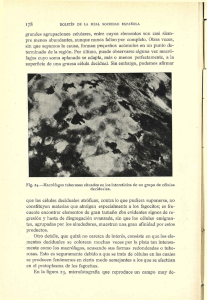
LOS FAGOCITOS Y SUS RECEPTORES Tipos de fagocitos
Anuncio

LOS FAGOCITOS Y SUS RECEPTORES Tipos de fagocitos: Los fagocitos pueden ser de dos tipos: monocitos y polimorfonucleares neutrófilos (granulocitos. - Los monocitos nacen en la médula ósea y pasan a la sangre, donde su tiempo de vida es breve (1-3 días), excepto si abandonan el torrente circulatorio hacia los órganos y los tejidos, donde se convierten en macrófagos (viven mucho tiempo). En condiciones normales la mayoría de los monocitos no son necesarios y mueren por apoptosis, solo unos pocos se convierten en macrófagos. Estas células reciben nombre específicos: células de Kupffer (en el hígado), células de la microglía (en el SNC), células mesangiales (en el riñón), osteoclastos (en el hueso) o macrófagos alveolares (en los pulmones). Los monocitos, cuando abandonan la sangre, sufren procesos de maduración adquieriendo un aparato de Golgi prominente y numerosos lisosomas, denominándose así magrófagos. En conjunto, los macrófagos forman una red denominada sistema mononuclear fagocítico. Se localiza en zonas por donde pudieran entrar microbios o se filtre sangre o la linfa. - Los polimorfonucleares o granulocitos (tienen núcleos con varios lóbulos y números gránulos en su citoplasma) pasan de la médula ósea al torrente circulatorio donde tienen una vida media muy corta. Hay tres tipos: -Neutrófilos: son los más abundantes, y su principal función es fagocítica. -Basófilos: cuya principal función es la de realizar reacciones inflamatorias de emergencia. -Eosinófilos: se emplea frente a los parasitos que por su tamaño no pueden ser fagocitados. Lleva a cabo la liberación del contenido de sus gránulos al exterior (exocitosis). Los polimorfonucleares (a diferencia de los monocitos) no abandonan el torrente circulatorio a no ser que se produzca una inflamación. En estos lugares se liberan moléculas quimiotácticas que inducen la extravasación de los fagocitos a los tejidos. En estado de reposo, los fagocitos tienen una actividad limitada que se potencializa (se activan) si interaccionan con una serie de citocinas entre la que se destaca el Interferón gamma. Otras moléculas que pueden activar los fagocitos son la endotoxina o lipopolisacárido bacteriano (LPS). Funciones de los fagocitos La principal función es la fagocitosis. Para ellos, debe reconocer el patógeno a fagocitar, y en este reconocimiento de microorganismos existen diferencias entre las células de la inmunidad innata (fagocitos) y la adquirida. Las células de la inmunidad innata tienen los mismos receptores, por lo que esta inmunidad reconoce moléculas o patrones generales, comunes a muchos patógenos (son los PMAP). En cambio, la inmunidad adquirida reconoce moléculas específicas del patógeno (como el LPS). Los fagocitos permiten una rápida y eficaz eliminación de microorganismos con los que nunca hemos estado en contacto previamente. Si el patógeno no es reconocidos por los fagocitos o consigue sobrepasarlos, se produce un foco infeccioso. Será en este momento cuan entraría en juego la respuesta adquirida (más lenta). Cuando el microorganismo es reconocido por los receptores de los fagocitos, se produce su internalización dentro de una vacuola (fagosoma) y su fusión con los lisosomas (fagolisosoma) donde se destruirán los patógenos. Entre los receptores mejor caracterzados están el receptor de la manosa (reconoce carbohidratos con gran número de manosas) y el receptor de detritos o carroñero (une y media la fagocitosis de LDL oxidadas o acetiladas). La fagocitosis se potencia mediante la opsonización (o preparación del patógeno) del patógeno, mediante el recubrimiento de éste mediante complemento o anticuerpos. Si es mediante anticuerpos, los receptores de la superficie del fagocito reconocen la porción Fc del anticuerpo (la IgG se unen a los receptores Fc gamma RI, RII y RIII). La activación del complemento en la superficie de los patógenos, en especial la opsonina C3b, que es reconocida por los fagocitos a través de los receptores del complemento CR1, CR3 y CR4. La activación del complemento por la vía clásica o de las lectinas sobre la superficie de un patógenos, hace que sea reconocido por los fagocitos a través del receptor de C1q. Los fagocitos también eliminan células propias deterioradas, envejecidas o muertas, con el objetivo de mantener la homeostasis del organismo. Dentro