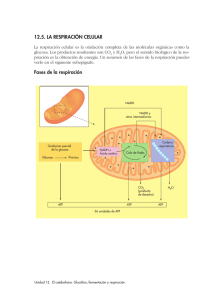
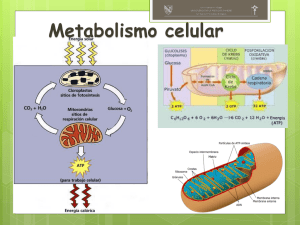
FACULTAD DE FARMACIA Y BIOQUÍMICA E. P. FARMACIA Y BIOQUÍMICA BIOLOGÍA CELULAR METABOLISMO CELULAR MOVILIDAD INTRACELULAR VÍAS SECRETORAS Y ENDOCÍTICAS MG. JULIO REYNALDO RUIZ QUIROZ [email protected] LIMA 2020 CONTENIDO METABOLISMO CELULAR MOVILIDAD INTRACELULAR VÍAS SECRETORAS Y ENDOCÍTICAS Metabolismo celular ANABOLISMO Y CATABOLISMO ➢ Las reacciones metabólicas de la célula la obligan a estar en constante actividad. ➢ Las concentraciones de innumerables sustancias cambian continuamente en el interior de la celular, y otras más entran o salen de ésta. ➢ Durante esa gran actividad, las reacciones químicas tienen una secuencia programada diferente para cada función, según el tipo de célula. A esta secuencia se le conoce como RUTA o VÍA METABÓLICA; en algunos casos, las distintas vías metabólicas pueden interrelacionarse. Algunas hazañas celulares: ✓ En 1 s, una célula normal de nuestro cuerpo emplea un promedio de 10 millones de moléculas de ATP y sintetiza otras tantas. ✓ No es raro que una enzima celular catalice 25000 reacciones o más por segundo; la mayoría de las células contiene cientos o miles de enzimas diferentes. El METABOLISMO CELULAR es la suma de todas las reacciones químicas que ocurren en una célula, cuyas vías o rutas metabólicas pueden dividirse en anabólicas y catabólicas Metabolismo celular ANABOLISMO Y CATABOLISMO El ANABOLISMO, conocido también como BIOSÍNTESIS, es la parte del metabolismo en la que se incorporan sustancias simples a moléculas más complejas necesarias para la célula. El anabolismo se encarga sobre todo de obtener materiales y energía química de las moléculas del alimento, y de canalizarlas para que se realicen las funciones de reparación y crecimiento de la célula. También se encarga de la construcción de nuevas moléculas más complejas que integrarán las reacciones de recambio y reutilización de algunos bloques de moléculas. La energía liberada de los alimentos almacena en forma de compuestos fosfatados de gran energía como el ATP, que posteriormente dirigirá la actividad celular mediante distintas secuencias de reacciones químicas implicadas en la transformación de una sustancia en otra. Estas secuencias, conocidas como rutas o vías metabólicas, serán diferentes de acuerdo con el tipo de célula y la función específica de que se trate (un ejemplo es la fotosíntesis) Un ejemplo de anabolismo es la enzima de ADN ligasa (mostrada en color) que rodea a la doble hélice para reparar una cadena de ADN. Metabolismo celular ANABOLISMO Y CATABOLISMO El CATABOLISMO está representado por las funciones del metabolismo (o vías de metabólicas) de degradación que se acompañan de liberación de energía química, producto, sobre todo, de las moléculas complejas que se descomponen en otras más simples. Una parte de esta energía química se almacena en forma de compuestos de gran energía como el ATP, mientras que otros compuestos más sencillos, como la glucosa, sirven como sustratos para otras reacciones exotérmicas. División de las vías o rutas metabólicas Metabolismo celular ANABOLISMO Y CATABOLISMO De acuerdo con la forma en que la célula obtiene sus materiales y energía para llevar a cabo su metabolismo, la nutrición celular es de dos tipos: AUTÓTROFA y HETERÓTROFA. En la primera, las células obtienen su energía a partir de productos inorgánicos (como la luz solar y elementos químicos), y en la segunda la energía se obtiene de alimentos. Las células capaces de fabricar, mediante reacciones anabólicas, sus propios alimentos y otras sustancias, a partir de productos inorgánicos, se