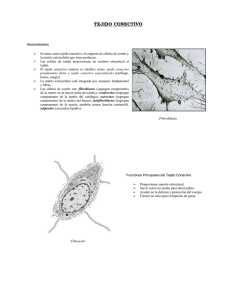
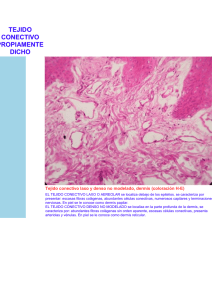
1 UNCPBA Facultad de Ciencias Veterinarias Departamento de Ciencias Biológicas CURSO DE HISTOLOGÍA, EMBRIOLOGIA Y TERATOLOGIA GUIA DE ESTUDIO: TEJIDO CONECTIVO AUTOR M.V. M Sc. Ricardo Alzola AÑO 2002 Introducción: El término tejido conectivo o conjuntivo incluye tejidos conocidos como tejido conectivo propiamente dicho y un subgrupo de tejidos conectivos especiales. El tejido conectivo propiamente dicho forma un compartimiento extenso y contínuo en todo el organismo limitado por la lámina basal de los diferentes epitelios de revestimiento y de las glándulas que secretan hacia la superficie. El tipo de desarrollo que siguen las células de tejido conectivo especializado reflejará su función. Si la función de estos tejidos conectivos especiales son mecánicas, predominan las fibras extracelulares y la sustancia fundamental; es el caso de los ligamentos, tendones, cartílagos y hueso. Si las funciones especializadas son de protección (reacciones inflamatorias e inmunológicas) o de conservación de las reservas energéticas, las células son las que predominan, es el caso del tejido linfático y del adiposo. El tejido conectivo se caracteriza morfológicamente por presentar diversos tipos de células, separadas por abundante material intercelular sintetizado por ellas, es una estructura polimorfa y compleja que presenta una notable dinámica funcional estando ampliamente distribuido por el organismo. El material intercelular está representado por las fibras y por la sustancia amorfa. 2 Clasificación: Las variaciones regionales permiten clasificar al tejido conectivo en: - Tejido Conectivo Laxo Tejido Conectivo Denso: modelado y no modelado Tejido conectivo de Propiedades Especiales: - Adiposo - Elástico - Reticular - Hemocitopoyético Linfoide y Mieloide - Mucoso - Cartilaginoso - Oseo. Funciones: Las funciones más importantes del tejido conectivo son: - Sostén y relleno: Los tejidos epitelial y muscular están asociados al tejido conectivo que les sirve de soporte, rellena los espacios entre las células. Las fibras colágenas constituyen trabéculas y tabiques en el interior de distintos órganos, constituyendo el estroma. - Almacenamiento: Al almacenar lípidos representa una notable reserva nutritiva. Por su riqueza en mucopolisacáridos almacena agua y electrolitos. También almacena proteínas. Se calcula que 1/3 de las proteínas plasmáticas del organismo se encuentra en los espacios intercelulares del tejido conectivo. Este tejido contiene células fagocitarias y células productoras de anticuerpos. - Reparación: Las áreas de conectivo destruidas por la inflamación o por lesión traumática son reconstruídas nuevamente por la proliferación del conectivo adyacente.. - Trasporte: El tejido conectivo trasporta sustancias nutritivas de los capilares sanguíneos a los diversos tejidos. Trasporta también productos de desecho del metabolismo en el sentido inverso. Origen: Los tejidos conectivos se originan del mesodermo y específicamente del tejido mesenquimático, formado por células mesenquimáticas sumergidas en abundante sustancia intercelular amorfa poco viscosa. La célula mesenquimática es una célula grande, de citoplasma irregular con forma de estrella, núcleo voluminoso de cromatina laxa y nucléolo marcado. El mesénquima se propaga por el interior del embrión envolviendo los órganos en formación y penetrando en ellos. Además origina otros tejidos como el glandular y el muscular. 3 Tejido conectivo laxo propiamente dicho: Fijas o Propias: Fibroblastos Fibrocitos Reticulares Mesenquimáticas Adiposas Células Cebadas Plasmáticas Errantes Macrófagos Leucocitos Fibras compuestas por proteínas fibrosas Sust. Intercelular Material amorfo constituído por sustancias intercelulares macromoleculares no fibrosas dispuestas en forma de un gel amorfo. CÉLULAS Células fijas: Fibroblastos: Son las células que producen las fibras y la sustancia intercelular amorfa de los tejidos conectivos. Es la células más común, durante la etapa en que producen activamente sustancias intercelulares pueden poseer amplios procesos citoplasmáticos o presentan forma de huso. Tamaño: 20 a 30µ de largo por 10 µ de ancho. Núcleo: claro, grande, redondo u oval con cromatina finamente granular y nucléolo prominente. Citoplasma: poco visible. Mitocondrias numerosas y grandes. El RER y el AG están bien desarrollados. La ultraestructura de esta célula es característica de células que secretan proteínas de manera muy activa. Sintetizan procolágena, tropoelastina y proteoglucanos. Pueden liberar sus productos de secreción desde cualquier parte de su superficie celular y no solo de la apical (como las células epiteliales). Aunque consideradas células fijas del tejido conectivo los fibroblastos son capaces de efectuar algunos movimientos. Rara vez experimentan división celular, pero pueden hacerlo durante la cicatrización de heridas. Cuando estas células disminuyen su actividad se las denomina fibrocitos. 