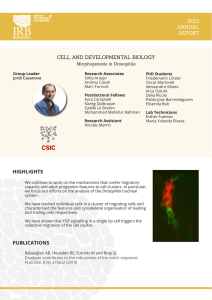
y CIENCIAS - Pontificia Universidad Católica del Ecuador
Anuncio

Revista Ecuatoriana de MEDICINA y CIENCIAS , BIOLOGICAS VOLUMEN XXVII NÚMEROS 1 Y 2 PÁGINAS 1 - 79 OCTUBRE /2005 D D Revista Ecuatoriana de MEDICINA y CIENCIAS BIOLÓGICAS VOLUMEN XXVII - NÚMEROS 3 Y 4 AGOSTO /2005 © Casa de la Cultura Ecuatoriana Cl "Benjamín Carrión", 2005 © Fondo Editorial C.C.E. 2005 Av. 6 de Diciembre N 16-224 YAv. Patria Impresión, Encuadernación: Editorial Pedro Jorge Vera Diseño gráfico: César E. Salazar O. ISSN: 00349313 Impreso en Ecuador - Printed in Ecuador E-mail: [email protected] www.cce.org.ec J DIRECTORIO Casa de la Cultura Presidente: Dr. Marco Antonio Rodríguez Secretario General: Dr. José Regato Sección de Ciencias Naturales, Biológicas y Ciencias Exactas Presidente: Secretario: Vocales Dr. Claudio Cañizares Proaño Dr. Oswaldo Morán Pinto Dr. Plutarco Naranjo Vargas Dra. Laura Arcos Terán Dr. david Niell Miembros: Guillenno Álvarez Domínguez Laura Arcos Terán Fernando Astudillo Arroyo Celín Astudillo Espinosa Enriqueta Banda de Naranjo Jaime Bhreil Augusto Bonilla Barco Claudio Cañizares Proaño Eugenia Del Pino Jaime Díaz Moreno Rodrigo Fierro Benítez Raúl Granda Alex Hirtz Eduardo Luna yépez Anlbal J arrín Oswaldo Morán Pinto Marcelo Moreano Dávila Miguel Moreno Espinoza Plutarco Naranjo Vargas Holm Nielsen Lauritz DavidNeil Arturo Nina León Milton Paz y Miño Hernán Proaño Edgar Samaniego Rojas Cristóbal Santacruz Fausto Sanniento Rodrlguez Jorge Suárez Annijos Fernando Sempértegui Ontaneda Jorge Soria José Varea Terán Henrik Vaslev Rodrigo Yépez Miño Comité Editorial: Comité Externo: Dr. Claudio Cañizares P. Dr. Oswaldo Morán P. Dr. Plutarco Naranjo V. Dra. Violeta Rafael Dra. Katya Romoleroux Dra. Eugenia del Pino D ISSN 0034 9313 Revista Ecuatoriana de Medicina y Ciencias Biológicas - Volumen XXVII - Números 1 y 2 Páginas 1 - 79 - Octubre 2005 , INDICE Directorio ................................................................Pág. 3 Editorial .................................................................Pág. 5 TRABAJOS ORIGINALES Importancia del estudio de la hemostasia y coagulación en las enfermadades tromboembólicas. Cañizares, C................................................ Pág. 7 Desarrollo neural de la rana Colostethus machalilla (ANURA: DENDROBATIDAE). Noboa, V. y del Pino, E. . ...................................................Pág. 15 Cario tipos Metafásicos de Drosophila inca y D. yangana, subgrupo inca, grupo repleta. Mafia, A.B...................................................Pág. 21 Catorce nuevas' especies del género Drosophila (Diptera, Drosophilidae) en el Bosque húmedo montano del Volcán Pasochoa, Pichincha, Ecuador. Vela, D. y Rafael, V. .......................................................Pág.27 Identificación de islas CpG en el genoma humano a través de las cadenas de Markov: Un modelo matemático basado en probabilidades. Quishpe, E.; Sánchez, M.E.; Oleas, G. y Paz - y - Miño, C. .........................Pág. 43 Elementos Alu en el análisis filogenético molecular del infraorden Platyrrhini, Primates. Ludeña, B.; Escobar, J.C. y Pintaud, J-C ................................ Pág. 47 e TRABAJOS DE REVISI6N La originalidad del desarrollo de la anestesia en el Ecuador.- Siglo XX - 1960 - 2000. Morán, O .................................................................Pág. 53 Bioética y práctica médica. Chaves, O......................................... Pág. 57 El denominador "nacidos vivos" en investigación de salud pública. Albán, E.; Hermida, c.; Hennida, J.; Vásconez, J.; Vergara, M.; Hinojosa, M.; Machuca, M. y Oviedo, N ....................................................Pág. 61 "Deshumanización de la medicina y bioética". Álvarez, G ........................ Pág. 67 HOJAS DE VIDA Doctor Plutarco Naranjo ....................................................Pág.71 Instructivo para los autores ................................................Pág. 75 D EDITORIAL ....................................-.--............ ..----------if---------------------------------------------------------------------­ L a comunidad científica está de plácemes. El Presidente Dr. Alfredo Palacio logró establecer un 5% de los fondos del FEIREP para ciencia y tecnología, pudo haber sido más pero el Congreso Nacional no captó la importancia de la inversión en investigación científica, ya que sus intereses son diferentes. No son conscientes de que el componente de ciencia y tecnología es más importante que el financiero, banca­ rio, industrial, comercial para el desarrollo de un país. Las sociedades que aplican este concepto logran in­ menso desarrollo, a pesar de que algunos de ellos son países de territorio pequeño. Singapur, la ciudad na­ ción, formó en su país y en el extranjero en una década 20.000 doctores en las diferentes ramas de la cien­ cia, igual que Corea del Sur, Hong Kong, para no mencionar otros más grandes. De todas maneras lo logrado es plausible, nos llena de esperanza. Anteriormente FUNDACYT sola­ mente recibía el 0.25% del monto de los contratos de sector público que apenas significaba 1 millón 200 mil dólares anuales. Con ese presupuesto las acciones para el desarrollo científico del país eran muy redu­ cidas. Durante los años 2.000 - 2002 se pudo aplicar un préstamos del BID para ciencia y tecnología por 20 millones de dólares con una contrapartida nacional de 10 millones. Esto se logró durante la vicepresiden­ cia del Economista Alberto Dahik. No se pudo lograr otro préstamo por que el Ecuador no cumplió con la contrapartida. El efecto de esa inversión fue muy fructífero. 180 doctorados en el exterior y algunas maes­ trías. Instalación de varios centros de investigación en las universidades. 35 proyectos de investigación uni­ versitaria, 6 proyectos de innovación del sector privado. La formación de la Red Ecuatoriana de Ciencia y tecnología REICYT a la cual están inscritas 35 instituciones nacionales. Infortunadamente después del 2002 estas acciones volvieron a decaer y se redujeron notablemente por que solamente se contó con las reducidas asignaciones de FUNDACYT. Ahora el horizonte del desarrollo científico y tecnológico se visualizan prometedoras por la impor­ tante asignación de fondos. Debemos congratularnos por eso D Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vol. XXVII Números 1 y 2: octubre de 2005 D D TRABAJOS ORIGINALES ----------------------------------------lf---­ Importancia del estudio de la hemostasia y coagulación en las enfermadades tromboembólicas Claudio Cañizares Proaño Servicio de Hematología y Banco de Sangre Hospital Carlos Andrade Marín IESS Recibido 3 de marzo 2005, aceptado 17 marzo 2005. RESUMEN. La prevalencia de la enfermedad tromboembólica es alta en nuestro medio. Existen factores de riesgo como la edad, obesidad. hipertensión, diabetes, sedentarismo, anticonceptivos, sepsis, cáncer, tabaquismo, dietas ricas en grasas, venas varicosas. La causa desencadenante es la arteroesclerosis, flebitis que producen la formación de trombos in situ asociados a la hipercoagula­ bilidad, inflamación y desórdenes del metabolismo de los lípidos, así como también la migración de émbolos . Las manifestaciones clínicas dependen del órgano afectado pero el tratamiento es el mismo, que consiste en la terapia anticoagu­ lante. Nuestros resultados demuestran la importancia de estudiar los parámetros de hipercoagulabilidad. Encontramos que el 83% de los casos son por hipercoagulabilidad, sóla o asociada a otras causas como por inflamación en el 45.5% y por hiperlipidemia en el 45.7%. PAI.ABRAS CLAVES: Arterioesclerosis, Hipercoagulabilidad. ABSTRACT. The frequency of the thromboembolic disease is high in our populatíon. Several risk factors have been identified as favoring the development of thrombosis, such as: age, obesity, hypertension, diabetes, immobilization, estro gen therapy, sep­ sis, cancer, smoking, lipid diets, varicose veins. The ultimate cause of these events is arteriosclerosis in the arteries, phlebitis in the veins, that produce the formation or migra­ tion of clots in the circulation due to hypercoagulation , inflammation and lipid metabolic disorders. Even though the clínical features depend on the affected organ the treatment ís the same, consisting in anticoagu1ant therapy. Our results place emphasis in the importance on studdíng the parameters of hypercoagulation. We found Hypercoagulation in 83% of the cases, along or combine with inflammation in 45.5% or lipid metabolic disorders in 45.7%. Further data is stated in the paper. KEYWORDS. Artherioesclerosis, Hypercoagulation. INTRODUCCIÓN La enfermedades tromboembólicas son una patología común en la práctica médica, en nuestro país están como la segunda causa de morbilidad. Los episodios afectan diferentes órganos y siste­ mas, como infarto cerebral, infarto de miocardio, infartos mesentéricos, trombosis arteriales periféri­ cas, trombosis venosas profundas y superficiales, tromboembolias pulmonares, trombosis retinianas arteriales y venosas y otros. La sintomatología de estos problemas depen­ de del órgano afectado pero la etiología es la misma y se debe a la formación de trombos en el sitio afec­ tado o a la migración del trombo a distancia por el desprendimiento del mismo, que se conoce como émbolo. El trombo se forma por el despulimiento de la pared vascular ya sea por acción metabólica y de flujo circulatorio o por inflamación por proceso infeccioso (1,2) Las causas predisponentes ( 3,4,5) son: Hipertensión arterial Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vol. XXVII Números 1 y 2: 7-14, octubre del 2005 Cañizares 8 Diabetes Obesidad Sedentarismo Tabaquismo Dietas grasas Cáncer Várises e insuficiencia venosa circulatoria especialmente en miembros inferiores Constitucionales: edad, raza, embarazo, me­ nopausia, anticonceptivos. Familiar: predisposición a enfermedad trombótica por homocistinuria, hiperviscosidad sanguínea. Las causas desencadenantes (6,7) son: Arterioesclerosis, para los casos arteriales Tromboflebitis, Infección local, para los ca­ sos venosos Defectos