denominas AUTÓTROFAS. La palabra proviene de las raíces griegas, auto, por sí mismo y trophos, que se alimenta o nutre. Se conocen dos procesos de nutrición celular autótrofa de metabolismo anabólico: la quimiosíntesis y la fotosíntesis. ✓ LA QUIMIOSÍNTESIS es característica de organismos procariontes quimiosintéticos o quimioautótrofos. Consiste en obtener energía a partir de compuestos inorgánicos reducidos. Utilizan además el CO2 como fuente principal de carbono. ✓ LA FOTOSÍNTESIS es un proceso que se lleva a cabo en presencia de luz y es característica de células procariontes y eucariontes que poseen clorofila. A estas células se les conoce como células fotosintéticas o fotoautótrofas. ANABOLISMO Y CATABOLISMO Tipos de organismos con nutrición autótrofa Metabolismo celular La QUIMIOSÍNTESIS se realiza en organismos procariontes capaces de utilizar la energía química que obtienen de la oxidación de productos inorgánicos gracias a la acción de sistemas enzimáticos muy especializados. ANABOLISMO Y CATABOLISMO Organismos heterótrofos La nutrición heterótrofa, Metabolismo celular de las raíces griegas hetero (diferente), y tropos (alimentar), es aquella que efectúan las células y los organismos incapaces de fabricar por sí mismos sus alimentos. De acuerdo con las particularidades que presentan los distintos tipos de nutrición heterótrofa, a ésta se le divide en: holozoica, saprofita y parásita. Un organismo unicelular holozoico el Dinidium sp. (a la izquierda) está a punto de devorar a un Paramecium sp. Hongos saprofitos como el Penicilium sp., son responsables de la putrefacción y “reciclamiento” de la materia orgánica. Parásitos como la tenia humana o solitaria (Taenia solium) que se aloja en el intestino, se alimentan de los nutrimientos que el huésped consume, con lo que causa daños. Metabolismo celular RESPIRACIÓN CELULAR Y PRODUCCIÓN DE ENERGÍA ¿Qué es lo que proporciona combustible a la vida dentro de las células? EL ATP Una célula solo contiene ATP suficiente como para sostener de 30 segundos a unos minutos de actividad normal. Debido a su alta energía potencial, el ATP es inestable y no se almacena. Como consecuencia, la mayoría de las células está fabricando ATP todo el tiempo. ¿Qué ocurre cuando se oxida la glucosa? Cuando la glucosa sufre la reacción incontrolada de oxidación que denominamos COMBUSTIÓN, parte de la energía potencial almacenada en sus enlaces químicos se convierte en energía cinética, en forma de calor y luz. Se libera 685 Kcal En nuestras células no ocurre la combustión. La glucosa se oxida mediante una larga serie de reacciones redox cuidadosamente controladas. En lugar de liberar toda la energía en calor, buena parte de ella se utiliza para fabricar ATP. ➢ La respiración celular y la combustión oxidan completamente la glucosa hasta CO2 y H2O ➢ La fermentación no oxida completamente la glucosa. Metabolismo celular RESPIRACIÓN CELULAR Y PRODUCCIÓN DE ENERGÍA La OXIDACIÓN DE LA GLUCOSA mediante la RESPIRACIÓN CELULAR se puede considerar un proceso de 4 pasos utilizado para convertir la energía química de la glucosa en energía química del ATP. 1. GLICÓLISIS. En este paso se descompone la glucosa de 6 carbonos a dos moléculas de piruvato. Durante este proceso se produce ATP a partir de ADP y la dinucleótido de nicotinamida y adenina (NAD+) se reduce para formar NADH. 2. PROCESAMIENTO DEL PIRUVATO. El piruvato se procesa para formar una molécula de CO2, y los dos carbonos restantes se utilizan para formar el compuesto acetil CoA. La oxidación del piruvato hace que NAD+ se reduzca a NADH. 3. CICLO DEL ÁCIDO CÍTRICO. Cada acetil CoA se oxida para obtener dos moléculas de CO2. Durante esta secuencia de reacciones se produce más ATP y NADH, y el dinucleótido de flavina y adenina (FAD) se reduce para formar FADH2. 