4 Fibrocitos: Cuando el fibroblasto disminuye su actividad se lo denomina fibrocito. Estos son incapaces de dividirse y por ello la restitución del tejido conectivo se efectúa mediante el crecimiento de jóvenes fibroblastos derivados principalmente de los pericitos. Tienden a ser fusiformes con pocas prolongaciones. Poseen un núcleo pequeño, alargado y más oscuro que el de los fibroblastos. Tamaño: 15 a 20µ por 6 a 7µ. Citoplasma: acidófilo, con un RER y AG poco desarrollados. Miofibroblastos: Son fibroblastos modificados. Histológicamente no se distinguen los filamentos de los miofibroblastos con facilidad mediante microscopia óptica. Con microscopia electrónica se observa que tienen haces de filamentos de actina y cuerpos densos semejantes a los de las células del músculo liso. Difieren de las células musculares lisas porque carecen de lámina externa. Son abundantes en zonas de cicatrización y en el ligamento periodontal. Reticulares: Son semejantes a los fibroblastos. Se observan en la médula ósea y órganos linfáticos. Citoplasma: alargado, con prolongaciones que contactan con las células vecinas formando una malla o retículo. Función: forman fibras reticulares, además poseen actividad fagocítica. Mesenquimáticas: Los histólogos clásicos postularon la existencia de células que retenían la multipotencialidad de las células mesenquimáticas embrionarias en el tejido conectivo laxo del adulto. Estas células fueron llamadas células mesenquimáticas indiferenciadas y se creian que originaban células diferenciadas cuyo fin era participar en los fenómenos de reparación a gran escala y en la formación de nuevo tejido, como neovasos sanguíneos o cartílago y huesos ectópicos. Uno de los precursores postulados de tales células es el pericito, también llamado célula adventicial o célula perivascular. Los pericitos se encuentran alrededor de los capilares y vénulas. Están rodeados por material de lámina basal que se continúa directamente con la lámina basal del endotelio capilar. El pericito envuelve parcialmente al capilar y a su núcleo.Tiene una configuración aplanada y curva que se adapta al contorno tubular del vaso. El citoplasma presenta escasas mitocondrias pequeñas, gran cantidad de ribosomas libres, algunas vesículas y pocas cisternas del RER. El pericito presenta prolongaciones laterales que se extienden desde el cuerpo celular y rodean parcialmente al capilar. Los pericitos poseen características de las células de músculo liso y de las células endoteliales, se cree que bajo ciertas condiciones pueden diferenciarse en otras células. Adiposas: Son células especializadas para sintetizar y almacenar lípidos. Se presentan aisladas y en grupos. 5 Cuando ocupan una gran extensión en el tejido conectivo laxo conforman el tejido adiposo.. Forma: esférica o poliédrica. Al acumularse gran cantidad de vacuolas lipídicas el núcleo resulta desplazado a la periferia y el citoplasma aparece ocupado por una gran vacuola de allí el nombre de célula en forma de “anillo de sello”. Tamaño: 130 µ. Origen: células mesenquimáticas o fibroblastos. Células errantes: Cebadas o Mastocitos: Están distribuidas ampliamente en pequeños grupos por el tejido conectivo particularmente cerca de los vasos sanguíneos pequeños. Se encuentran en el tejido conectivo subepitelial de los sistemas respiratorios y digestivos y en la piel. Mientras las meninges contienen mastocitos el tejido conectivo perivascular dentro de la sustancia encefálica y médula carece de ellas, lo que protege al SNC de los efectos potencialmente nocivos del edema característico de las reacciones alérgicas. El timo y en grado menor otros órganos hemocitopoyéticos con excepción del bazo, poseen gran cantidad de mastocitos. Forma y tamaño: es variable entre las distintas especies (20 a 30 µ). En general son grandes, redondeadas u ovoides sin prolongaciones. Presentan el citoplasma lleno de gránulos basófilos que se tiñen intensamente. Núcleo: esférico, central y pálido. Pueden ser binucleadas. Con frecuencia no es visible por estar cubierto por los gránulos citoplasmáticos. Origen: Las células precursoras de los mastocitos se originan probablemente en la médula ósea, circulan por un tiempo breve en sangre y posteriormente entran al tejido conectivo en el que se diferencian en mastocitos y adquieren sus gránulos característicos. Tienen un lapso de vida de unos cuantos meses y a veces experimentan división celular. Citoplasma: contienen numerosas mitocondrias, cierta cantidad de RER y prominente AG. Los mastocitos son difíciles de observar en las preparaciones teñidas con H/E, pero se destacan fácilmente en los cortes coloreados con azul de toluidina, que tiñe los gránulos de color rojo, esta capacidad de modificar la coloración del colorante utilizado se denomina “metacromasia”, es debida a la presencia de numerosos radicales ácidos en las estructuras. Lo más evidente de su citoplasma es su contenido de gránulos secretorios grandes (0.2 a 0.8 nm), c/u de ellos está encerrado en una membrana. Los gránulos están constituídos principalmente por glucoproteínas ácidas y neutras. Las sustancias que liberan son: 1. Histamina 2. Sustancia de reacción lenta de la anafilaxia (Leucotrieno C). 3. Factor quimiotáctico para eosinófilos 4. Heparina 5. Proteasas neutras 6 6. Arilsulfatasa 7. Factor quimiotáctico para neutrófilos. 1 y 2: estas sustancias, en la respuesta inmune, aumentan la permeabilidad vascular y por lo tanto producen edema de los tejidos. 