de la fagocitosis, hay que hablar del proceso de la propia fagocitosis y de la lisis de microorganismo: La fagocitosis es llevada a cabo principalmente por los monocitos/macrófagos y por los neutrófilos reclutados al lugar de inflamación. Una vez reconocido al patógenos como tal mediante los receptores, será fagocitado. La fagocitosis se producirá mediante pseudópodos que rodean a la partícula y la introducen en el citoplasma en el interior de una vacuola (fagosoma). Posteriormente, a los fagosomas se le unen los lisosomas (acidificará el ambiente) formando fagolisosomas, donde las diferentes hidrolasas ácidas llevarán a cabo la lisis bacetriana. Al activarse los fagocitos, se incrementa la captación de oxígeno que es utilizado para generar productos intermediarios reactivos de oxígeno, capaces de destruir a microorganismos. Es fenómeno de generación de radicales libres de oxígeno se conoce como explosión oxidativa. Forma moléculas muy reactivas que reaccionara con las moléculas del microbio y lo destruirán, moléculas como el NADOH oxidasa (reduce el oxígeno molecular a superóxido) o la mieloperoxidasa (utiliza el peróxido de hidrógeno para la halogenación de los microoganismo). Además de la explosión oxidativa, los macrófaos cuenta con un segundo sistema de generación de radicales libres: la sintasa del óxido nítrico inducible (NOS2) que genera óxido nítrico (NO, también un potente microbicida). La actividad inmunomoduladora En la superficie de los fagocitos están presentes los receptores de señalización (tipo Toll o TLR) que al reconocer moléculas propias de los patógenos activan los fagocitos. Un ejemplo es el TLR4 que reconoce el LPS bacteriano. Al activarse, los fagocitos inducen la secreción de una serie de mediadores de la inflamación. La inflamación se produce sobre todo por los macrófagos que secretan citocinas. Estos mediadores junto con la activación del complemento, las cininas y la degranulación de los mastocitos tisulares son responsables mayoritariamente del incio de la respuesta inflamatoria. La actividad pro y antiinflamatoria Las citocinas que tienen mayor impostancia son el TNF alfa y las quimiocinas. El TNF alfa aumenta el diámetro de los vasos sanguíneos próximos al lugar de inflamación y la permeabilidad vascular. Las quimiocinas reclutan leucocitos que provienen de la sangre al lugar de la inflamación: los neutrófilos inician la destrucción bacteriana y tisular y luego los monocitos convertidos en macrófagos termian de eliminar las bacterias e inician la reparación tisular. Además, en la fase de la inmunidad adquierida, los macrófagos pueden diferenciarse en células dendríticas, que son presentadoras de antígeno y que migran a los órganos linfoides secundarios donde desarollarán la respuesta inmunitaria específica y crean linfocitos T y B de memoria. Los macrófagos pueden estar en dos estados de activación: - - La activación clásica o M1: los macrófagos tienen una actividad proinflamatoria eliminando los microorganismos restantes y loas células muertas por apoptosis. La activación alternativa o M2: los macgrófagos tienen una actividad de reparación o reconstrucción denominada antiinflamatoria. El balance entre ambas actividades (pro-anti) es crítico, es decir, es necesario un equilibrio entre ambas. Los linfocitos T de tipo Th1 pueden activar los macrófagos mediante interferón gamma. Los macrófagos así activados les permite la eliminación de patógenos intracelulares. La activación de la inmunidad adaptativa Las células de la respuesta adaptativa sólo responden a los patógenos una vez han sido reconocidos por el sistema innato. En los linfocitos, se producen receptores para todos los antígenos posibles, incluso los antígenos propios. Debido a que los fagocitos tienen receptores específicos para los patógenos (son los receptores de tipo Toll y otros los que lo reconocen primariamente), estos son los que controlan que la activación de los linfocitos se produzca exclusivamente contra los patógenos, y no contra antígenos propios. Después de reconocer al antígeno, las células presentadoras de antígeno (APC), que son los macrófagos y las células dendríticas, dirigirán la respuesta adaptativa frente a esos microbios mediante la presentación de antígeno. El repertorio de receptores de la inmunología innata es más reducido e inespecífico que el sistema de los receptores B, Ac y receptores T de la inmunología adaptativa