4. TRANSPORTE DE ELECTRONES Y FOSFORILACIÓN OXIDATIVA. Los electrones del NADH y FADH2 se mueven a través de una serie de proteínas denominada cadena de transporte de electrones (ETC). La energía liberada en esta cadena de reacciones redox se utiliza para crear un gradiente de protones a través de una membrana; el consiguiente flujo de vuelta de protones a través de la membrana se emplea para fabricar ATP. Dado que este modo de producción de ATP enlaza la fosforilación del ADP con la oxidación del NADH y el FADH2 se denomina fosforilación oxidativa. Metabolismo celular RESPIRACIÓN CELULAR Y PRODUCCIÓN DE ENERGÍA Metabolismo celular RESPIRACIÓN CELULAR Y PRODUCCIÓN DE ENERGÍA La respiración celular oxida la glucosa para formar ATP Cada componente produce moléculas de alta energía en la forma de nucleótidos (ATP) y/o portadores electrónicos (NADH o FADH2). Como los 4 componentes están conectados, la oxidación de la glucosa es una vía metabólica integrada. Los tres primeros pasos oxidan la glucosa para producir NADH y FADH2 que luego alimentan la cadena transportadora de electrones. Metabolismo celular RESPIRACIÓN CELULAR Y PRODUCCIÓN DE ENERGÍA La respiración celular interacciona con otras vías catabólicas y anabólicas Diversos compuestos de alta energía de los hidratos de carbono, grasas o proteínas pueden descomponerse mediante reacciones catabólicas y emplearse en la respiración celular para la producción de ATP. Varios de los productos intermedios de la respiración celular actúan como moléculas precursoras en reacciones anabólicas que conducen a la síntesis de hidratos de carbono, nucleótidos, lípidos y aminoácidos. Metabolismo celular RESPIRACIÓN CELULAR Y PRODUCCIÓN DE ENERGÍA Vía de la glucólisis Esta secuencia de 10 reacciones oxida la glucosa a piruvato. Cada reacción es catalizada por una enzima diferente para producir 2 ATP netos (se producen 4 ATP, pero 2 se gastan), dos moléculas de NADH y dos moléculas de piruvato. En el paso 4, la fructosa-1,6bifofostato se divide en 2 productos que son procesados en los pasos 6 a 10. Las cantidades indicadas para “lo que entra” y “lo que sale” son los totales combinados para ambas moléculas. 1. Hexocinasa 2. Fosfoglucosa isomerasa 3. Fosfofructocinasa 4. Fructosa-bifosfato aldolasa 5. Triosa fosfato isomerasa 6. Gliceraldehido-3-fosfato deshidrogenasa 7. Fosfoglicerato cinasa 8. Fosfoglicerato mutasa 9. Enolasa 10. Piruvato cinasa Metabolismo celular RESPIRACIÓN CELULAR Y PRODUCCIÓN DE ENERGÍA La fosforilación a nivel de sustrato implica a una enzima y a un sustrato fosforilado La fosforilación a nivel de sustrato ocurre cuando una enzima cataliza la transferencia de un grupo fosfato de un sustrato fosforilado a ADP, formando ATP. La fosfofructocinasa tiene dos lugares de unión para el ATP Modelo de una de las cuatro subunidades idénticas de fosfofructocinasa. En el sitio activo, el ATP se utiliza como sustrato para transferir uno de sus grupos fosfato a la fructosa6-fosfato. En el sitio regulador, la unión del ATP inhibe la reacción, cambiando la forma de la enzima. Metabolismo celular RESPIRACIÓN CELULAR Y PRODUCCIÓN DE ENERGÍA Metabolismo celular RESPIRACIÓN CELULAR Y PRODUCCIÓN DE ENERGÍA Estructura de la mitocondria Procesamiento del piruvato El piruvato se oxida a acetil CoA La reacción mostrada está catalizada por la piruvato deshidrogenasa. En el proceso, un carbono (en el diagrama, en rojo) se oxida completamente para formar CO2 y se libera. Metabolismo celular RESPIRACIÓN