3: estimula a los eosinófilos para que migren hacia los sitios donde están los mastocitos (los eosinófilos contrarrestan los efectos de la Histamina y de la sustancia de reacción lenta) 4:es un anticoagulante e interviene en el metabolismo de los lípidos al activar la lipasa de lipoproteína. Plasmáticas: Son células poco frecuentes en el conectivo laxo a excepción de la mucosa gastrointestinal, urogenital y respiratoria. Origen: de los linfocitos B. Ubicación: se encuentran en los tejidos hemocitopoyético linfoide y mieloide. Forma: ovoide Núcleo: es esférico u ovoide La cromatina se observa en grumos compactos y toscos que se alternan con áreas claras de igual tamaño, lo que da frecuentemente un aspecto similar a los radios de una rueda de carro. Se ubica excéntricamente. Citoplasma: muy basófilo por su abundante RER. El AG se encuentra al lado del núcleo. Hay abundantes mitocondrias. Estas células contienen los cuerpos o inclusiones de Russell que son cuerpos densos o inclusiones de 2 a 3 µ de diámetro y de función desconocida. Presentan una intensa síntesis de proteínas, porque producen anticuerpos. Los anticuerpos son proteínas específicas de la clase de las gammaglobulinas fabricadas por el organismo en respuesta a la penetración de moléculas extrañas que reciben el nombre de antígenos. Cada anticuerpo formado es específico para el antígeno que provoca su formación y se combina con el mismo. Macrófagos: Son células pleomorficas, que en reposo tienen una forma de huso mientras que cuando migran la tienen oval o redondeada con prolongaciones citoplasmáticas. Los histólogos creyeron que los macrófagos derivaban de células precursoras del Sistema Retículo Endotelial, que incluía tanto a las células no fagocíticas como a los reticulocitos. Más recientemente se sustituyó esta clasificación por la de Sistema de Fagocitos Mononucleares. Origen: Los componentes del Sistema de Fagocitos mononucleares se originan en una célula madre común en la médula ósea, poseen lisosomas, son capaces de efectuar fagocitosis y ponen de manifiesto receptores FC y receptores para complemento. Esta célula madre origina a los monocitos que circulan por la sangre. Al recibir la señal apropiada dejan la sangre y migran por el endotelio de los capilares o las vénulas. En el tejido conectivo maduran, se convierten en macrófagos y poseen una vida media de 2 meses. Los macrófagos de ciertas regiones reciben nombres específicos: - células de Kupffer: se ubican en el hígado - células polvorosas: “ “ “ “ pulmón - células de Langerhans: “ “ “ la piel 7 - Monocitos: tejdo sanguíneo. Osteoclastos: del hueso Microglia: del SNC. Macrófagos del tejido conectivo, bazo, ganglio linfático, timo, médula ósea. Tamaño: varía según la especie entre 10 y 20 µ. Pueden ser fijos o libres. Los fijos se denominan histiocitos. Los libres son los que migran por medio de movimientos ameboideos. Los fijos son fusiformes o estrellados. De núcleo ovoide. En el tejido conectivo laxo los macrófagos fijos son casi tan numerosos como los fibroblastos. Los libres son más activos en la fagocitosis que el fijo. El macrófago libre y fijo representa fases diversas del mismo tipo celular. Citoplasma: AG grande; RER, REL bien desarrollados; mitocondrias abundantes, vesículas secretorias y lisosomas. Los lisosomas son las estructuras que mejor indican la capacidad fagocítica especializada en esta célula. Además presenta vacuolas endocíticas y lisosomas secundarios. Los productos de secreción liberados por los macrófagos incluyen una variedad de sustancias relacionadas con la respuesta inmune, anafilaxia e inflamación. La liberación de proteasas neutras y GAGasas (enzimas que degradan los Glucosaaminoglucanos) facilita la migración de los macrófagos a través del tejido conectivo. La superficie de la célula presenta múltiples pliegues y prolongaciones digitiformes. Los repliegues de la superficie intervienen en el proceso de endocitosis, dado que rodean las sustancias a ser fagocitadas. Lo mismo sucede con estructuras de gran tamaño como ser otras células, los repliegues se extienden por la superficie de estas y luego las fagocitan. Función: Aunque la principal función del macrófago es la fagocitosis, bien como actividad de defensa (fagocitosis de bacterias) o como operación de limpieza (fagocitosis de detritos celulares) también cumpliría un papel en las reacciones inmunes al concentrar antígenos proteicos y celulares de materiales extraños fagocitados y presentarselos a los linfocitos. Por otro lado cuando los macrófagos encuentran cuerpos extraños de gran tamaño, se fusionan para formar enormes células de hasta 100 núcleos. Estas células multinucleadas reciben el nombre de “células gigantes de cuerpo extraño”. Leucocitos: Son glóbulos blancos de la sangre que se encuentran con frecuencia en el tejido conectivo. Provienen de la sangre por migración a través de la pared de los capilares y vénulas. Los leucocitos más frecuentes son los eosinófilos y los linfocitos. Además, pueden observarse neutrófilos, basófilos y monocitos. Los eosinófilos presentan gránulos citoplasmáticos acidófilos que son lisosomas y un núcleo bilobulado. En enfermedades alérgicas y por parásitos aumentan el número de esosinófilos en el tejido coectivo. Fagocitan los complejos antígeno-anticuerpo. Los linfocitos miden de 6 a 10 µ presentan un núcleo esférico con una leve escotadura, citoplasma basófilo y muy escaso. Se diferencian en plasmatocitos o células plasmáticas. 