuroprotectores e inmunomoduladores de la vitamina D y el papel potencial que los análogos de esta vitamina podrían tener en el tratamiento de enfermedades neurodegenerativas y neuroinmunes. Jurutka PW, Whitfield GK, Jsieh JC, Thompson PD, Haussler CA, Haussler MR. Molecular nature of the vitamin D receptor and its role in regulation of gene expression. Rev Endocr Metab Disord 2001; 2: 203-16. 15. Enlaces web www.beyonddiscovery.org www.cc.nhi.gov/ccc/supplements/vitd.html#rda www.ncbi.nlm.nih.gov/omim/ medlineplus.gov/spanish/ vdr.bu.edu/ 824 Mathieu C, Adorinin L. The coming age of 1,25-hidroxyvitamin D3 analogs as immunomodulatory agents.Trends in Molec Med 2002; 8: 174-9. Revisión sobre los últimos avances en el estudio del mecanismo de acción de la vitamina D sobre el sistema inmune y el posible papel de la vitamina D y de sus análogos en el tratamiento de enfermedades autoinmunes y de los rechazos a injertos. McLaren DS, Loveridge N, Duthie G, Bolton-Smith C. Fatsoluble vitamins. En: Garrow JS, James WPT (eds.). Human Nutrition and Dietetics, 9th ed. Churchil Livingston. Edimburgo, UK, 1993; Chapter 13: 208-38. Capítulo en el que se puede obtener información clara y concisa sobre el mecanismo de acción de la vitamina D. Norman AW, Okamura WH, Bishor JE, Henry HL. Update on biological actions of 1 alpha, 25(OH)2 vitamin D3 (rapid effects) and 24R,25(OH)2 vitamin D3. Mol Cell Endocrinol 2002; 197: 1-13. Revisión en la que se recoge la información existente sobre los receptores de membrana de las distintas formas activas de la vitamina D y sobre las acciones mediadas por estos receptores. Zittermann A. Vitamin D in preventive medicine: are we ignoring the evidence? Br J Nutr 2003; 89: 552-72. Artículo que recoge los argumentos a favor de la utilización de la vitamina D en la prevención de ciertas enfermedades: (1) datos epidemiológicos que han demostrado una relación entre niveles bajos de vitamina D y enfermedades como el cáncer o la esclerosis múltiple y (2) estudios clínicos que han demostrado efectos beneficiosos de la vitamina D en la hipertensión, la diabetes, la artritis reumatoide o la esclerosis múltiple.