CELULAR Y PRODUCCIÓN DE ENERGÍA El ciclo del ácido cítrico completa la oxidación de la glucosa Aconitasa El acetil CoA entra el el ciclo de Krebs y salen CO2, NADH, FADH2 y GTP o ATP. El GTP o ATP se producen por fosforilación a nivel de sustrato. Isocitrato deshidrogenasa Isocitrato sintasa El ciclo del ácido cítrico se regula mediante inhibición por retroalimentación α-cetoglutarato deshidrogenasa Malato deshidrogenasa Fumarasa ATP: regulador alostérico NADH: inhibidor competitivo El ciclo de Krebs se ralentiza cuando el ATP y el NADH abundan Succcinil CoA sintetasa Succinato deshidrogenasa Metabolismo celular RESPIRACIÓN CELULAR Y PRODUCCIÓN DE ENERGÍA Metabolismo celular RESPIRACIÓN CELULAR Y PRODUCCIÓN DE ENERGÍA La oxidación de la glucosa produce ATP, NADH, FADH2 y CO2 La glucosa se oxida completamente a dióxido de carbono mediante la glucólisis, el procesamiento del piruvato y el ciclo del ácido cítrico Metabolismo celular RESPIRACIÓN CELULAR Y PRODUCCIÓN DE ENERGÍA Variación de la energía libre a medida que se oxida la glucosa ✓ Se liberan 685 Kcal/mol de energía libre de la oxidación de la glucosa. ✓ Buena parte de la energía se aprovecha en forma de ATP, NADH y FAH2. ✓ Los óvalos verdes numerados identifican los pasos de reacción en la glucólisis y el ciclo del ácido cítrico. Basándose en los datos de esta gráfica, ¿Cuál de las tres moléculas de alta energía producidas durante la oxidación de la glucosa cabría esperar que transporte la mayor cantidad de energía química? Metabolismo celular RESPIRACIÓN CELULAR Y PRODUCCIÓN DE ENERGÍA En una cadena de transporte de electrones tiene lugar una serie de reacciones de oxidaciónreducción La energía potencial en los electrones compartidos disminuye desde los transportadores de electrones NADH y FADH2 a través de una cadena transportadora de electrones hasta un aceptor final de electrones. En la cadena de transporte de electrones, el oxígeno es aceptor de electrones y forma agua como subproducto. La variación global de energía libre de 52 kcal/mol (desde el NADH hasta el oxígeno) se descompone en una serie de pequeños pasos. Metabolismo celular RESPIRACIÓN CELULAR Y PRODUCCIÓN DE ENERGÍA Metabolismo celular RESPIRACIÓN CELULAR Y PRODUCCIÓN DE ENERGÍA ¿Cómo funciona la cadena transportadora de electrones? Los componente individuales de la cadena transportadores de electrones mostrados en la figura se encuentran en la membrana interior de las mitocondrias. Q y el citocromo c transportan electrones de un complejo a otro: Q también bombea protones a través de la membrana. La flecha de color naranja indica el movimiento de Q entre un sitio y otro, desde la matriz mitocondrial al espacio intermembranoso. Si el transporte de electrones no fabrica ATP, ¿QUÉ OTRA COSA LO HACE? Añada una flecha que atraviesa la membrana y etiquétela como “gradiente de protones”. En los recuadros de la parte inferior, enumere “lo que entra” y “lo que sale” para cada complejo. Metabolismo celular RESPIRACIÓN CELULAR Y PRODUCCIÓN DE ENERGÍA Evidencia de la hipótesis quimiosmótica INVESTIGACIÓN CUESTIÓN: ¿cómo están vinculadas la cadena de transporte de electrones y la producción de ATP? 3. Iluminar la vesícula, de modo que la bacteriorrodopsina bombee protones hacia fuera de la vesícula, creando un gradiente de protones. HIPÓTESIS QUIMIOSMÓTICA: la conexión es indirecta. La CTE crea un gradiente protónico y la ATP sintasa utiliza el gradiente para sintetizar ATP. HIPÓTESIS ALTERNATIVA: la conexión es directa. Se necesitan proteínas de CTE específicas para la síntesis de ATP por la ATP sintasa 1. Crear vesículas a partir de membranas artificiales; añadir ATP