8 SUSTANCIA INTERCELULAR La mayoría de las células de los organismos pluricelulares están en contacto con una intrincada trama de macromoléculas extracelulares que constituyen la sustancia o matriz extracelular. Estas moléculas de proteínas y de polisacáridos se segregan de forma local y se ensamblan formando una trama organizada en el espacio extracelular de la mayoría de los tejidos. Además de actuar como cemento biológico universal también forman estructuras altamente especializadas como el cartílago, tendones, láminas basales, huesos y dientes. Las macromoléculas que constituyen la matriz extracelular están segregadas por células locales, especialmente fibroblastos, distribuidos en la matriz. En formas especializadas de la matriz como el cartílago y el hueso, son segregadas por los condroblastos y osteoblastos respectivamente. La matriz extracelular del tejido conectivo propiamente dicha está compuesta por una sustancia básica hidratada del tipo de los geles con fibras embebidas en ella. La primera de estas resiste a las fuerzas de compresión y la última soporta las fuerzas de tensión o tracción. El agua de la hidratación permite el intercambio rápido de nutrientes y productos de desecho trasportados por el líquido tisular a su paso por la sustancia básica. Las fibras son de tres tipos colágenas, elásticas y reticulares. La sustancia básica está compuesta por glucosaminoglucanos, proteoglucanos y glucoproteínas de adhesión. Las principales macromoléculas son: 1. Fibras 2. Sustancia básica: Polisacáridos Glucosaminoglucanos (GAG), que suelen estar unidos covalentemente a proteínas formando los Proteoglucanos (PG) y glucoproteínas de adhesión. Las moléculas de GAG y PG forman una “sustancia fundamental” gelatinosa, hidratada en la que están incluidas las fibras de colágena. 1.- Fibras: Las fibras de la matriz extracelular brindan resistencia tensil y elasticidad a esta sustancia. Los histólogos clásicos han descrito 3 tipos de fibras basados en su morfología y su reactividad con los colorantes histológicos. Los 3 tipos de fibras son de colágena, reticular y elástica. Colágena: Los grandes acúmulos de fibras de colágena parecen de color blanco resplandeciente en el individuo que está vivo. Los haces de fibras de colágena se conocen también como fibras blancas. Las fibras de colágena del tejido conectivo suelen medir menos de 10 µm de diámetro y son incoloras. Al teñirse con hematoxilina y eosina se manifiestan como haces de fibras sonrosadas, onduladas y largas. Las largas fibras de colágena: refuerzan y contribuyen a organizar la matriz, la fase acuosa del gel polisacárido permite la difusión de nutrientes, metabolitos y hormonas entre sangre y tejidos. Las fibras colágenas constituyen una familia de proteínas fibrosas que se encuentran en todos los animales pluricelulares. Son las proteínas más abundantes en los mamíferos aproximadamente 25%. El rasgo principal de todas las moléculas de colágena es su rígida estructura helicoidal de 3 hebras. Se trata de 3 cadenas polipeptídicas de colágena denominadas “ cadenas α” que están enrolladas entre si formando una hélice regular, la cual constituye una molécula de colágena, en forma de cuerda de aproximadamente 300 nm de largo y 1,5 nm de diámetro. 9 Hasta el momento se han definido detalladamente 7 cadenas α, genéticamente distintas, y de unos 1000 residuos de aminoácidos c/u,. a partir de las diferentes combinaciones se han descrito 15 tipos diferentes de moléculas de colágena, según la secuencia de aminoácidos de sus cadenas α solo 6 de ellas tienen un interés primordial. Cada cadena α es codificada por un mRNA separado. Estos tipos diferentes de colágena estan localizados en regiones específicas del cuerpo en las que desempeñan funciones diversas. Tipos de colágena: I: El tipo más frecuente forma fibras gruesas y se encuentra en el tejido conectivo propiamente dicho, hueso, diente (dentina y cemento). II: Forma fibras más delgadas y se encuentra casi exclusivamente en las matrices de los cartílagos hialino y elástico. III: Se conocen también como fibras reticulares porque se creía que difería de la colágena. En la actualidad se sabe que la fibra reticular es un tipo de colágena que se glucosila en grado elevado y forma fibras delgadas o redes. Se encuentran como un reticulado fino muy relacionado con las células. Rodean a los adipocitos, a las células del músculo liso, se encuentran por debajo del endotelio de los capilares a los que dan cierta rigidez. Forman el retículo del tejido linfoide y de la médula ósea y rodean a las células parenquimatosas de las glándulas. A causa del gran contenido de grupos carbohidratos las fibras de colágena tipo III se tiñen de manera preferencial con sales argénticas o por medio de la reacción del ácido peryódico de Schiff (PAS). IV: No forma fibras y no manifiesta periodicidad de 67 nm. Forma una red de moléculas de procolágeno que están agregados unos contra otros para formar un tapete de sostén de la lámina basal. V: Forma fibrillas muy delgadas, posee periodicidad de 67 nm. Se encuentra relacionada con la colágena del tipo I. VII: Forma pequeños agregados conocidos como fibrillas de fijación, que sujetan a la lámina basal los haces de fibras de colágena de los tipos I y III subyacentes. Formación de las fibras de colágena: Las cadenas polipeptídicas de la colágena se sintetizan en ribosomas unidos a membrana y se inyectan en el lumen del retículo endoplásmico en forma de grandes precursores denominados Procadenas a. Estos precursores presentan no solo el “polipéptido de señal” necesario para trasportar las proteínas segregadas a través de la membrana del retículo endoplásmico sino también otros aminonoácidos en sus extremos amino y carboxilo terminal denominados péptidos de extensión. En el lumen del retículo endoplásmico c/procadena α se combina con otras 2 formando una molécula helicoidal de 3 hebras estabilizadas por enlaces