prohemostáticos e hipercoagulabi­ lidad La arterioesclerosis es un proceso patológi­ co que consiste en la formación de placas fibróticas en las paredes arteriales. Formación de la Placa de Ateroma o Este proceso es lento y consiste en que en los lugares de tensión circulatoria permanente de roza­ miento (shear stress), especialmente en sitios de di­ visión de flujo y con alta concentración molecular de lipoproteínas de baja densidad (LDL) y fibrinó­ geno, se forman ovillos membranosos entre las cé­ lulas endoteliales que recubren las arterias y la membrana basal del endotelio, asociadas a partícu­ las de lipoproteínas, que se conocen con el nombre de "material semejante a los liposomas". Estas for­ maciones presiden la adherencia de los leucocitos mononucleares, tal parece que este material actúa como activador de los receptores de membrana de los leucocitos que son glicoproteínas (8). Los leucocitos mononucleares penetran , probablemente por las fenestraciones endoteliales o por las uniones endoteliales laxas, y se localizan en el espacio subendotelial, donde los monocitos son activados por las Metaloproteínas (familia de enzi­ mas especializadas dependientes del Zinc) y las moléculas de adhesividad del endotelio como la Vascular Adhesive Molecule (VCAM-l), la Mo­ nocyte Colony Stimulating Factor (M-CS), se con­ vierten en macrófagos o células rapaces (scavan­ gers). Estas células pueden unir e interiorizar las li­ poproteínas oxidadas y toman forma de células es­ pumosas . Las LDL oxidadas siguen dos caminos o compartimentos en estas células; uno es regulado por la Acilcoenzima A, Colesterol Aciltransferasa (ACAT) que interioriza estas moléculas en la célula y la otra está regulada por la Adenosintrifosfato Binding Cassette Protein (ABC-l) que por el con­ trario libera estas LDL oxidasas y las transporta fuera de la célula, propiciando su aclaración al com­ binarse con las HDL. Este es un mecanismo favora­ ble. Las células espumosas pueden llevar a la esti­ mulación genética que produce citoquinas inflama­ torias, como la Interleukine-l (IL 1), Tumor Necro­ sis Factor (TNF) , Transforming Growth Factor? (TGF?) Y moléculas reguladoras del crecimiento celular como Platelet Derived Growth Factor (PDGF), Insulin like Growing Factor (ILGF), He­ parin Binding Endotelial Growing Factor (HBEGF) y otros que estimulan el crecimiento de las células musculares (9,10,11). Las células subyacentes del músculo liso se estimulan con los factores de crecimiento prolife­ rando y migrando de la capa media a la íntima. Ade­ más el TGF? puede estimular la formación de tejido conectivo en la íntima, como proteoglican, coláge­ no, proteínas fibrosas elásticas, dando finalmente u­ na respuesta proliferativa fibrosa del músculo liso, característica que se observa a largo plazo. Formación del coágulo oclusivo: El origen de este cuadro es la ruptura de la placa de ateroma y puede darse en cualquier territo­ rio orgánico. Cuando se produce la ruptura se pone en exposición el colágeno, especialmente de Tipo I y III que induce la agregación plaquetaria , con la formación de grumos plaquetarios, actuando los agonistas de la agregación plaquetaria como el ADp, el Tromboxano A2, que también es vasocons­ trictor. Se activan las glicoproteínas receptoras de adhesividad de la membrana plaquetaria especial­ mente la GPIIbIlIla, que se une al fibrinógeno, la G­ PIb que se une fundamentalmente al Factor von Wi­ llebrand del subendotelio. Así se forman los grumos plaquetarios adheridos a la placa y se desencadena la activación de los factores de contacto de la coa­ gulación (XII, XI) Yse desencadena la cascada de la coagulación a través de la y moléculas reguladoras del crecimiento celular la Vía Intrínseca, en la que están involucrados los Factores Antihemofílicos (VIII, IX). También el desprendimiento de substan- Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vol. XXVII Números 1 y 2: 8-14, octubre del 2005 D Importancia del estudio de la hemostasia y coagulación en las enfermedades tromboembólicas lATE R O G E N E SI FIGURA 1 9 SJ ............... . .n........ _IlaIll. .I...l. ...... "'MeY" '"O."'tlLlAL .a...... ............... D".'CALa ........ • L .......,., HDl ....~.... " ...IOY......" . 11 ..., , ... e • Con la lesión endotelial se activan los monocitos, linfocitos T y plaquetas. Los monocitos se convierten en células rapaces (scavengers) que intemalizan las lipoproteínas oxidadas formando las células espumosas que activan las citoquinas estimuladoras del crecimiento del músculo liso y la colágena formando tejido fibroso que constituye la placa de ateroma. cias tromboplásticas del endotelio lesionado, Factor Tisular, y de las plaquetas substancias fosfolípidi­ cas, Factor 3 plaquetario que estimulan concomi­ tantemente también la Vía Extrínseca de la coagula­ ción en que actúan fundamentalmente los factores protrombínicos (X, VII, ll), así se desencadena a través de la Vía Común de la coagulación, donde actúa el Factor X, la formación de la fibrina y la consolidación del coágulo trombótico (12,13) CJ MATERIAL Y MÉTODOS Se estudia 35 pacientes con cuadros de trom­ boembolia: 20 con tromboembolia pulmonar 5 con trombosis venosa profunda en miembros inferiores 2 trombosis mesentérica 2 trombosis de fístula arteriovenosa, en diálisis 2 infarto de miocardio 4 infarto isquémico cerebral A los pacientes que vienen sin tratamiento antícoagulante se les estudia de inmediato la hiper­ coagulablidad y a los que ya han recibido trata­ miento anticoagulante se espera que terminen dicho tratamiento preventivo para suspender el anticoagu­ lante o aspirina por 15 días como mínimo para ha­ cerles el estudio de hipercoagulabilidad. El estudio de hipercoaiulabilidad consiste de los siguientes parámetros: Pruebas para estudiar la tendencia procoagu­ lante fisiológicos Generación de trombina Tiempo de protrombina Tiempo de tromboplastina parcial Tolerancia a la heparina in vitro Para estudiar deficiencia de anticoagulantes fisiológicos: Factor V Leiden Proteína C Antitrombina III Anticuerpos antifosfolipídicos Anticoagulante lúpico Para estudiar la hiperfunción plaquetaria: Revista Ecuatoriana de Medicina y Ciencias Biológicas· Vol. XXVII Números 1 y 2: 9-14, octubre del2005 D .... _-~ .. ~ ... _ - - - .... _~~~_ .... ~_ ... ~~ ._~--. --------------Cañizares 10 FIGURA 2 COAGULACrON PARED VASCULAR ".,.,. I'MF ........u. . El endotelio normal tiene una acción antiagregante plaquetaria a través del óxido nítrico y la prostaciclina que secreta. En caso de lesión vascular el endotelio cambia a carga + y con el desprendimiento de fibras de colágena activa las plaquetas. Por otro lado las plaquetas también tienen relación con el proceso de la coagulación, se activan con la formación de trombina y otros ago­ nistas como ADP y al adherirse al Factor von Willebrand y a la fibrina forman grumos quedando englobados en la red de fibrina, que es el coágulo. Recuento de plaquetas. Agregación plaquetaria Adhesividad plaquetaria Para estudiar la fibrinolisis: Euglobulinas, Dosificación de plasminógeno Alfa 2 antitripsina Para estudiar inflamación: Proteína e Reactiva Fibrinógeno Para estudiar alteración metabólica: Perfillipídico RESULTADOS Se estudiaron 35 pacientes y los resultados se presentan en las tablas 1,2,3. Se considera hipercoagulabilidad global. es cuando la curva de generación de trombina está Revista Ecuatoriana de Medicina y Ciencias Biológicas Vol. XXVII Números 1 y 2: 10-14, octubre del 2005 D Importancia del estudio de la hemostasia y coagulación en las enfennedades tromboembólicas 1. Estudio de hipercoagulabilidad Hipercoagulabilidad global por exceso de procoagulantes Antifosfolipídico 5 (14.6%) Factor II 3 ( 8.6) Factor VIII 2 (5.8) II 10 (29%) Hipercoagulabilidad global por deficiencia de anticoagulantes fisiológicos Proteína e 2 (5.8) Factor V Leiden 1 (2.7) 6 (17) Hipercoagulabilidad final por deficiencia de fibrinolisis Plasminógeno 4 (11.4%) aantitripsina 2 (5.8) no identificada 2 (5.8) 8 (23) Hipercoagulabilidad inicial asociada a hiperfunción plaquetaria Trombocitemia 3 (8.6%) Hípera gregación 2 (5.8) 5 (14) No Hipercoagulabilídad 6 (17) Tabla l. La hipercoagulabílidad global es la más frecuente 2. Alteración asociada a inflamación crónica Proteína e reactiva 5 (14%) 11 (31.5) Fibrinógeno aumentado Tabla 2. La alteración inflamatoria con incremento de fibrinógeno es más frecuente 3. Alteración asociada metabólica 16 (45.7%) Hiperlipidemias Tabla 3. Las hiperJipidemias son frecuentes en los casos de enfennedad tromboembóJica acortada de principio a fin en todos sus paráme­ tros, por aumento de los procoagulantes. Se puede asociar al acortamiento de las pruebas de Tiempo de protrombina, Tiempo de tromboplastina par­ cial y sobre todo al de la tolerancia a la heparina in vitro Hay hipercoagulabilidad global cuando la prueba de generación de trombina está también acortada en todos sus parámetros por disminu­ ción de los anticoagulantes fisiológicos o la prue­ ba de Factor V de Leiden está positiva o de anti­ cuerpos antifosfolipídicos y el anticoagulante lú­ pico son positivas o la Proteína e y S están dismi­ nuídas. Hay hipercoagulabilidad final. cuando la prueba de generación de trombina esta acortada en los parámetros finales, por deficiencia de fibrinoli­ sis cuando la prueba de Euglobulinas está alargada o la prueba de plasminógeno o la antitripsina están disminuidas. Revista Ecuatoriana de Medicina y Ciencias Biológicas Vol. XXVII Números 1 y 2: 11-14., octubre del 2005 12 Cafiizares La hipercoa~ulabilidad inicial es cuando la curva de generación de trombina esta acortada en sus parámetros iniciales, Generalmente se asocia a hiperfunción plaquetaria, cuando el recuento pla­ quetario está aumentado, o cuando la adhesividad y sobre todo la agregación plaquetaria está incremen­ tada o con grumos gruesos. Se considera inflamación crónica con riesgo ateroesclerótico cuando la Proteína C Reactiva es positiva o el Fibrinógeno está aumentado. Finalmente hay riesgo ateroesclerótico cuan­ do el metabolismo lipídico esta alterado y las prue­ bas de colesterol, triglicéridos y