sintasa, una enzima presente en las mitocondrias. PREDICCIÓN DE LA HIPÓTESIS QUIMIOSMÓTICA: se producirá ATP dentro de la vesícula. PREDICCIÓN DE LA HIPÓTESIS ALTERNATIVA: no se producirá ATP sin CTE RESULTADOS: Se produce ATP dentro de la vesícula en ausencia de la cadena de transporte de electrones ATP sintasa Bacteriorrodopsina 2. Añadir bacteriorrodopsina, una proteína que funciona como una bomba de protones activada por la luz CONCLUSIÓN: la conexión entre el transporte de electrones y la síntesis de ATP es indirecta; la síntesis de ATP solo requiere un gradiente de protones Metabolismo celular RESPIRACIÓN CELULAR Y PRODUCCIÓN DE ENERGÍA La fosforilación oxidativa necesita el motor de la ATP sintasa y un gradiente de protones La ATP sintasa tiene dos componentes principales, designados como F0 y F1, conectados por un eje. La unidad F0 gira cuando los protones pasan a su través. El eje transmite la rotación a la unidas F1, para dar lugar a la fabricación de ATP a partir de ADP y Pi. producción de ATP enlaza la fosforilación del ADP con la oxidación del NADH y el FADH2 se denomina fosforilación oxidativa. Metabolismo celular RESPIRACIÓN CELULAR Y PRODUCCIÓN DE ENERGÍA Metabolismo celular RESPIRACIÓN CELULAR Y PRODUCCIÓN DE ENERGÍA Producción de ATP durante la respiración celular La producción real de ATP por cada glucosa (29 ATP) es menor que el cálculo teórico (38 ATP) debido a la energía requerida para importar el NADH desde el citoplasma y debido al uso de la fuerza protónica para transportar activamente Pi hacia la matriz mitocondrial Metabolismo celular RESPIRACIÓN CELULAR Y PRODUCCIÓN DE ENERGÍA Respiración celular El oxígeno es el aceptor de electrones utilizado por todos los eucariotas y una gran diversidad de procariotas. Las especies que dependen del oxígeno como aceptor de electrones para la CTE utilizan respiración AERÓBICA y reciben el nombre de organismos aeróbicos. CTE: Cadena transportadora de electrones Las células que dependen de otros aceptores de electrones distintos del oxígeno se dice que utilizan respiración ANAERÓBICA. NO3-, SO42-, H2, H2S, CH4 u otros compuestos inorgánicos como donantes de electrones en lugar de la glucosa. En bacterias y arqueas, la CTE y la ATP sintasa se localizan en la membrana plasmástica. Metabolismo celular RESPIRACIÓN CELULAR Y PRODUCCIÓN DE ENERGÍA La respiración celular y la fermentación son vías alternativas para producir energía Si no hay ningún aceptor de electrones para mantener la ETC operando, el piruvato sufre una serie de reacciones conocidas con el nombre de fermentación La fermentación regenera NAD+ de modo que la glucólisis pueda continuar Metabolismo celular FOTOSÍNTESIS La fotosíntesis utiliza la luz solar para fabricar hidratos de carbono Los organismos fotosintéticos no pueden almacenar la energía electromagnética de la luz salvo la conviertan en otra forma. En la fotosíntesis, la energía de la luz del sol se transforma en energía química en los enlaces C-C y C-H de los hidratos de carbono. La reacción global se simplifica así: CO2 + H2O + ENERGÍA LUMÍNICA → (CH2O)n +O2 Transferencia de electrones durante la reducción de la glucosa Metabolismo celular FOTOSÍNTESIS La fotosíntesis tiene dos componentes relacionados En las reacciones fotosintéticas que captan la luz, la energía lumínica se transforma en energía química en forma de ATP y NADPH. Durante el ciclo de Calvin, se emplean el ATP y el NADPH producidos en las reacciones que capturan la luz, para reducir el dióxido de carbono a azúcar. Metabolismo celular FOTOSÍNTESIS Los pigmentos fotosintéticos absorben la luz La luz solar incluye la luz blanca, compuestas