de H. Los péptidos de extensión son importantes para guiar la formación de la triple helice y pueden serlo también en el empaquetamiento de las moléculas de procolágena con otras macromoléculas de la matriz, proceso que se produce dentro de la célula antes de la secreción. Las cadenas α de colágena son extremadamente ricas en glicina y en prolina, aminoácidos que son importantes para la formación de una triple hélice estable. La glicina es el único aminoácido que es suficientemente pequeño para ocupar el abarrotado interior de la triple hélice de colágena y en la mayoría de las regiones de la cadena α constituye un residuo de c/tres. Algunos de los residuos de prolina (y de lisina) se hidroxilan en el retículo endoplásmico 10 antes que las procadenas α se asocien formando las moléculas de triple hebra de procolágena. Los grupos hidroxilo de los residuos de hidroxiprolina forman enlaces de H entre las cadenas que contribuyen a estabilizar la triple hélice (Condiciones que impiden la hidroxilación de la prolina, deficiencias de O, Fe, ácido ascorbico (Vitamina C), inhiben la formación de la hélice de procolágena. En el escorbuto enfermedad humana causada por una deficiencia en la dieta de vitamina C, las procadenas α no hidroxiladas se degradan en la célula y la piel y los vasos sanguineos se vuelven extremadamente frágiles). Tras su secreción al espacio extracelular, las moléculas de colágena se ensamblan en polímeros ordenados denominados Fibrillas de colágena que son largas estructuras (de hasta muchos µm) finas ( de 10 a 300 nm de diámetro) en forma de cuerda (visibles al Microscopio Electrónico). A menudo estas fibrillas están agrupadas en haces mayores (visibles al microscopio óptico) formando fibras de colágena de varios µm de diámetro. Al microscopio electrónico las fibrillas de colágena aisladas, una vez fijadas y contrastadas, muestran unas estriaciones transversales c/67 nm. este patrón refleja la disposición de las moléculas de colágena dentro de la fibrilla. Probablemente para aumentar al maximo la resistencia del agregado a las fuerzas de tracción (extensión), las moléculas de colágena están apiladas de forma que las moléculas adyacentes quedan desplazadas longitudinalmente casi una cuarta parte de su longitud, esta disposición produce las estriaciones en las fibrillas observadas con tinción negativa. Elásticas: Estas fibras suelen ser más delgadas, largas y ramificadas en el tejido conectivo laxo, pero pueden formar haces más densos en los ligamentos y en las láminas fenestradas. En preparados en fresco, no coloreados, las fibras elásticas se visualizan como filamentos muy finos (0.2 a 1 µm de diámetro) de gran refringencia. Las fibras elásticas frescas tienen un color amarillento solo observable cuando se encuentran en grandes cantidades o son gruesas, por ejemplo en las bandas elásticas de la columna vertebral. Las fibras elásticas pueden colorearse selectivamente con resorcina-fucsina con lo que adquieren una coloración violeta oscuro. En los ligamentos elásticos las fibras están dispuestas paralelamente y son más gruesas que en el tejido conectivo laxo, con un diámetro de 5 a 15 µm. Además la elastina se presenta también como membranas por ejemplo en las paredes arteriales. Las fibras elásticas son elaboradas por los fibroblastos y por las células musculares lisas de los vasos sanguíneos. Para poder desempeñar su función, tejidos como la piel, los vasos sanguineos y los pulmones requieren elasticidad ademas de resistencia a la tracción. Una extensa red de fibras elásticas en la matriz extracelular de estos tejidos les proporciona la capacidad necesaria de recuperarse tras una distensión transitoria. El principal componente de las fibras elásticas es la elastina, proteína de 70.000 dalton que al igual que la colágena es rica en prolina y en glicina pero que a diferencia de la colágena, contiene muy poca hidroxiprolina y nada de hidroxilisina y también contiene los aminoácidos, poco frecuentes, desmosina e isodesmosina. Estos dos aminoácidos forman un enlace cruzado considerable de las moléculas de elastina, e imparten un grado elevado de elasticidad a las fibras elásticas al grado de que estas fibras pueden estirarse hasta cerca de 150% de sus longitudes en reposo antes de romperse. Después del estiramiento vuelven a su longitud en reposo. El centro de fibras elásticas está compuesto por elastina y está rodeado por una vaina de microfibrillas. C/u de los cuales tiene un diámetro aproximado de 120 nm y está compuesta por la glucoproteína fibrilina. Durante la formación de fibras elásticas, primero se elaboran las microfibrillas y a continuación se deposita elastina en el espacio rodeado por las microfibrillas. Las moléculas de elastina se segregan al espacio extracelular donde forman filamentos y capas en los que las moléculas de elastina presentan un gran número de puentes cruzados entre ellas, dando lugar a una extensa red. 11 A diferencia de la mayoría del resto de las proteínas, la función de las moléculas de elastina requiere que su esqueleto polipeptidico quede sin plegar, en forma de los llamados “espirales al azar”. Las fibrillas de colágena entretejidas con las fibras elásticas limitan el grado de distensión y evitan asi que el tejido se desgarre. 2.