LDL están incre­ mentadas. DISCUSIÓN La enfermedad tromboembólica que causa alteraciones isquémicas en varios órganos, espe­ cialmente corazón (infarto de miocardio, enfer­ medad coronaria), cerebro (evento vascular cere­ bral), pulmones (tromboembolia pulmonar), ojos ( trombosis venosa o arterial de retina), miembros periféricos especialmente piernas ( trombosis ve­ nosa profunda, trombosis arteriales periféricas), abdomen (trombosis mesentéricas). Es una enfer­ medad causada por la formación de un trombo in situ o un trombo migrante (émbolo). Las manifes­ taciones dependen del sitio u órgano afectado pe­ ro la etiología es la misma, que es una oclusión vascular de una arteria o vena. Este concepto etiológico es fundamental ya que el tratamiento es el mismo, independientemente del sitio afectado (14). Considerando la etiología única de la oclusión vascular el tratamiento es también el mismo. Consis­ te en dar tratamiento anticoagulante en los casos de hipercoagulabilidad y de antiagregantes plaquetarios en casos de hiperfunción plaquetaria. El tratamiento anticoagulante se inicia con heparina por 3 a 4 días mientras hace efecto el anticoagulante oral, warfari­ na sódica, para luego continuar esta medicación por un tiempo de 3 meses para los primeros episodios, y de 6 meses para los cuadros recidivantes. El control debe ser apropiado ya que la deficiencia en la dosis propiciará que la trombosis progrese y nuevos episo­ dios se presenten; o el exceso en la dosis propiciará hemorragias, que a veces pueden poner en peligro la vida del paciente. Eso quiere decir que el control de­ be ser estricto y apropiado. Se lo realiza con la prue­ ba de TIempo de Protrombina cuyo porcentaje debe estar entre 25 y 30% y el INR (Intemational Norma­ lized Ratio) debe estar entre 2 a 3. Nosotros comple­ mentamos este control también con la prueba de To­ lerancia a la Heparina in vitro, ya que da mayor se­ guridad en el control del efecto del medicamento so­ bre la coagulación global. El TIempo de Protrombina solamente mide el efecto sobre el complejo protrom­ bínico, que es afectado por la warfarina y la Toleran­ cia a la Heparina in vitro mide el efecto sobre la coa­ gulación global, inclusive el efecto de las plaquetas, que puede afectarse con el medicamento. Estas prue­ bas inicialmente se las realiza cada 8 días hasta que el índice se estabilice y después se podrá controlar cada mes. El tratamiento con antiagregantes plaquetarios es muy seguro, se puede controlar con la prueba de agregación plaquetaria. El exceso de antiagregantes puede dar equímosis pero generalmente no da hemo­ rragias, aunque puede eventualmente potencializar sangrados digestivos en pacientes con gastritis cróni­ cas o antecedentes de úlceras gastroduodenales (15) El estudio de las enfermedades tromboembó­ licas significa que debemos evaluar los factores predisponentes: Obesidad, diabetes, hipertensión, enfermedades malignas, tabaquismo, sedentarismo, anticonceptivos, hábitos alimenticios con exceso de grasas, predisposición familiar y otros que ya seña­ lamos. Lo más importante, estudiar la hipercoagu­ labilidad en todos sus parámetros, así como tam­ bién la presencia de enfermedades inflamatorias crónicas y la hiperlipidemia (16) Por todo lo antes señalado es fundamental que antes de iniciar un tratamiento con anticoagu­ lantes se debe procurar estudiar la hipercoagulabili­ dad, para dar el tratamiento adecuado ya sea anti­ coagulantes orales o antiagregantes plaquetarios. No se debe usar ambos por que se potencializa el riesgo hemorrágico. En los casos que se inició la anticoagulación sin el estudio de hipercoagulación hay que esperar que termine el anticoagulante para estudiar la hipercoagulabilidad. En nuestro estudio hemos encontrado que la principal causa es la hipercoagulabilidad global ( 83%), no hubo hipercoagulabilidad en pocos casos 17%). En la hipercoagulabilidad la más frecuente fue por exceso de procoagulantes (29%), de ellos el más frecuente es el síndrome antifosfolipídico (14.6%) que causa en gente joven trombosis veno­ sas profundas, después exceso de factores de coa­ gulación como el Factor II (8.6%) y vm (5.8%). Sigue en frecuencia la hipercoagulabilidad Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vol. XXVII Números 1 y 2: 12-14, octubre del 2005 Importancia del estudio de la hemostasia y coagulación en las enfennedades tromboembólicas global por deficiencia de anticoagulantes fisiológi­ cos (17%), que cuando son en personas jóvenes de corta edad se las llama trombofilias generalmente congénitas (17), de la cual la más frecuente es la de­ ficiencia de Antitrombina III (8.6%), sigue la baja de Proteína C (5.7%) Y en poca frecuencia la presencia de Factor V Leiden (2.7%), que también afecta el funcionamiento del anticoagulante Proteína C. La hipercoagulabilidad por deficiencia de la fibrinolisis (23%) se presenta con mayor frecuencia por deficiencia de plasminógeno (11.4), menos por baja en la ?2 antitripsina (5.8%) y no identificables (5.8%) La hipercoagulabilidad por aumento en la ac­ tividad plaquetaria (14%) es más porincremento en el recuento plaquetario o trombocitosis y trombofi­ lias (8.6%) y menos por hiperagregación plaqueta­ ria (5.8%). En unos pocos casos no se pudo detectar hi­ percoagulabilidad (17%). En algunos casos se encontró el parámetro inflamatorio presente especialmente manifestado por un aumento en el fibrinógeno (31.5%), menos por la presencia de la Proteína C Reactiva (14%). Finalmente en un buen número de casos se pudo comprobar la presencia de una alteración de los lípidos con incremento en las LDL (45%). Lo importante de este estudio es haber com­ probado que la hipercoagulabilidad es muy frecuen­ te como causa de enfermedad tromboembólica, se asocia a hiperlipidemias y procesos inflamatorios crónicos. Para poder establecer un tratamiento adecua­ do y sostenido, ya sea con anticoagulantes orales, como la warfarina, o con antiagregantes plaqueta­ rios como la aspirina, es importante estudiar la pre­ sencia de hipercoagulabilidad y sus diferentes pará­ metros. No es conveniente asociar los dos medica­ mentos, ya que se potencializa el peligro de hemo­ rragias. Hay casos en que no se puede dar warfarina, como en pacientes de alto riesgo hemorrágico, co­ mo hipertensos severos y de avanzada edad o en embarazadas por el efecto fetotóxico. En estos ca­ sos es conveniente mantenerlos con heparina de ba­ jo peso molecular subcutánea (18). También hay que considerar que en algunos casos en que requieren aspirina, no se puede admi­ nistrarla por su efecto irritante en la mucosa gástri­ ca, en estos pacientes hay que usar otros antiagre­ gantes como Clopidogrel o Tic1ide. En casos muy I3 especiales se está usando antiagregantes especiales, como son los bloqueadores de la Glicoproteína de adhesividad GPIIbIIIa de la membrana plaquetaria (Ximelagatran, Dabigatran) para prevenir la re­ trombosis en las angioplastias coronarias. Se puede utilizar el tratamiento anticoagulante preventiva­ mente en casos en hay riesgo de formación de coá­ gulos in situ como en la fibrilación auricular y las prótesis valvulares. (19-30) Finalmente hay que mencionar en cuanto al tratamiento que hay casos de trombosis severas o tromboembolias masivas que deben tratarse con trombolfticos. Así como también trombo lisis direc­ ta coronaria en casos de infarto agudo de miocardio reciente. Nosotros hemos utilizado para estos casos la Estreptoquinasa intravenosa con muy buenos re­ sultados (31). BmLlOGRAFÍA l.- MARCUS, A.J. Thrombosis and inflamationas multicellu­ lar prosses: patbophysiologic significance of transcellular metabolismo Blood 1990; 76: 1903-1907 2.- BADIMON, I. Endotelio, trombosis y arteriosclerosis. Rev. Iberoamer. Tromb. Hemostasis. 1998; 11 (Suplemen­ to 1): 38-41 3.- BADIMON, l.; MERINO, A; BADIMON, U. Trombosis arterial y activación plaquetaria. Fisiopatología. Rev Ibe­ roamer. Tromb. Hemostasisl992; 4: 73-80 4.- ROSENDAAL, ER.; HELMERHORST, EM.; VADEN­ BROUCKE, J .P. Oral contraceptics, honnon replacement tberapy and thrombosis. Thromb. Haemost. 2001; 86:1I2­ 123 5.- ROUVIER, J.; SCAZZIOTA, A. Factores y marcadores de riesgo trombótico. Rev. Iberoamer. Trombo. Hemostasis 1994;7: 192-209 6.- LAZZARI, M.A Marcadores trombóticos. Epidemiología de factores de reisgo pertenecientes al sistema de hemosta­ sia. CLAHT. Simposium Internacional. Río de Janeiro, 16-17 de julio 1998: 47-50 7.- FENSTEIN, D.; GREEN, D.; FEDERlCI, AB.; GOOD­ NIGHT, Jr. S.H. Diagnosis and management of patients witb spontaneously adquired inhibitors of coagulation. He­ matology 1999. American Society of Hematology. Educa­ tional Program Book. New Oeleans, 3-7 December 1999, 192-205 8.- DAVIS, M.; HAGEN, P.O. The vascular endotbelium. A new horizon. Ann. Surger. 1993; 1I8: 593-609 9.- LOWE, G.D.; FORBES, C.D. Bl00d rheology and tbrom­ bosis. Clinics in Haematology. Thrombosis. Prentice CRM. W.B. Saunder, London 1981; 10: 343-367 10.- KROLL, M.H.; HELLMUMS, J.D.; MCINTIRE, L.v.; SCHAFER, A. MONK, EJ. Platelets and shear stress. Blood 1996; 88: 1525-1541 11.- ULUTIN, T.; UCYSIK, N.; HAYTAM, C.; KMOKAK, K.; SULTUYBEK. G.; ULKII, B. As a fibrinolytic system indi­ cator. Plasma tPAlPAl-1 complex in patients witb coronary artery disease. XIII tb. Meeting of tbe Intemational Society of Hematology (European & Africa Division) Abstraes Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vol. XXVII Números 1 y 2: 13-14, octubre del 2005 o 14 Book. Instanbul Türkiye. 3-8 September 1995,997 12.- CAÑIZARES, C. La hemostasia. Las Plaquetas FUN­ DACYT. Quito, 1997, 11-17 13.- SCHAFER, A.L Focussing of the clot. Normal and patho­ 10gic mechanisms. Ann. Rev. Med. 1987; 38: 211-220 14.- FUSTER, V. Progression-regression of atherosclerosis molecular cellular and clinical bases. Circulation 1992; 86 (suppl. 111): 123 15.- BAUER, KA; GOODNlGHT, M.D.; RlDKER, P.M. Hy­ percoaguble. Translocation of risk factor to clinical practi­ ce. Hematology 1998 American Society of Hematology. Education Program Book. Miami, 4-8 December, 255-273 16.