por todas las longitudes de onda de la parte visible del espectro electromagnético a la vez. Cuando un fotón incide sobre un objeto, el fotón puede absorberse, transmitirse o reflejarse. Una molécula de pigmento absorbe fotones de longitudes de onda concretas. Metabolismo celular FOTOSÍNTESIS Los pigmentos fotosintéticos contienen estructuras en anillo Existe una fuerte correlación entre el espectro de absorción de los pigmentos y el espectro de acción de la fotosíntesis a. Aunque las clorofilas a y b son muy similares estructuralmente, tienen los espectros de absorción característicos (ver figura anterior) b. El β-caroteno es un pigmento naranja que se encuentra en la raíz de la zanahoria y en tejidos de otras plantas. Metabolismo celular FOTOSÍNTESIS El esquema Z conecta los fotosistemas I y II El esquema Z propone que los electrones del agua son primero energizados por el fotosistema II para generar ATP y luego por el fotosistema I para reducir NADP+ a NADPH Metabolismo celular FOTOSÍNTESIS Los electrones pasan del agua al NADP+ a través de una vía lineal En la membrana tilacoide, el fotosistema II utiliza la luz para proporcionar energía a electrones extraídos del agua y los pasa a través de una CTE que incluye las plastoquinona (PQ), complejo citocromo y las plastocianinas (PC). La CTE produce una fuerza protónica que se utlliza para fabricar ATP. El fotosistema I proporciona energía a los electrones a partir de la PC y los pasa a ferredoxina, con el fin de reducir NADP+ a NADPH. Metabolismo celular FOTOSÍNTESIS El dióxido de carbono se reduce en el ciclo de Calvin Se muestran el número de reactivo y productos resultantes de tres ejecuciones de ciclo. De los seis G3P que se generan durante la fase de reducción, uno se emplea en la síntesis de glucosa o fructosa y los otros cinco se usan para regenerar la RuBP. Las tres RuBP que se regeneran participan en las reacciones de fijación correspondientes a ejecuciones adicionales del ciclo RuBP: bifosfato de ribulosa; 3PGA: 3-fosfoglicerato; G3P: gliceraldehido-3-fosfato Movilidad intracelular DISTRIBUCIÓN DE PROTEÍNAS Una "hoja de ruta" simplificada del tráfico de proteínas dentro de un célula eucariota Los orgánulos delimitados por membranas importan proteínas por uno de tres mecanismos posibles Movilidad intracelular DISTRIBUCIÓN DE PROTEÍNAS INVESTIGACIÓN CUESTIÓN: ¿contiene la proteína nucleoplasmina una señal de “enviar a núcleo” HIPÓTESIS: La nucleoplasmina contiene una señal discreta “enviar al núcleo” que reside en la cola o la región central. HIPÓTESIS NULA: La nucleoplasmina no requiere una señal para entrar en el núcleo, o bien toda la proteína sirve como señal Movilidad intracelular DISTRIBUCIÓN DE PROTEÍNAS Algunas secuencias señales típicas Movilidad intracelular DISTRIBUCIÓN DE PROTEÍNAS Las secuencias señal conducen a las proteínas al orgánulo correspondiente Proteína citosólica (con secuencia señal) Proteína del RE con secuencia señal eliminada secuencia señal del RE adherida a la proteína citosólica Proteína del RE Secuencia señal del RE A. NORMAL Proteína citosólica con secuencia señal del RE B. SECUENCIAS SEÑAL INTERCAMBIADAS Movilidad intracelular DISTRIBUCIÓN DE PROTEÍNAS Estructura de la envoltura nuclear y complejo del poro nuclear Las subunidades ribosómicas y varios tipos de ARN salen del núcleo; los nucleótidos y determinadas proteínas entran en él. En una célula típica, más de 500 moléculas atraviesan cada uno de los 3000-4000 poros nucleares por segundo. El nivel de tráfico a través de los poros nucleares es extraordinario. ¿Cómo se dirige y regula? Movilidad intracelular DISTRIBUCIÓN DE PROTEÍNAS Las