-Sustancia Básica: Los GAG, anteriormente conocidos como mucopolisacáridos, son largas cadenas de polisacáridos no ramificadas, compuestas por unidades repetidas de disacáridos. Actualmente reciben el nombre de GAG debido a que uno de los 2 residuos de azucar del disacárido repetido siempre es una aminoazúcar (N-acetilglicosamina o N-acetilgalactosamina). El otro típicamente un ácido urónico (idurónico-glucurónico).Los GAG tienen una intensa carga negativa debido a la presencia en muchos de los residuos de azúcar de grupos carboxilo. Se han diferenciado 7 grupos de GAG según sus residuos y el número y localización de los grupos sulfato. Se trata del: 1. Ácido hialurónico (es el único en que ninguno de sus azícares está sulfatado). 2. Condroitín-4-sulfato 3. Condroitín-6-sulfato 4. Dermatan sulfato 5. Heparan sulfato 6. Heparina 7. Queratan sulfato El ácido hialurónico se presenta como una única cadena muy larga de carbohidrato, de varios miles de residuos de azúcar y que presenta una secuencia regular, repetida de unidades de disacárido. Métodos de tinción: Aunque los GAG sulfatados son lo suficientemente polianiónicos (ácidos) para absorber ciertas hematoxilinas en preparaciones de hematoxilina/eosina de la matriz del cartílago, su concentración en la sustancia intersticial amorfa del tejido conectivo laxo es demasiado baja como para que se tiñan eficazmente por este medio. Los métodos de tinción que pueden indicar su presencia en este tejido incluyen las tinciones catiónicas (básicas) de alta afinidad Proteoglucanos: Los PG pueden ser de diversos tamaños y varian entre unos 5000 daltons (decorina y betaglucano) hasta cerca de 3 millones de daltons (agrecan). Tienen una heterogeneidad potencial casi ilimitada. Pueden diferir notablemente en contenido proteico, en tamaño molecular, en número y tipos de cadenas de GAG por molécula. Los centros proteínicos de los PG se elaboran en el RER y los grupos GAG se enlazan de manera covalente con la proteína en el AG. Suelen contener hasta un 90% y un 95% en peso de carbohidratos en forma de numerosas cadenas de GAG largas, no ramificadas y por lo general sin ácido siálico. Ejemplo: un PG de cartílago podría consistir en unas 100 cadenas de condroitin sulfato y unas 60 cadenas de queratan sulfato unidas a un núcleo proteico rico en serina de unos 1900 aminoácidos (aproximadamente una cadena de GAG por c/12 residuos de aminoácidos). Muchos PG en especial el agrecan, molécula que se encuentra en el cartílago y en el tejido conectivo propiamente dicho, están unidos con ácido hialurónico. El modo de inserción incluye proteínas de enlace junto con la proteína central del agrecan, asi como con los grupos azúcares de los ácidos hialurónicos. Como el ácido hialurónico puede tener 20 µm de longitud, el resultado de esta asociación es un compuesto de agrecan que ocupa un volumen gigantesco (puede tener una masa molecular de varios cientos de millones de daltons). Esta inmensa molécula es la encargada de producir el estado de gel de la matriz extracelular y actúa como una barrera contra la difusión rápida 12 de los depósitos acuosos, como se pone de manifiesto cuando se observa la desaparición lenta de una burbuja acuosa después de su inyección subdérmica. Funciones de los PG: - Las cadenas de GAG tienden a adoptar conformaciones enrolladas al azar muy extendidas y a ocupar un volumen inmenso para su masa. Al ser muy hidrofílicas atraen grandes cantidades de agua, formando asi geles hidratados incluso a concentraciones muy bajas. Esta tendencia está notablemente incrementadas por su elevada densidad de cargas negativas que atraen a cationes osmoticamente activos. Esta propiedad de atraer el agua genera una presión de hinchamiento o turgencia en la matriz extracelular, que resiste a las fuerzas de compresión (a diferencia de las fibrillas de colágena que resisten a las fuerzas de tracción). - Debido a su organización porosa e hidratada, las cadenas de GAG permiten la rápida difusión de las moléculas hidrosolubles, la migración de células y el desarrollo de procesos celulares. - Llenan eficazmente el espacio extracelular, a pesar de que la cantidad de GAG del tejido conectivo es inferior al 10% en peso de la cantidad de proteínas fibrosas (colágena y elastina). - C/vez se dispone de más pruebas de que el ácido hialurónico tiene una función especial en los tejidos a través de los que migran las células durante el desarrollo o la reparación de heridas. No solo se produce en grandes cantidades en estos tejidos sino que además su degradación por medio de la enzima hialuronidasa esta asociada con el cese de la migración celular. - Los PG poseen también sitios de fijación para ciertas moléculas de señalamiento como el factor β de transformación del crecimiento (TGF-β). Al fijarse a estas moléculas de señalamiento, los PG pueden impedir su función al evitar que las moléculas lleguen a sus destinos o incrementarlas al concentrarlas en una localización específica. Glucoproteínas de Adhesión: La capacidad de las células para adherirse a los componentes de la matriz extracelular se encuentra mediada, en gran medida por las glucoproteínas de adhesión. Estas grandes macromoléculas tienen varios dominios, uno de los cuales por lo menos suele fijarse a las proteínas de la superficie celular llamadas Integrinas, uno a las fibras de colágena y uno más a los PG. De esta manera, las glucoproteínas de adhesión comprimen a los diversos componentes de los tejidos entre si. Los tipos principales de PG son: 1. Fibronectina: Gran dímero compuesto por dos subunidades polipeptídicas similares, c/u de cerca de 220000 daltons, unidas entre sí por sus extremos carboxilo mediante enlaces disulfúricos. Se sintetiza principalmente en los fibroblastos. Marca las vias migratorias de las células embrionarias, de modo que las células del organismo en desarrollo que están emigrando pueden llegar a su destino. 