- GRANT, P.J. The genetics of atherothrombosis disorders: a clinician's view. J. Thromb. Haemost. State of the Art 2003. XIX Congress of the International Society on Th­ rombosis and Haemostasis. 2003; 1: 1381-1390 17.- BAUER, KA Update in thrombofilia. Hematology 1999; American Society of Hematology Education Program Book. New Orleans, 3-7 December 1999,231-235 18.- YACOVALCA, T.; ALTER, M. Antiacoagulant for venous thromboembolism., what are the current options? Post­ grad. Med. 2000; 108:43-46,51-54 19.- JUFFE, A.; CUENCA, J.; PASALOOOS, J.A. Cirugía de revascularizaci6n miocárdica y/o trombolisis. Rev. Iberoa­ mero Tromb. Hemostasis. 1994; 7: 174-175 20.- WEITZ, J.L; MIDDELOORP, S.; GEERTS, W.; HEIT, J.A. Thrombophilia and New anticoagulant drugs. Hema­ tology 2004. American Society of Hematology. Education Program Book. San Diego, California 4-7 December 2004, 434-438 21.- MAHAFFEY, K.; FERGUSON, J. American College of Cardiology 2004 Scientific Sessions 2004 22.- OLSSON, S.B. Executive Steering Committee on behalf of the SPORTlF III Investigators. Stroke prevention with the oral direct thrombin inhibitor ximelagatran compared with warfarin in patients with non-valvular atrial fibrilla­ tíon (SPORTIF Ill): randomised controlled trial. Lancet. 2003;362: 1691-1698 23.- HALPERlN, J.L. for the Executive Steering Committee on behalf of the SPORTIF V Investigators. Stroke Preven­ tion Using the Oral Thrombín Inhibitor Ximelagatran in Cañizares Patients with Nonvalvular Atrial Fibrillation V - SPOR­ TIF V. Program and abstracts from the American Heart As­ sociation Scientific Sessions 2003; November 9-12,2003; Plenary Session VII: Late Breaking Clinical Trials. 24.- WALLENTIN, L.; WILCOX, R.G.; WEAVER, W.D.; EMANUELSSON, H.; GOODVIN, A.; NYSTROM, P. y col; ESTEEM Investigators. Oral ximelagatran for secon­ dary prophylaxis after myocardial infarction: the ES­ TEEM randomised controlled trial. Lancet. 2003; 362: 789-797 25.- BROUWER, MA; VAN DEN BERGH, P,J.; AENGE­ VAEREN, W.R.; VEEN, G.; LUUTEN, H.E.; HERTZ­ BERGER, D.P. Y col. Aspirin plus coumarin versus aspirin alone in the prevention of reocclusion after fibrinolysis for acute myocardial infarction: results of the Antithrombotics in the prevention of reocclusion in coronary thrombolysís (APRlCOT)-2 trial. Circulation 2002; 106: 659-665. 26.- VAN ES, R.E; JONKER, JJ.; VERHEUGT, EW.; DEC­ KERS, J.w.; Grobbee DE; 360: 109-113. 27.- THE Clopidogrel in Unstable Angina to Prevent Recurrent Events Trial Investigators. Effects of Clopidogrel in Addi­ tion to Aspirin in Patients with Acute Coronary Syndro­ mes without ST-Segment Elevatíon. N Engl J Med 2001; 345: 494-502 28.- COTTER, G.; SHEMESH, E.; ZEHAVI, M.; DINUR, l.; RUDNICK, A.; MILO, O. y col. Lack of aspirin effect: Aspirin resistance or resistance to taking Aspirin? Am Heart J 2004; 147: 293-300. 29.- YORK, M.; AGARWAL, A.; EZEKOWITZ, M. Physi­ cians' attitudes and the use of oral anticoagulants: surve­ ying the present and envisioning future. J Thromb Throm­ bolysis,2003; 16: 33-37 30.- MEZZANO, D.; LEIGTHON, E Haemostatic Cardiovas­ cular Risk Factors: Differential Effects of Red Wine and Diet on Healthy Young Population. Liga del Mediterrá­ neo. Eslovenia, 2204 31.- LANGE, R.A.; HlLLIS, L.D. Concurrent antiplatelet and fibrinolytic therapy. New Eng J Med 2005; 352: 1248­ 1250. D Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vol. XXVII Números 1 y 2: 14-14, octubre del 2005 Desarrollo neural de la rana Colostethus machalilla (ANURA: DENDROBATIDAE) Vanessa Noboa y Eugenia M. del Pino Pontificia Universidad Católica del Ecuador, Escuela de Ciencias Biológicas, Apartado 17-01-2184, Quito, Ecuador. RESUMEN. Se analizó el desarrollo neural de la rana dendrobátida Colostethus machalilla, que es endémica a la costa ecuatoria­ na y deposita sus huevos en la hojarasca. Se realizaron inmunotinciones en montaje entero con anticuerpos neurales específicos. Se encontró retardo en el desarrollo neural de C. machalilla en comparación con la rana acuática Xenopus laevis. PALABRAS CLAVE. Néurula, tubo neural, Colostethus machalilla, Xenopus laevis, Gastrotheca riobambae. ABSTRACT. This work studies the neural development of the dendrobatid frog Colostethus machalilla that is endemic to the Ecuadorian coast. It has a terrestrial way of life and it deposits its eggs directly on the ground. Immunostaining in whole embryos was done to study the pattern of neural molecular markers. The neural development of C. machalilla is retarded in comparlson with the aquatic frog Xenopus laevis. KEYWORDS. Neurula, neural tube, Colostethus machalilla, Xenopus laevis, Gastrotheca riobambae. INTRODUCCIÓN En este trabajo se estudia el desarrollo neural de la rana Colostethus machalilla desde el estadio 13, cuando se hace evidente la placa neural, hasta el estadio 18, que representa a la néurula tardía. Con este estudio se pretende do­ cumentar, en parte, las diferencias en el desa­ rrollo neural que existen entre C. machalilla y la rana acuática Xenopus laevis. Se han deter­ minado los patrones de expresión de las proteí­ nas neurales NCAM y el antígeno 209 median­ te inmuno reacción específica en montaje ente­ ro de los embriones para identificar al tubo neu­ ral y a las células de la cresta neuraL Las ranas venenosas (Dendrobatidae) son un grupo monofilético distribuidos en los trópi­ cos de Sudamérica y Centroamérica (1). El gé­ nero Colostethus presenta una coloración críp­ tica y no posee toxinas en la piel (1). Colostet­ hus machalilla es una especie endémica de la costa ecuatoriana, con huevos terrestres. Los machos presentan cuidado parental del nido te­ rrestre y transportan los renacuajos en su espal­ da hasta depositarlos en el agua (2). Esta rana requiere aproximadamente 5 días desde la fe­ cundación hasta la presencia de la placa neural y 21 días para la eclosión de los renacuajos (3). En contraste, los embriones de X. laevis eclo­ sionan a los cuatro días (4). El desarrollo de C. machalilla en el nido terrestre ha sido caracte­ rizado en 25 estadios (3). La molécula de adhesión celular neural, N-CAM, fue la primera molécula de adhesión celular aislada de tejido neural (5). Se la identi­ ficó como una molécula de adhesión célula-cé­ lula independiente de calcio y estructuralmente se la clasifica en la superfamiJia de las inmuno­ globulinas (5). Estudios realizados en el pollo y en el ratón demostraron que las tres isoformas principales de NCAM son el resultado del em­ palme (splicing) alternativo de un solo gen, que se encuentra en el cromosoma 9 de las dos es­ pecies (6, 7). En el desarrollo neural, NCAM tiene un papel indispensable por sus numerosas funciones, que incluyen mediar la adhesión en- Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vol. XXVII Números 1 y 2: 15-20, octubre del 2005 D Noboa y del Pino 16 tre células neurales, estimular el crecimiento axonal y regular la plasticidad sináptica en pro­ cesos como la memoria y el aprendizaje (8). Por su importancia en el desarrollo, los genes que codifican para moléculas de adhesión celu­ lar son altamente conservados a lo largo de la evolución (6). Comparaciones entre las secuen­ cias de NCAM entre el pollo y el ratón han de­ terminado que hay un 85% de semejanza, ade­ más estudios inmunológicos y ensayos de ad­ hesión celular mostraron que la estructura y función de NCAM se ha preservado entre ver­ tebrados por más de 600 millones de años (6). En X. laevis, Balak el al. (9) describieron la expresión espacio temporal de NCAM me­ diante la utilización de anticuerpos polic1ona­ les. Las inmuno detecciones mostraron que N­ CAM se expresa en la placa neural, el sistema nervioso central en desarrollo, los nervios cra­ neales y algunos ganglios. En cambio, la cresta neural resultó negativa. Se concluyó que, en es­ ta especie, NCAM se expresa después de la in­ ducción neural, de forma transitoria, y tiene co­ mo función la regulación de la morfogénesis de la placa y tubo neura} y del desarrollo de fibras nerviosas. Durante la neurulación, ocurre la migra­ ción de las células de la cresta neural. Estas cé­ lulas tienen un papel esencial en el desarrollo del sistema nervioso periférico y otros deriva­ dos. Son células progenitoras multipotentes y presentan un carácter prominente y conservado entre los embriones de vertebrados (10). Exis­ ten 4 poblaciones de cresta neural: la cresta neural craneal, del tronco, vago y sacral. La cresta neural se origina en la zona limítrofe en­ tre la placa neural y la futura epidermis, en los pliegues neurales (11). El cerebro posterior o rombencéfalo se di vide en 8 compartimentos más pequeños co. nocidos como rombómeros (12). Estos rombó­ meros representan territorios diferenciados. To. dos los rombómeros producen células de la cresta neural craneal (13). La cresta neural cra­ neal está involucrada en el desarrollo de la ca­ beza y migra en tres ramas discretas: mandibu­ lar, hioidea y branquial (14). En el ratón y en el pollo, se ha observado que las células de la cresta neural migran a los tres primeros arcos branquiales en ramas separadas por espacios vacantes de migración, adyacentes a los rom­ bómeros 3 y 5 (15). En este trabajo se ha utilizado el anticuer­ po anti-NCAM para reconocer su expresión en embriones tempranos de C. machalilla y el an­ ticuerpo contra el antígeno neural 2G9 para identificar su expresión en la cresta neural de esta rana. El anticuerpo 2G9 reconoce regiones de la cresta neural craneal y del tronco en la ra­ na Gastrotheca riobambae (16). En Xenopus laevis, el anticuerpo 2G9 reconoce regiones del sistema nervioso central en algunas ramas de la cresta neural craneal, en la cresta neural del tronco y en nervios craneales (17). MATERIALES Y METODOS Se realizaron inmuno detecciones en montaje entero de los embriones de Colostet­ hus machalilla utilizando dos anticuerpos mo­ nocIonales, el anticuerpo 4D, que es específico para la proteína neural NCAM, y el anticuerpo 2G9, que marca la cresta neural. Se estudió la expresión de estas proteínas en embriones de los estadios de la neurulación temprana, desde la finalización de la gástrula, hasta el estadio de formación de arcos branquiales, que correspon­ de a los estadios 13 al 18 de C. machalilla (3). Para cada estadio del desarrollo se realizó la in­ muno detección en tres o más embriones por estadio. Se utilizaron las metodologías estándar para la inmuno detección en montaje entero en embriones de anfibios (16). Los embriones fue­ ron fotografiados en montaje entero. Algunos embriones fueron seccionados. RESULTADOS El marcador neural NCAM en C. machulilÚl La néurula temprana (estadios 13 y14) se caracteriza morfológicamente por la aparición de la placa neural. Embriones del estadio 13 fueron negativos para NCAM (no se muestra). Revista Ecuatoriana de Medicina y Ciencias B,iológicll8 - Vol. XXVII Números 1 y 2: 16-20, octubre de12005 -------~- - ._­ D Desarrollo neural de la rana Colostethus machalilla (ANURA; DENDROBATIDAE) 17 17 Fig 1. Expresión del antígeno 2G9 y de la molécula de adhesión celular neural NCAM en embriones C. ma­ chalilla. El estadio de los embriones se indica en la parte superior derecha de cada microfotografía. (A) Inmuno de­ tección de un embrión de estadio 16 con el anticuerpo 2G9. Se detecta la cresta neural craneal (nc) a los lados del tu­ bo neural. (B) Inmuno detección de un embrión de estadio 17 con anti-NCAM para destacar el tubo neural. Los ner­ vios faciales (VII) recorren paralelos a1límite rostral de los otocistos (o). Los límites de los rombómeros son ligera­ mente visibles. Los otocistos se encuentran localizados frente al rombómero 4 (r4), como ocurre en X. laevis. (C) In­ muno detección de un embrión de estadio 18 con el anticuerpo 209. La localización profunda de la cresta neural (nc) a los lados del tubo neural (nt), se observa en esta sección transversa del cerebro posterior. Las barras representan 100 11m en Ay B Y200 11m en C. La expresión de NCAM se pudo detectar por primera vez en embriones del estadio 14 de C. machalilla. En cambio, embriones de X. laevis en el estadio de la placa neural son NCAM po­ sitivos (no se muestra). La néurula media se caracteriza por el acercamiento de los pliegues neurales y por el cerramiento del tubo neural (estadios 15-16). El estadio 16 se destaca porque durante éste se cierra el tubo neural (Fig. lA). El patrón de expresión de NCAM en este estadio se restrin­ ge al tubo neural (no se muestra). Los estadios 17 y 18 representan la néurula tardía. En las inmuno detecciones para NCAM del estadio 17, se observó la especificidad de expresión de esta molécula, que estuvo restringida al tu- Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vol. XXVII Números 1 y 2: 17-20. octubre del 2005 ro 18 Noboa y del Pino bo neural (Fig. lB). Se pueden apreciar ciertas estructuras específicas para NCAM como el tubo neural, los nervios espinales, el nervio craneofacial VII. Se detectó una reacción dife­ rencial entre los diferentes rombómeros, y se han identificado tres de ellos, r1, r2 y r4 por su posición anatómica respecto a los otocistos (Fig. lB). El nervio craneofacial VII surge del rombómero 4, rostral a cada otocisto (Fig. lB). La expresión de NCAM se desvanece en la región de la cola (no se muestra). En la re­ gión de las vesículas ópticas, se puede detec­ tar una reacción positiva para NCAM en la re­ gión exterior y posterior de estas estructuras (no se muestra). El antígeno 2G9 como marcador de la cresta neural en C. machalilla La primera detección de la cresta neural por la expresión del antígeno 2G9 se detectó en embriones avanzados del estadio 14. La expre­ sión del antígeno 2G9 en el estadio 15 se detec­ ta en las ramas branquiales de la cresta neural cefálica y también se puede notar que existe u­ na reacción específica en el cerebro posterior. En esta región se puede observar que entre blo­ ques celulares con reacción positiva existen secciones donde la inmuno detección resultó negativa. Se puede sugerir que estas regiones, con diferente expresión del antígeno 2G9, son los rombómeros del cerebro posterior. Una sec­ ción transversa de un embrión inmunoteñido contra el antígeno 2G9 en el estadio 18 muestra intensa marcación de la cresta neural (Fig. 1C). La misma tiene una localización profunda, a los lados del tubo neural. DISCUSIÓN En C. machalilla, la expresión de N­ CAM se encuentra a 10 largo de todo el tubo neural del embrión, perdiéndose en la yema de la cola, al igual que en X. laevis (9). En la re­ gión del tronco se detectan los nervios espina­ les tal como se reporta en los nervios periféri­ cos de X. laevis (9). En la región cefálica, el ganglio del nervio facial (VII) y las vesículas ópticas fueron NCAM positivas al igual que en G. riobambae (16). La única diferencia es que en C. machalilla, al igual que X. laevis, las ve­ sículas ópticas son más conspicuas que en G. riobambae. El antígeno 2G9 es un antígeno neural extracelular que ha sido detectado en el sistema nervioso central en X. laevis, también ha sido detectado en ratones y en salamandras, lo cual sugiere que se ha conservado a 10 largo de la evolución de los vertebrados (17). El primer es­ tadio donde se reporta la expresión de este antí­ geno en G. riobambae corresponde a una néu­ rula tardía. La marcación positiva se encuentra en las ramas de la cresta neural hioidea, bran­ quio-posterior y vago (16). El sistema nervioso central y las células de la cresta neural de las ra­ mas branquio-anterior y mandibular son nega­ tivos (16). En estadios más avanzados, las ra­ mas de migración de la cresta neural craneal se hacen más prominentes, y la expresión de 2G9 continúa en las ramas mandibular, hioidea y branquio-posterior (16). En el embrión de esta­ dio 14 de G. riobambae, el antígeno 2G9 se ex­ presa en todos los rombómeros, excepto en r3 y r5. Se expresa también en los nervios: trigemi­ nal, facial y vago (16). En C. machalilla, las ramas de migración de la cresta neural craneal no son tan prominen­ tes como las que se encuentran en G. riobam­ bae. El antígeno 2G9 se expresa en la cresta neural craneal y del tronco como en G. riobam­ bae. Las ramas de migración de los arcos bran­ quiales, hioideo y mandibular en C. machalilla fueron determinadas en un trabajo anterior (16). Sin embargo, como no se han logrado dis­ tinguir las dos ramas branquiales, anterior y posterior claramente a lo largo de todo el desa­ rrollo neural, se sugiere que en C. machalilla las dos ramas no se dividen totalmente como ocurre en G. riobambae. Por 10 tanto, las ramas posteriores a la rama hioidea en C. machalilla corresponden a una única rama branquial, no específicamente anterior o posterior. Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vol. XXVII Números 1 y 2: 18-20, octubre del 2005 Desarrollo neural de la rana Colostethus 17Ulchalilla (ANURA: DENDROBATIDAE) El desarrollo neural aparece como una característica altamente conservada a 10 largo de la evolución. Esto se ha determinado gracias a estudios morfológicos y a los patrones de ex­ presión génica de marcadores neurales como NCAM y el antígeno 2G9. Algunas moléculas, como NCAM, son independientes de las estra­ tegias reproductivas, ya que se presenta en pa­ trones similares entre anfibios y en compara­ ción con otros vertebrados. Las observaciones realizadas en la neurulación de C. machalilla brindan una base de comparación con otras es­ pecies. Este estudio demuestra que la expresión de la molécula NCAM, tiene retardo en el ini­ cio de su expresión en los embriones de C. ma­ chalilla en comparación con X. laevis. Este es el aporte principal de este trabajo, que se corre­ laciona positivamente con los eventos de retar­ do en el desarrollo del notocordio y somitas de C. machalilla (18, 19). En efecto, el notocordio de C. machaUlla inicia su alargamiento una vez que se ha cerrado el blastoporo, en tanto que en X. laevis el notocordio se alarga mucho antes, en el estadio de la gástrula media (18). De mo­ do similar, los somitas de X. laevis se pueden detectar y expresan marcadores moleculares específicos en la neurula media (19). Estos eventos de retardo morfológico y molecular que se han detectado en el desarrollo temprano de C. machalilla son indicadores de diferencias en los mecanismos de control del desarrollo embrionario. Por tal motivo, esta rana es un or­ ganismo de interés para los estudios comparati­ vos de la embriogénesis. Agradecimientos Agradecemos a F. Brown, F. Sáenz, O. Pérez y S. Benítez por su ayuda, profesional y apoyo. Igualmente se reconoce la colaboración de D. Donoso, I. Alarcón e I. Moya. Este trabajo recibió el apoyo de las becas de investigación de la Pontificia Universidad Católica del Ecuador y del proyecto FUN­ DACYT PFN 034. 19 BmLlOGRAFÍA 1.- SANTOS, J.c.; COLOMA, L.A. & CANNATELLA D.C. Multiple, recurring origins of aposetism and diet speciali­ zation in poison frogs. Proceedings oC the NationaJ Aca­ demyoCSciences, 2003; 100: 12792-12797. 2.- QUIOUANOO-UBILLÚS, A. Bruptflege bei Pfegiftfros­ chen. DRACO, 2000; 3: 16-23. 3.- DEL PINO, E.M.; ÁVILA, M.E.; PÉREZ, O.D.; BENÍ­ TEZ, M.S.; ALARCÓN, l.; NOBOA, V. & MOYA, I.M. Development of the dendrobatid frog Colostethus macha­ lilla. International Journal of Developmental Biology, 2004; 48: 663-670. 4.- NIEUWKOOP, P.D. & FABER, J. Normal Table of Xeno­ pus laevis (Daudin). OarIand Publishing, New York, Lon­ don, 1994. 5.- WOLPERT, L.; BEDDINOTON, J.; JESSELL, T.; LAW­ RENCE, P. & MEYEROWITZ, E. PrincipIes of DeveIop­ ment. Current Biology Ltd. USA, 1998. 6.- RUTISHAUSER, U. & JESSELL, T.M. Cell Adhesion MoIecuIes in Vertebrate Neural Development. Physiologi­ ca] Reviews, 1983; 68: 819-859. 7.- WATANABE, M.; FRELINOER, A.L. & RUTISHAU­ SER, U. Topography of N-CAM struetural and funetional determinants. I. Classifieation of monoclonal antibody epitopes. Journal of Cell Biology, 1986; 103:1721-1727. 8.- KISELYOV, V.V.; SKLADCHIKOVA, O.; HINSBY, A.M.; HOLME JENSEN, P.; KULAHIN, N.; VLADIS­ LAV, S.; PEDERSEN, N.; TSETLIN, v.; POULSEN, EM.; BEREZIN, V. & BOCK, E. Struetural Basis for a Di­ rect Interaetion between FOFRl and NCAM and Eviden­ ce for a Regulatory Role of ATP. Structure, 2003; 11: 691-701. 9.- BALAK, K.; JACOBSON, M.; SUNSHINE, J. & RUTIS­ HAUSER, U. Neural eell adhesion molecule expression in Xenopus embryos. Developmental Biology, 1987; 119: 540-550. 