proteínas destinadas al núcleo se transportan en forma activa a través de los poros nucleares Cómo la hidrólisis de GTP por Ran en el citosol proporciona direccionalidad al transporte nuclear Receptores de importación nuclear (importinas) Movilidad intracelular DISTRIBUCIÓN DE PROTEÍNAS Las proteínas se importan al interior de una mitocondria en forma desplegada Translocación de proteínas precursoras del cloroplasto en el espacio tilacoide Movilidad intracelular DISTRIBUCIÓN DE PROTEÍNAS La síntesis de las proteínas del citosol y la de los orgánulos delimitados por membranas (ej. RE) ocurre en un conjunto común de ribosomas Translocación co-traduccional y postraduccional de proteínas Movilidad intracelular DISTRIBUCIÓN DE PROTEÍNAS La hipótesis señal explica como las proteínas destinadas a la secreción entran en el sistema endomembranoso Según la hipótesis de la señal, las proteínas destinadas a ser secretadas contienen un pequeño fragmento de aminoácidos que interacciona con una partícula de reconocimiento de la señal (SRP) en el citosol. Esta interacción dirige la síntesis del resto de la proteína hacia la luz del RE rugoso. Movilidad intracelular DISTRIBUCIÓN DE PROTEÍNAS Una secuencia señal para RE y una para SRP dirigen el ribosoma hacia la membrana La SRP se une a la secuencia señal expuesta y al ribosoma y, por este motivo, torna más lenta la síntesis de proteica. A continuación, el complejo SRP-ribosoma se une al receptor de la SRP en la membrana del RE. Se libera entonces la SRP, en tanto el ribosoma pasa a un canal de translocación proteica en la membrana del RE. Por último, el canal de translocación inserta la cadena polipeptídica en la membrana y comienza a transferirla a través de la bicapa lipídica. partícula de reconocimiento de la señal (SRP) Movilidad intracelular DISTRIBUCIÓN DE PROTEÍNAS Una proteína soluble cruza la membrana del RE e ingresa en la luz cadena polipeptídica que emerge del ribosoma secuencia señal canal de translocación péptido señal escindido canal de translocación cerrado peptidasa señal proteína soluble madura en la luz del RE Una proteína transmembrana de paso doble emplea una secuencia de comienzo de transferencia interna al integrarse en la membrana del RE En estas figuras se omitió el ribosoma unido a la membrana Una proteína transmembrana de paso único se integra en la membrana del RE TRANSPORTE VESICULAR Las vesículas brotan de una membrana y se fusionan con otra; transportan los componentes de la membrana y las proteínas solubles entre los compartimientos celulares Las moléculas de clatrina forman jaulas similares a canastas que ayudan a dar a las membranas la forma de vesículas. Transporte vesicular TRANSPORTE VESICULAR Uso de diferentes capas para diferentes pasos en el tráfico de vesículas Electron micrographs of clathrin-coated, COPI-coated, and COPII-coated vesicles. COP: Coat protein TRANSPORTE VESICULAR Vesículas recubiertas con clatrina transportan moléculas de carga seleccionadas Las moléculas de la proteína DINAMINA hidrolizan GTP que tienen unido para desprender la vesícula TRANSPORTE VESICULAR RECONOCIMIENTO INICIAL Las proteínas Rab y las proteínas SNARE ayudan a dirigir las vesículas de transporte a sus membranas diana ACOPLAMIENTO FUSIÓN Las proteínas Rab están asociadas a GTP ACOPLAMIENTO DE LA VESÍCULA DE TRANSPORTE membrana diana UNIÓN DE MEMBRANA membrana diana FUSIÓN DE MEMBRANAS Las proteínas SNARE desempeñan una función central en la fusión de la membrana VÍAS SECRETORAS Muchas proteínas son glucosiladas en el RE VÍAS SECRETORAS El tamaño del RE es controlado por la cantidad de proteína que fluye a través de él Las