2. Laminina: Glucoproteína muy grande 950000 daltons compuesta por 3 cadenas polipeptídicas de gran tamaño A, B1 y B2. Las cadenas B se envuelven alrededor de las cadenas A y se mantienen en su posición por puentes disulfuro. La laminina se localiza casi estrictamente en la lámina basal. tiene sitios de fijación para heparan sulfato, colágena tipo IV, entactina y membrana celular. 3. Entactina: Se fija a la molécula de laminina en el sitio en que se unen entre si los 3 brazos cortos de esa molécula, también lo hace a la colágena de tipo IV y por tanto facilita la fijación de la laminina a la red de colágena. 4. Tenascina: Es una gran glucoproteína compuesta por 6 cadenas polipeptídicas que se conservan unidas entre si por enlaces disulfúricos. Tiene sitios de fijación para los sindecanes y fibronectina. Se ubica en el tejido embrionario. 5. Condronectina y Osteonectina: son semejantes a la fibronectina. La primera tiene sitios de fijación para colágena tipo II, condroitin sulfatos, ácido hialurónico e integrinas de condroblastos y condrocitos. 13 La segunda posee dominios para colágena tipo I, PG e integrinas de osteoblastos y osteocitos. Facilita la fijación de cristales calcicos de hidroxiapatita a la colágena de tipo I en el hueso. MEMBRANA BASAL La interfase entre el epitelio y el tejido conectivo está ocupada por una región acelular estrecha, llamada membrana basal, que se tiñe bien con la reacción de PAS y con otras tinciones histológicas que identifican a los GAG. Hay una estructura similar a la membrana basal, la Lámina Externa que rodea a las células musculares lisas y esqueléticas, a los adipocitos y a las células de Schwann. La membrana basal se define mejor mediante Microscopia electrónica en dos constituyentes: a.- Lámina Basal elaborada por las células epiteliales. b.- La Lámina reticular elaboradas por células del tejido conectivo. a.- Lámina Basal: La Microscopia electrónica de la lámina basal ponen de manifiesto sus dos regiones: a.1. La lámina lúcida: región electrolúcida de 50 nm de espesor justamente por debajo del epitelio. Consiste principalmente en las glucoproteínas extracelulares laminina y entactina. a.2. La lámina densa región electrodensa de 50 nm de espesor. Consiste en una malla de colágena de tipo IV, que está cubierta en los lados por el PG Perlacano. La laminina tiene dominios que se fijan a la colágena del tipo IV, al heparan-sulfato y a las integrinas de la membrana celular epitelial, por lo que fijan a la célula epitelial contra la lámina basal. La lámina basal parece estar bien fija a la lámina reticular por diversas sustancias entre ellas Fibronectina, fibrillas de fijación (colágena del tipo VII) y microfibrillas todas elaboradas por los fibroblastos del tejido conectivo. La lámina basal funciona como filtro molecular a la vez que como un sostén flexible firme para el epitelio suprayacente. El aspecto de filtración se debe a la colágena de tipo IV cuya malla entretejida forma un filtro físico con tamaños de poros específicos, sino también a las cargas negativas de su constituyente heparansulfato que restringe el paso de moléculas de carga negativa. Una función adicional de la lámina basal es dirigir la migración de las células a lo largo de su superficie como sucede en la reepitelización durante la reparación de las heridas o en el restablecimiento de las uniones neuromusculares durante la regeneración de los nervios motores. b.- Lámina reticular: Los fibroblastos la elaboran, es de espesor variable, está compuesta por colágenas de los tipos I y III. Se encuentra en interfase entre la lámina basal y el tejido conectivo subyacente, y su espesor varía según el grado de las fuerzas de fricción que recibe el epitelio suprayacente. Es muy gruesa en la piel y muy delgada por debajo de la túnica epitelial del alvéolo pulmonar. Las fibras de colágena de los tipos I y III del tejido conectivo hacen asas dentro de la lámina reticular, sitio en el que entran en interacción con las microfibrillas y las fibrillas de fijación de la lámina reticular y se fijan a ellas. Más aún, los grupos básicos de las fibras de colágenas forman enlaces con los grupos ácidos de los GAG de la lámina densa. Los dominios de fijación de colágena y de GAG de la fibronectina ayudan más aún a la fijación de la lámina basal a la lámina reticular. La cubierta epitelial esta fija al tejido conectivo subyacente por medio de estas interfases acelulares resistentes, la lámina basal y la reticular. 