10.- BELLMEYER, A.; KRASE, J.; LINDOREN, J. & LA­ BONNE, C. The Protooneogene c-Mye Is an Essential Re­ gulator of Neural Crest Formation in Xenopus. DeveJop­ mental Cell, 2003; 4: 827-839. 11.- MAYOR, R. & AYBAR, M.J. Induetion and development of neural erest in Xenopus laevis. Cell & Tissue Re­ search, 2001; 305:203-209. 12.- GILBERT, S. E Developmental Biology. Sinauer Associa­ tes, Ine. Publishers. Sunderland, Massaehusetts, USA, 2000. 13.- BIROBAUER, E.; SECHRIST, J.; BRONNER-FRASER, M. & I''RASER, S. Rhombomerie origin and rostroeaudaI assortment of neural erest eells revealed by intravital mi­ eroseopy. Development, 1995; 121:935-945. 14.- CARL, T.E; DUFTON, c.; HANKEN, J. & KLYM­ KOWSKY, M.W. Inhibition of Neural Crest Migration in Xenopus Using Antisense Slug RNA. Developmental Biology, 1999; 213: 101-115. 15.- CHRISTIANSEN, J.H.; COLES, E.O. & WILKINSON, D.O. Molecular control of neural erest formation, migra­ tion and differentiation. Current Opinion in Cell Bio­ logy, 2000; 12:719-724. Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vol. XXVII Números 1 y 2: 19-20, octubre del 2005 o 20 16.- DEL PINO, E.M. & MEDINA. A. Neural development in the marsupial frog Gastrotheca riobambae. International Journal of Developmental Biology, 1998; 42:723-731. 17.- JONES, EA & WOODLAND, H.R Spatial aspects of neural induction in Xenopus laevis. Development, 1989; 107:785-791. 18.- BENÍTEZ, M.S. & DEL PINO, E.M. Expression of Bracbyury during development of the dendrobatid frog Colostethus machalilla. Developmental Dynamics, 2002; 225:592-596. Noboa y del Pino 19.- MOYA, I.M. & DEL PINO, E.M. Sornitogénesis de Co­ lostethus machalilla (Anura: Dendrobatidae). Memorias del ler Congreso Nacional de Investigación, Tecnología e Innovación. FUNDACYT, Quito, 2005; (en prensa). e Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vol. XXVII Números 1 y 2: 20-20, octubre del 2005 Cariotipos Metafásicos de Drosophila inca y D. yangana, subgrupo inca, grupo repleta. Ana Beatriz Mafla Laboratorio de Genética, Escuela de Ciencias Biológicas, Pontificia Universidad Católica del Ecuador. Apartado 17-01-2184. Quito-Ecuador. [email protected] Recibido 15 marl:O, 2005. Aprobado 15 junio, 2005 RESUMEN. Las especies del género Drosophila: D. inca y D. yangana pertenecientes al nuevo subgrupo inca del grupo de es­ pecies repleta son caracterizadas en términos de propiedades de los cromosomas metafásícos de ganglios cerebrales. En la descrip­ ción de los cariotipos se introduce la nomenclatura recomendada por Levan et al. (1964) con el objeto de precisar los rearreglos a nivel de las porciones heterocromáticas las cuales pasan desapercibidas en los análisis cariotípicos de cromosomas politénicos. PALABRAS CLAVE. Cario tipos metafásicos, heterocromatina, subgrupo inca. ABSTRACT. This work supply the information about the metaphase karyotypes of Drosophila inca and D. yangana ; similarities and differences. introducing in the description the nomenclature recommended by Levan et al.(l964) it allow to detect arrange­ ments at the leve! of the heterochromatin, which are unprovided in the polytene chromosomes. KEYWORDS. Metaphase karyotypes, heterochromatin, inca subgroup. Introducción o El subgrupo inca del grupo repleta está constituido por tres especies: Drosophila inca Dobzhansky & Pavan, 1943 (1); D. huancavil­ cae Rafael & Arcos, 1989 (2) YD. yangana Ra­ fael & Vela, 2003 (3), estas dos últimas descu­ biertas en el Ecuador. Se desconocen las parti­ cularidades biológicas de este nuevo subgrupo de especies; por lo que se ha abordado estudios que permitan su caracterización y eventual­ mente el establecimiento de las relaciones filo­ genéticas, tanto dentro del subgrupo inca, como con los miembros de los otros subgrupos. Los ciclos de vida de D. inca y D. yanga­ na, se han estudiado en condiciones de labora­ torio, las dos especies presentan una duración promedio, desde la eclosión del huevo hasta la eclosión del adulto, de 22 días; aunque se ob­ serva que Drosophila inca tiene períodos de duración ligeramente menores en las etapas de huevo y primer estadio larvario y mayores en la fase de pupa (4). En cuanto a la información citológica de Drosophila inca es muy limitada por eso se presenta en este trabajo los cariotipos metafási­ cos de D. inca junto a los de D. yangana: sus parecidos y diferencias, introduciendo en la descripción la nomenclatura recomendada por Levan et al. (5). Materiales y Métodos Las muestras analizadas corresponden a tres iso líneas de D. inca y tres de D. yangana fundadas a partir de hembras recolectadas en la naturaleza durante marzo del 2002 en la locali­ dad andina de Yangana, provincia de Loja, Ecuador (4° 23' S, 79° 11' W). Se disectaron larvas de tercer estadio para extraer los gan­ glios cerebrales, en un portaobjeto excavado se colectaron de lOa 12 ganglios cerebrales en so­ lución de Ringer-Drosophila, se transfirieron a otro porta excavado conteniendo una solución de Ringer: co1chicina O,OlM en proporción 2: 1 por 30 minutos; se extrae esta solución y se la Revista Ecuatoriana de Medicina y Ciencias Biológicas Vol. XXVII Números 1 y 2: 21-25, octubre del 2005 22 o Mafia remplaza por KCI 0,075M durante 20 minutos. Después del tratamiento hipotónico se fija el tejido en metanol: ácido acético (3: 1) por míni­ mo 30 minutos al cabo de los cuales se extrae el fijador y se dispersa el tejido en ácido acético 60%, se añade dos gotas de fijador y se gotea en portaobjetos los cuales fueron mantenidos en etanol y en refrigeración; las placas con la suspensión se dejan secar en una plancheta a 65°C (el shock térmico permite una mejor dis­ persión de los cromosomas). Las placas fueron teñidas con Giemsa diluído en agua desioniza­ da en proporción 1:20. Los mejores núcleos fueron fotografia­ dos usando el lente de inmersión Neofluar 100, oculares 8X y aumento 1,25 de la cámara foto­ gráfica obteniéndose una amplificación total de 1000X. Los cariotipos se montaron recortando el perfil de los cromosomas en fotografias con­ vencionales, se midieron los cromosomas y se organizaron en orden descendente. En la identi­ ficación de la forma cromosómica se sigue las recomendaciones de Levan et al. (1964); nomi­ nando a los cromosomas st, sm o T de acuerdo a la posición del centró mero en la región sub­ terminal, submedial o en el punto Terminal, respectivamente. Los idiogramas se construye­ ron normalizando los valores de 20 cariotipos (15dd, 5dd) de D. inca y 32 (27dd, 599) de D. yangana. La normalización se realizó calculan­ do la longitud relativa de cada cromosoma y luego promediando los valores de cada par ho­ mólogo, excepto para el par XY; igual procedi­ miento se aplicó en la normalización del índice centrómerico. Resultados y Discusión El número diploide en las dos especies es igual; 2n == 12. Tomando como referencia el cariotipo de D. inca que fuera descrito por Dobzhansky & Pavan, 1943 como compuesto de 5 R 1 JI podemos decir que los resultados obtenidos en este trabajo muestran coinciden­ cia parcial, puesto que en las placas mitóticas 1 Del inglés R: Rods obtenidas por suspensión celular se ha podido definir con mayor precisión la posición del centrómero, de modo que de las 5 varillas re­ portadas precisamos que en D. inca del Ecua­ dor los cromosomas más grandes son st y los pequeños son sm. Los cariotipos fueron construidos orde­ nando los cromosomas por tamaño decreciente y reuniendo en el grupo A los 3 cromosomas más grandes: dos pares de autosomas y el par sexual heteromórfico, en este conjunto resalta el cromosoma Y por su heteropicnosis en tanto que el X se confunde con los demás. En el gru­ po B se reúne a los tres auto somas pequeños; en este conjunto es claramente identificable el par autosómico 6 por su menor tamaño y hetero­ picnosis. II 3 4 •• 5 le 6 Fig. 1 a-b. a) Núcleo mitótico de ganglio cerebral de Drosophila inca Q, b) Cariotipo mostrando los grupos cromosómicos A y B. La barra corresponde a 4 !l. En D. inca se evidencian tres pares de cromosomas st y 3 sm (Fig. la, b). El grupo A con cromosomas cuyas longitudes relativas (LR) están entre 11 ,35 a 9,07; el más grande corresponde al cromosoma sexual X, seguido del auto soma 2 con LR 9,92; y el auto soma 3 con LR de 9,07; estos tres cromosomas tienen el centrómero en posición s1. El grupo B reúne = varillas; J =forma de esta letra; en la nomenclatura clásica de cromosomas de Drosophila. Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vol. XXVII Números 1 y 2: 22-25, octubre del 2005 J Cariotipos Metafásicos de Drosophila inca y D. yangana, subgrupo inca, grupo repleta 23 Tabla 1. Longitud relativa y morfología de los cromosomas de Drosophila inca y D. yangana Grupo A D. inca D.yangana D GrupoB X 2 Y 3 4 5 6 11,35 9,92 9,21 9,07 8,32 7,08 5,08 st st sm st sm sm sm 10,192& 10,94 10,06 9,58 8,90 7,73 2,58 2,14 sm sm sm sm sm sm T T a los 3 auto sornas más pequeños con centró­ mero sm y cuyas LR están entre 8,32 y 5,08; el autosoma 6 aparece fuertemente heteropicnóti­ co y su longitud corresponde a poco más de la mitad de la longitud del cromosoma más gran­ de (X). En orden decreciente el heterocromoso­ ma y ocupa la tercera posición con LR 9,21 y centrómero sm. (Tabla 1). En D. yangana hay 5 pares sm y uno T (Fig.2 a, b). El cromosoma más grande corres­ ponde al autosoma 2 con LR de 10,94 seguido por el cromosoma X con una LR de 10,19; yel autosoma 3 con LR 9,58, todos los del grupo A tienen el centrómero sm. El grupo B compren­ de a los autosomas 4 y 5 de centrómero también sm y de LR 8,90 y 7,73 Y al6 T y LR 2,58; es­ te último también es heteropicnótico y su tama­ ño corresponde a un cuarto de la del cromoso­ ma más grande (autosoma 2). En los cariotipos de D. yangana el hete­ rocromosoma y de LR 10,06 también ocupa la tercera posición (Tabla 1); es heteropicnótico con centrómero sm y presenta una constricción secundaria en el tercio distal del brazo largo (Fig. 3b). Llama la atención la coincidencia de este marcador con lo reportado para D. uniseta y D. starmeri miembros del subgrupo mulleri (6, 7); otro sub grupo del