chaperonas previenen que las proteínas mal plegadas o ensambladas en forma parcial abandonen el RE Las proteínas mal plegadas en la luz del RE activan la producción de chaperonas y la expansión del RE Proteína malenovelada: proteína mal plegada Proteína enovelada apropiadamente: proteína plegada correctamente VÍAS SECRETORAS Las proteínas se modifican y se distribuyen aun más en el complejo de Golgi Man, mannose; GlcNAc, N-acetylglucosamine; Gal, galactose; NANA, N-acetylneuraminic acid (sialic acid). VÍAS SECRETORAS Las proteínas secretoras se liberan de la célula por exocitosis En las células secretoras las vías reguladas y constitutivas de la exocitosis divergen en la red trans de Golgi Proteínas solubles recién sintetizadas para la secreción constitutiva Lípidos de la membrana plasmática recién sintetizados Vesícula de transporte SECRECIÓN CONSTITUTIVA Fusión de la membrana no regulada Las vesículas secretoras empaquetan y descargan cúmulos concentrados de proteínas Proteína de la membrana plasmática recién sintetizada Red trans de Golgi Membrana plasmática Hormona o neurotransmisor que actúan como señal Transducción de señal SECRECIÓN REGULADA Complejo de Golgi Vesícula secretora que almacena proteínas de secreción Fusión de la membrana regulada CITOSOL ESPACIO EXTRACELULAR VÍAS ENDOCÍTICAS Las células fagocíticas especializadas ingieren partículas grandes Células fagocíticas que ingieren otras células VÍAS ENDOCÍTICAS Las líquidos y las macromoléculas son captados por pinocitosis Formación de vesículas recubiertas con clatrina de la membrana plasmática Representación esquemática de la macropinocitosis Caveola en la membrana plasmática de un fibroblasto VÍAS ENDOCÍTICAS La endocitosis mediada por receptor proporciona una vía específica hacia el interior de las células animales La LDL ingresa en la célula por medio de la endocitosis mediado por receptores ENDOCITOSIS REGRESO DE LOS RECEPTORES DE LDL A LA MEMBRANA PLASMÁTICA PÉRDIDA DE LA CUBIERTA endosoma DESPRENDIMIENTO DE LAS VESÍCULAS DE TRANSPORTE FUSIÓN CON EL ENDOSOMA TRANSFERENCIA AL LISOSOMA Colesterol libre lisosoma Enzimas hidrolíticas VÍAS ENDOCÍTICAS Las macromoléculas endocitadas se distribuyen en los endosomas 1. Reciclado Endosoma temprano Vesícula de transporte Unión estrecha 2. Degradación Lisosoma 3. Transcitosis Membrana plasmática basolateral Núcleo Los lisosomas son los sitios principales de la digestión intracelular VÍAS ENDOCÍTICAS Existen tres vías para reciclar material en el lisosoma La endocitosis y la fagocitosis mediadas por receptor implican acarrear material desde el exterior y rodearlo con una bicapa lipídica desde la membrana plasmática. Los endosomas maduran en lisosomas o, como los fagosomas, se fusionan con los lisosomas existentes. En la autofagia, el material del citoplasma se encapsula con una membrana interna antes de fusionarse con el lisosoma. BIBLIOGRAFÍA CONSULTADA 1. Alberts B, Bray D, Hopkin K, Jhonson A, Lewis J, Raff M, et al. Introducción a la biología celular. 3ª ed. Buenos Aires: Editorial Médica Panamericana; 2011. 2. Alberts B, Johnson A, Lewis J, Morgan D, Raff M, Roberts K, et al. Molecular biology of the cell. Sixth edition. New York: Garland Science, Taylor & Francis Group; 2015 3. Audesirk T, Audesirk G, Byers BE. Biología. La vida en la Tierra con Fisiología. Décima edición. México D.F.: Pearson Educación de México S.A.; 2017 4. Freeman S, Quillin H, Allison L, Blach M, Podgorski, Taylor E. Fundamentos de Biología. 6ta ed. Madrid: Pearson Educación S.A.; 2019. 5. Gama M, Fernández D. Biología 1. Cuarta edición. México D.F.: Pearson Educación de México S.A.; 2018. GRACIAS