14 INTEGRINAS Son proteínas transmembranales semejantes a los receptores de la membrana celular porque forman enlaces con ligandos. A diferencia de los receptores sus regiones citoplasmáticas se encuentran fijas al citoesqueleto y sus ligandos no son moléculas de señalamiento sino miembros estructurales de la matriz extracelular como colágena, laminina y fibronectina. Las integrinas son mucho más numerosas que los receptores. Las integrinas son heterodímeros (de unos 250000 daltons) compuestos por cadenas glicoproteínicas α y β cuyos extremos carboxilo están enlazados con la Talina y Actinina α del citoesqueleto. Sus extremos amino poseen sitios de fijación para macromoléculas de la matriz eztracelular. Como las integrinas enlazan al citoesqueleto con la matriz extracelular se conoce también como Enlazadores Transmembranales. La cadena α de la molécula de integrina fija Ca+2 o Mg+2, cationes divalentes necesarios para la conservación de la fijación apropiada con el ligando. Son numerosas las integrinas que difieren en su especificidad por ligandos, su distribución celular y su función. VARIEDADES DEL TEJIDO CONECTIVO Tejido conectivo Laxo: También denominado areolar, es el más común. Rellena los espacios entre las fibras y haces musculares, sirve de apoyo a los epitelios. Forma una capa alrededor de los vasos sanguíneos y linfáticos. Las células más abundantes son los fibroblastos y los macrófagos. Las fibras elásticas, colágenas y reticulares están presentes. El tejido conectivo laxo es de consistencia delicada, flexible y poco resistente a las tracciones. Tejido conectivo Denso: Predominan las fibras colágenas. Las células son menos numerosas que en el laxo y entre ellas sobresalen los fibroblastos. Es menos flexible que el laxo y mucho más resistente a las tracciones. Cuando las fibras colágenas se disponen en haces sin orientación fija tenemos el tejido denso no modelado que se encuentra por ejemplo en la dermis profunda de la piel. `Las fibras colágenas se orientan según una organización fija se trata del tejido conectivo denso modelado por ejemplo los tendones. Los tendones son estructuras cilíndricas alargadas que unen los músculos esqueléticos a los huesos. Debido a su riqueza en fibras colágenas son blancos e inextensibles. Están formados por haces paralelos de fibras colágenas, entre las cuales existe escasa cantidad de sustancia intercelular amorfa y de fibrocitos con núcleos alargados paralelos a las fibras y citoplasma delgado que tiende a envolver los haces colágenos. Los haces colágenos del tendón (haces primarios) forman conjuntos (haces secundarios) envueltos por tejido conectivo laxo, que contiene vasos y nervios. El tendón está envuelto externamente por una vaina de tejido conectivo denso. En algunos tendones esta vaina está dividida en dos capas, una sujeta al tendón y otra unida a las estructuras vecinas. Se forma una cavidad revestida por mesotelio, en la que hay un líquido que contiene agua, musopolisacáridos, proteínas e iones que facilita el deslizamiento del tendón. 15 Tejido conectivo Elástico: Está formado por fibras elásticas gruesas paralelas y organizadas en haces separados por tejido conectivo laxo. Es poco frecuente, se lo encuentra por ejemplo en los ligamentos amarillos de la columna vertebral. Tejido conectivo Reticular: Constituído por fibras reticulares en íntima asociación con las células reticulares primitivas. Se lo halla en los órganos hemocitopoyéticos constituyendo el armazón que soporta las células libres existentes. Las células reticulares primitivas poseen largas prolongaciones que se unen a la de células vecinas. Sus núcleos son grandes. Tejido conectivo Mucoso: Predomina la sustancia intercelular amorfa. Contiene fibras colágenas y raras elásticas y reticulares. Las células son principalmente fibroblastos. Este tejido es el principal componente del cordón umbilical (gelatina de Wharton). BIBLIOGRAFÍA: 1. Alberts B., Bray D., Lewis J., Raff M., Roberts K., Watson J. Biología molecular de la Célula. Ed. Omega S.A. Barcelona. España. 1983 2. Banks W. Histología Veterinaria Aplicada. Editorial el manual moderno. México. 1986. 3. Bloom W., Fawcett D.W.Tratado de Histología. Ed. Labor S.A. España. 8 Ed. 1981. 4. Cormack D. H. Histología de Ham. Ed. Harla. México. 9na. Ed. 1987. 5. Dellmann H. Textbook of Veterinary Histology. Ed. Lea y Febiger. Philadelphia. 4ta. Ed. 1993. 6. Dellmann H.D., Carithers J.R. Citología e Histología. Serie Nacional de Medicina Veterinaria. Ed. Inter- Médica S.A. I. C. I. Bs. As. Argentina. 1999. 7. Gartner L.P., Hiatt J.L. Histología, Texto y Atlas. Ed. McGraw-Hill Interamericana Editores S. A. de C.V. Mexico.1997. 8. Geneser F. Histología. Ed. Médica Panamericana S. A. De C. V. Mexico. 1ra. Ed. 9na. Reimpresión. 1992. 9. Hib J. Histología de Difiore. Texto y Atlas. Ed. El Ateneo Bs. As. 1ra. Ed. 2001. 10. Junqueira L. C., Carneiro. Histología básica. Ed. SALVAT S.A. Barcelona. España. 1ra. Ed. 1988. 11. Labat-Robert J., Bihari-Varga M., Robert L. Extracellular matrix. FEBS 268 (2): 386-393. 1990. 12. Leblond C. P., Inove S. Structure, Composition and Assembly of Basement Membrane. The American Journal of Anatomy 185: 367-390. 1989. 13. Leeson T. S., Leeson R., Paparo A. Texto y Atlas de Histología. Ed. Interamericana. - Mc Graw-Hill S. A. Mexico. 1ra. Ed. 1990. 14. Mecham R. P., Heuser J. E. The Elastic Fiber. Cell Biology of Extracellular Matrix. Second Edition. Edited by E. Hay Plenum Press. New York. Chapter 3 pp 79-109. 1991. 16 15. Pecci Saavedra J., Vilar O., Pellegrino de Iraldi A Histología Médica. Ed. López Libreros Editores S.R.L. bs. As. Arg. 1ra. Ed. 1984. 16. Ross M., Reith E., Romrell L. Histología. Texto y Atlas Color. Ed. Médica Panamericana S.A. Bs. As. 2da. Ed. 1992. 17. Schwartz N. B., Smalheiser N. R. Biosynthesis of Glycosaminoglycans and Proteoglycans. Neurobiology of Glycoconjugates. Edited by R. V. Margolis and R. K. Margolis. Plenum Publishing Corporation. Chapter 5. Pp 151-186. 1989. 18. Trautmann A.Histología y anatomía microcópica comparada de los Animales domésticos. Ed. Labor S.A. Madrid. 1ra. Ed. 1950. 19. Von Lawzewitsch I. Lecciones de Histología Veterinaria. 9 tomos. Ed. Hemisferio Sur. Bs. As. 1987