grupo repleta. Además en D. yangana se han registrado núcleos con uno o dos cromosomas supernu­ merarios de LR 2,14, posición centromérica T y heteropicnóticos. (Tabla 1). B Comparado con el cariotipo de D. inca el de D. yangana presenta las siguientes diferen­ cias (Fig. 3) : 10 el cromosoma X ocupa la se­ gunda posición en orden decreciente, 2 0 los miembros del grupo A tienen centrómeros sm; 3o en el grupo B el auto sorna 6 es T y corres­ ponde a la mitad del tamaño del de D. inca, en las placas metafásicas aparece corno un punto; 4 o resalta la presencia de cromosomas supernu­ merarios que son considerados excepcionales en el género Drosophila; el cromosoma B su­ mado al cromosoma 6 da una longitud corres­ pondiente al 6 de D. inca (Tabla 2). a) D b) A ., a. a, Xy B ,~ 4 2 S I 3 •• 6 Fig. 2 a-b. a) Núcleo mitótico de Drosophila yan­ ganad, b) Cariotipo mostrando los grupos cromosómi­ cos A y B. La barra corresponde a 4 !l. La flecha señala la constricción secundaria del cromosoma Y. Revista Ecuatoriana de Medicina y Ciencias Biológicas VoL XXVII Números I y 2: 23·25, octubre del 2005 ------------ Mafla 24 b) a) X st 2 st 3 4 5 6 y 2 X 3 4 st sm sm sm sm sm sm sm sm 5 sm 6 T y sm Fig. 3 a-b. Idiogramas de a) Drosophila inca, grupo A cromosomas st. Autosoma 6 sm. b) Drosophila yangana, grupo A cromosomas sm. Autosoma 6 T. Tabla 2. Comparación del porcentaje de heterocromatina en los cromosomas de DJnca y D.yangana Drosophila inca D.yangana X 4,84 5,32 2 4,19 5.63 3 4,33 4,75 4 4,29 4,62 5 3.79 4.23 6 10,00 5,17 Cromosoma C----_ D 4,30 B y 15,35 16,77 Total 46,79% 50,79"10 Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vol. XXVII Números 1 y 2: 24-25, octubre del 2005 D Cariotipos Metafásicos de Drosophila inca y D. yangana, subgrupo inca, grupo repleta Las dos especies son semejantes en la morfología de los autosomas 4 y 5, del sexual Y que ocupa la tercera posición, así como la hete­ ropicnosis de los cromosomas Y y 6. Como 10 señala White (8) en Drosophila los cromosomas mitóticos permiten detectar rearreglos a nivel de la heterocromatina, los cuales pasan desapercibidos en los cariotipos de cromosomas politénicos; en este sentido, la presente descripción de los cariotipos de las dos especies de Drosophila, usando la nomen­ clatura recomendada por Levan et al. (1964) nos permite discernir que los brazos cortos de los cromosomas X, 2, 3, 4 Y 5 de D. inca, con­ tienen menor proporción de heterocromatina que los correspondientes de D. yangana. Su­ mando las longitudes de las porciones hetero­ cromáticas (Tabla 2) podemos estimar el conte­ nido total de cada genoma, en D. inca tenemos un valor de 46,79% en tanto que en D. yangana llega a 50,79%. 25 REFERENCIAS 1.- DOBZHANSKY, TH. & PAVAN, C. Studies on Brazilian Species of Drosophila. Separata do Bol. Fac. Fil. Cien. E Letr. Univ. S. Piulo, N° 36-Biologia Geral 1943; W 4: 44-46. 2.- RAFAEL, V. & ARCOS, G .. Subgrupo inca, un nuevo subgrupo del grupo repleta, con descripción de Drosophi­ la huancavilcae nov. sp. (Diptera, Drosophilidae). Evolu­ ción Biológica, Bogotá, 1989; 3 (3): 233 - 243. 3.- RAFAEL, V. & VELA, D. Drosophila yangana sp. nov. un nuevo miembro del grupo repleta, subgrupo inca (Díptera: Drosophilidae). Revista de la Pontificia Universidad Católica del Ecuador, Quito, 2003; 71: 129-139. 4.- MAFLA, A B. Life cycles and fitness components of Dro­ sophila inca and D. yangana (Diptera, Drosophilidae) Iberingia, Sér. Zool., Porto Alegre, 2005; 9S (1); 89-91. 5.- LEVAN, A; FREDGA, K. & SANDBERG, AA Nomen­ clature for centromeric position on chromosomes. Heredi­ tas, 1964; S2 (2). 6.- WASSERMAN, M. & KOEPFER, H. R. Cytogenetics of the South American Drosophila mulleri Complex: The martensis cluster, more interspecific sharing of inversions. Genetics 1979; 93: 935-946. 7.- WASSERMAN, M. Evolution of the repleta Group. In: The Genetics and Biology oC Drosophila. ASHBURNER, M., CARSON, H. L. & THOMPSON, J. N. IR., eds. Academic Press, London, 1982; 3b: 61-139. 8.- WHlTE, M.I.D.. Modes oCSpeciation. W.H.Freeman and Company, San Francisco, 1978, pg. 66. o D Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vol. XXVII Números l y 2: 25-25, octubre del 2005 D D Catorce nuevas especies del género Drosophila (Diptera, Drosophilidae) en el Bosque húmedo montano del Volcán Pasochoa, Pichincha, Ecuador. Doris Velal y Violeta RafaeP Laboratorio de Genética Evolutiva. Escuela de Ciencias Biológicas, Pontificia Universidad Católica del Ecuador, apartado 17-01-2184, Quito, Ecuador. [email protected],[email protected] Recibido 1 marzo 2005, aprobado 15 junio 2005 RESUMEN. En el presente estudio se describen catorce nuevas especies del género Drosophila descubiertas recientemente en el Volcán Pasochoa, las nuevas especies pertenecen a cinco grupos de especies del género l. grupo repleta: Drosophila shuyu; 2. grupo tripunetata: Drosophila pataeorona, Drosophila quillu, Drosophila ninarumi, Drosophila surueueho, Drosophila iehubam­ ba y Drosophila ureu; 3. grupo flavopilosa: Drosophila suni, Drosophila taxohuayeu y Drosophila sisa; 4. grupo guarani: Dro­ sophila euseungu; 5. grupo annulimana: Drosophila yana y dos especies no agrupadas: Drosophila eondormaehay y Drosophila pugyu. PALABRAS CLAVE. Ecuador, Drosophila, nuevas especies, Volcán Pasochoa. ABSTRACT. Fourteen new species of genus Drosophila (Díptera, Drosophilidae) in bumid montane forest Pasochoa Vol­ cano, Pichincha, Ecuador. In the present study, fourteen new species of Drosophila genus are described, all species recently we­ re discovered in the Pasochoa Vo1cano, the new species belong to five Drosophila species group. l. repleta group: Drosophila shu­ yu; 2. tripuetata group: Drosophila pataearona, Drosophila quillu, Drosophila ninarumi, Drosophila surueueho, Drosophila iehu­ bamba and Drosophila ureu; 3. flavopilosa group: Drosophila suni, Drosophila taxohuaycu and Drosophila sisa; 4. guaraní group: Drosophila cuseungu; 5. annulimana group: Drosophila yana and two species no grouped: Drosophila eondormachay and Drosophila pugyu. KEYWORDS. Ecuador, Drosophila, new species, Pasochoa Volcano. INTRODUCCIÓN En trabajos realizados recientemente en el Bosque Pasochoa se han descubierto nume­ rosas nuevas especies del género Drosophila. Los grupos de especies encontrados fueron re· pleta, tripunctata, mesophragmatica, guaraní, flavopilosa y onnychophora (1, 2,3,4, 5 Y 6); estos hallazgos motivaron nuevos estudios en el bosque. Los resultados obtenidos revelan la existencia de 14 nuevas especies más del géne­ ro Drosophila, 12 de ellas pertenecen a los gru­ pos repleta, tripunctata, flavopilosa, guaraní y annulimana, y 2 especies no agrupadas. El bos­ que Pasochoa es el único bosque andino del Ecuador en el que se han realizado estudios so­ bre la drosofauna. La diversidad de especies encontradas en el Pasochoa, nos estaría indi­ cando la existencia de numerosos microhábi­ tats y, sin ser pretenciosas, sospechamos que en el cráter del volcán existen aún más nuevas es­ pecies. MATERIALES Y MÉTODOS En el Bosque Pasochoa, localizado en las laderas del Volcán Pasochoa (0° 28'S, 78° 29'W), se llevaron a cabo recolecciones de moscas del género Drosophila a lo largo de dos transectos de 1 km de longitud cada uno duran­ te un año, desde marzo del 200 1 hasta abril del 2002. El Transecto Uno (TI) va desde los 2 780 a los 2 850 msnm y el Transecto Dos (T2) des­ de los 2 900 a los 3 150 msnm; en cada transec- Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vol. XXVII Números 1 y 2: 27-41. octubre del 2005 o 28 Vela y Rafael to se marcaron 21 puntos de recolección sepa­ rados por 50m uno de otro. La captura de mos­ cas se realizó utilizando trampas de banano, las cuales pennanecieron en el bosque por espacio de 5 a 15 días. Para la identificación de las especies, se tomaron en cuenta sólo a los individuos ma­ chos, cada mosca fue analizada externamente, y la terminalia masculina fue sometida a ebu­ llición en KOH por 10 minutos y luego disec­ tada. El holotipo y paratipos de cada especie es­ tán depositados en el Museo de Zoología de la Pontificia Universidad Católica del Ecuador (QCAZ) en Quito. RESULTADOS Grupo repleta D. shuyu sp. nov (Figs. 1-5,61) 2 0.1 mm CJ 5 Figs. 1-5. Drosophila shuyu sp. nov. holotipo macho: 1, epan­ drio, placa anal y surestilo, vista posterior, lado izquierdo omi­ tido; 2, hipandrio, vista posterior; 3-5, edeago, apodema del edeago y paráfisis, vista ventral, dorsal y lateral izquierda res­ pectivamente. Material examinado. Holotipo Ó (disectado y guardado en microtubo), etiquetado D. shuyu holotipo DVela det. 2003 / Pasochoa, Pichincha, Ecuador DVela Ó col., jun. 2ool. Un paratipo Ó(disectado ~ guardado en microtubo). etiquetado D. shuyu paratipoODVela det. 2003/ Pasochoa, Pichincha, Ecuador DVela col., mar. 2001. Un paratipo Ó (disectado y guardado en microtu­ bo), etiquetado D. shuyu paratipo O DVela det. 2003/ Pasochoa, Pichincha, Ecuador DVela col., nov. 2001. Un paratípoÓ(disectado y suardado en microtubo), etique­ tado D. shuyu paratipo ÓDVela det. 2003 / Pasochoa, Pi­ chincha, Ecuador DVela col., abr. 2002. Holotipo y para­ tipos depositados en QC!\Z. Localidad tipo. ECUADOR, Pichincha: Volcán Pasochoa, 35 km al sur de Quito. Descripción. Cabeza de color marrón os­ curo casi negro polinoso. Ocelos de color ma­ rrón oscuro, triángulo ocelar del mismo color que la cabeza. Ojos de color rojo oscuro. Orbi­ tal media es 1/2 de la primera orbital y más cer­ ca del ojo, cada cerda orbital. nace de una man­ cha oscura. Carina prominente y levemente surcada. Una cerda oral prominente. Antena marrón, arista ramificada, con 3 ramas superio­ res, 2 ramas inferiores, más la bifurcación ter­ minal y pequeñas ramificaciones internas. Lon­ gitud del cuerpo 2.1mm. Tórax de color marrón oscuro y polinoso con manchas oscuras que fonna filas irregulares, de las cuales emergen los pelos acrosticales, ordenados en ocho filas frente a las cerdas dorsocentrales. Escutelo de color marrón oscuro. En las patas, la coxa y el fémur son de color marrón oscuro mientras que el tarso y metatarsos son de color marrón ama­ rillento. Alas claras, longitud 2.6mm. Índice costal 2.3. Abdomen amarillento, del primero al tercer tergitos son completamente oscuros, algunas veces el borde superior es menos pig­ mentado. Del cuarto al sexto tergito se presen­ ta la línea media dorsal dividiendo las bandas marginales en el centro del tergito. La pigmen­ tación de estas bandas marginales cubre la mi­ tad inferior del tergito. Terminalia Ó. Placa anal fusionada al epandrio en la parte media. Surestilo con 14 dientes primarios, 9-10 cerdas marginales y 12 cerdas en la parte baja, 1 en la parte media y 3 en la parte superior del epandrio que presenta Revista Ecuatoriana de Medicina y Ciencias Biológicas· Vol. XXVII Números 1 y 2: 28-41. octubre del 2005 el Catorce nuevas especies del género Drosophila (Díptera, Drosophilidae) en el Bosque húmedo montano del Volcán....... micropubescencia. Edeago quitinizado, en vis­ ta frontal es de forma ovalada y aguda en el ápi­ ce, el cual es menos quitinizado y con una lige­ ra depresión en la punta; gonopodio desnudo y de forma semiovalada. Índice del edeago 1.3. Etimología. "Shuyu" palabra del idioma quichua, que significa manchado. Esta especie presenta manchas en el tórax, característica tí­ pica de las especies del grupo repleta, y de al­ gunas especies del grupo guaraní (5). Grupo tripunctata D. patacorona sp. nov. (Figs. 6-9, 62) Ó Material examinado. Holotipo Ó (disectado y guardado en microtubo), etiquetado: D. patacorona ho­ lotipoÓDVela del. 2003/ Pasochoa, Pichincha, Ecuador DVela col., mar. 2001. Un paratipoÓ(disectado y guar­ dado en microtubo), etiquetado D. patacorona paratipo DVeJa det. 2003 / Pasochoa, Pichincha, Ecuador DVeJa col., abr. 2001. Holotipo y paratipo depositados en Q­ CAZ. Localidad tipo. ECUADOR, Pichincha: Volcán Pasochoa, 35 km al sur de Quito. 6 Umm 29 Descripción. Cabeza de color marrón claro polinoso. Ocelos de color blanco, trián­ gulo ocelar más oscuro que la cabeza. Carina prominente. Una cerda oral. Arista con 4 ramas superiores, 2 ramas inferiores, más la bifurca­ ción terminal y pequeñas ramificaciones inter­ nas. Longitud del cuerpo 3.2mm. Alas claras, longitud 3.6 mm. Índice costal 3.5. Tórax de color marrón claro polinoso con 8 filas de pe­ los acrosticales. Patas amarillentas. Abdomen amarillento, del primero al quinto tergitos pre­ sencia de la línea dorsal media, las bandas marginales son de forma triangular y no llegan a los ángulos laterales del tergito; en el prime­ ro y segundo tergitos, a veces, las bandas pier­ den la forma y son apenas líneas engrosadas; el sexto tergito tiene una mancha en la parte cen­ tral. Terminalia Ó . Placa anal libre. Surestilo con 10-11 dientes primarios, 4 dientes secun­ darios y 8-9 cerdas marginales, 11-13 cerdas en la parte baja del epandrio. Edeago quitini­ zado, en vista lateral se observa que la parte superior tiene una curvatura pronunciada, ex­ crecencias laterales quitinizadas, ápice y zona central superior membranosa, la parte ventral cubierta por una membrana micropubescente, el ápice presenta una invaginación conspicua, abertura dorsal 1/5 de la longitud total; gono­ podio fusionado a las paráfisis y unido al edeago por tejido membranoso. Índice del edeago 0.88. Etimología. "Patacorona" palabra del idioma quichua, que significa meseta, Pataco­ rona es una meseta del Volcán Pasochoa. D. quillu sp. nov. (Figs. 10-13,63) 8 Figs. 6-9.Drosophila patacorona sp. nov. holotipo macho: 6, suresúlo, vista posterior; 7-9, edeago y apoderna del edeago, vísta ventral, dorsal y lateral izquierda respectivamente. Material examinado. Holotipo Ó (disectado y guardado en microtubo), etiquetado D. quillu holoti­ po ÓDVela del. 2003 I Pasochoa, Pichincha Ecuador DVela col., mar. 2001. Cuatro paratipos O(disecta­ dos y guardados en microtubos), etiquetados D. quí­ llu paratipo Ó DVela det. 2003 I Pasochoa, Pichin- Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vol. xxvn Números 1 y 2: 29-41, octubre del 2005 D Vela y Rafael 30 cha-Ecuador DVela col., mar. 2001. Un paratipo (di­ sectado y guardado en microtubo), etiquetado D. qui­ l/u paratipoÓ DVela det. 2003 I Pasochoa, Pichincha, Ecuador DVela col., abr. 2001. Dos paratipos Ó (di­ sectados y guardados en microtubos), etiquetados D. quillu para tipo ? DVela det. 2003 I Pasochoa, Pichin­ cha, Ecuador DVela col., jun. 2001. Dos paratipos (disectados y guardados en microtubos), etiquetados D. quillu paratipo Ó DVela det. 2003 I Pasochoa, Pi­ chincha, Ecuador DVela col., jul. 2001. Ocho parati­ pos Ó (disectados y guardados en microtubos), eti­ quetados D. quillu paratipoÓDVela det. 2003 I Paso­ choa, Pichincha, Ecuador DVela col., abr. 2002. Ho­ lotipo y paratipos depositados en QCAZ. Localidad tipo. ECUADOR, Pichincha: Volcán Pasochoa, 35 km al sur de Quito. Descripción. Cabeza de color marrón cla­ ro, ocelos blancos, triángulo ocelar del mismo color que la cabeza. Ojos de color rojo brillan­ te. Orbital media es 1/2 o 1/3 de la primera. Or­ bital media más cerca del ojo. Carina promi­ nente. Una cerda oral prominente. Antena de 10 0.1 mm. o color marrón claro, pubescente. Arista ramifi­ cada, con 5 ramas dorsales, 2 ramas ventrales, más la bifurcación terminal y pequeñas ramifi­ caciones internas. Longitud del cuerpo 3.2 mm. Tórax de color marrón claro polinoso, pelos acrosticales ordenados en siete u ocho filas frente a las cerdas dorsocentrales. Escutelo de color marrón claro polinoso. Patas de color ma­ rrón claro ligeramente amarillentas. Alas cla­ ras, longitud 3.7 mm. Índice costal 4.8. Abdo­ men amarillo pálido, presencia esporádica de línea dorsal media del primero al cuarto tergi­ tos. El quinto y sexto tergitos presentan una banda oscura en el centro la cual no llega al borde superior. Terminalia Ó. Placa anal libre. Surestilo con una fila de 13 dientes primarios, 2-5 dien­ tes secundarios y 7-17 cerdas marginales, 2-6 cerdas en la parte baja y 1-2 cerdas en la parte media del epandrio. Edeago quitinizado, rama ventral de forma triangular, en vista lateral se observa la forma redondeada del edeago que en la parte central presenta una zona más quitini­ zada y de borde aserrado, abertura dorsal me­ nos de 1/2 de la longitud del edeago; apodema ancho y menos quitinizado en la base. Índice del edeago 1.27 (1.12). Etimología. "Quillu" palabra del idioma quichua que significa amarillo. Esta especie presenta coloración amarillenta en el abdomen y tórax. D. ninarumi sp. nov. (Figs. 14-17,64) 11 Figs. 10-13. Drosophila. quillu sp. nov. holotipo macho: lO, surestilo, vista posterior, lado izquierdo omitido; 11-13, edeago y apodema del edeago, vista ventral, dorsal y lateral izquierda respectivamente. Material examinado. Holotipo Ó (disectado y guardado en microtuOO) etiquetado D. ninarumi holotipo DVela det. 2003 I Pasochoa, Pichincha, Ecuador DVela col., abr. 2001. Un paratipo Ó, (disectado y guardado en microtubo), etiquetado D. ninarumi paratipoODVela det. 2003 I Pasochoa, Pichincha, Ecuador DVela col., jun. 2001. Dos paratiposÓ(disectados y guardadps en micro­ tubos), etiquetados D. ninarumi paratipo O DVela det. 2003 I Pasochoa, Pichincha, Ecuador DVela col., jul. 2001. Un paratipo Ó (disectado y guardado en microtu­ 00), etiquetado D. ninarumi paratipo Ó DVela det. 2003 I Pasochoa, Pichincha, Ecuador DVela col., ene. 2002. Dos paratiposÓ(disectados y guardados en microtubos), Revista Ecuatoriana de Medicina y Ciencias Biológicas Vol. XXVII Números 1 y 2: 30-41, octubre del200j Catorce nuevas especies del género Drosophila (Diptera, Drosophilidae) en el Bosque húmedo montano del Volcán....... etiquetados D. ninarumi paratipo Ó DVela det. 2003 I Pasochoa. Pichincha, Ecuador DVela col., febo 2002. Holotípo y paratipos depositados en QCAZ. Localidad tipo. ECUADOR, Pichincha: Volcán Pasochoa, 35 km. al sur de Quito. 0.1 mm o 15 31 las frente a las cerdas dorsocentrales. Escutelo de color marrón claro polinoso. Patas de color marrón amarillento. Alas claras, longitud 3.2rnm. Índice costal 3.1. Abdomen amarillen­ to, del primero al sexto tergitos presenta la lí­ nea dorsal media; el último tergito presenta la mitad inferior pigmentado, visto lateralmente los tergitos son obscuros y presentan áreas cla­ ras irregulares. Terminalia Ó. Placa anal libre. Surestilo con una fila de 9-14 dientes primarios, sin dientes secundarios, 12-22 cerdas marginales y 2-4 cerdas en la parte baja y 1-2 cerdas en la parte media del epandrio. Edeago quitinizado y delgado, ápice semimembranoso, en vista ven­ tral la cabeza tiene forma triangular con los bordes semiaserrados y el ápice membranoso y levemente bifurcado, abertura dorsal 1/4 de la longitud total; el apodema presenta una curva­ tura hacia la parte ventral; gonopodio desnudo. índice del edeago 0.82 (1.1). Relacionada con D. cuaso y D. tríangulina. Etimología. "Ninarumi" palabra del idio­ ma quichua, que significa piedra incandescen­ te, nombre de una pequeña loma cercana al Vol­ cán Pasochoa. 18 Figs. 14-17. Drosophila ninarumi sp. nov. holotipo macho: 14, sures tilo, vista posterior; 15-17, edeago y apodema del edeago, vista ventral, dorsal y lateral izquierda respectivamente. Descripción. Cabeza de color marrón cla­ ro, ligeramente polinosa. Ocelos transparentes, triángulo ocelar ligeramente más oscuro que la cabeza. Ojos de color rojo oscuro. Orbital me­ dia es 1/2 de la primera, todas las cerdas orbi­ tales están a la misma distancia del ojo. Carina prominente, una cerda oral prominente. Antena marrón, ligeramente más oscura que la cabeza. Arista ramificada, con 4 ramas dorsales y 2 ra­ mas ventrales más la bifurcación terminal y pe­ queñas ramificaciones internas. Longitud del cuerpo 3.0rnm. Tórax de color marrón claro po­ linoso, pelos acrosticales ordenados en ocho fi­ D. surucucho sp. nov. (Figs. 18-21,65) Material examinado. Holotipo Ó (disectado y guardado en microtubo), etiquetado D. surucucho holoti­ po Ó DVela det. 2003 I Pasochoa, Pichincha, Ecuador DVela col., abr. 2001. Dos paratipos Ó (disectados y guardados en microtubos), etiquetados D. surucucho pa­ ratipo Ó DVela det. 2003/ Pasochoa, Pichincha, Ecua-' dor DVela col., abr. 2001. Dos paratipos Ó (disectados y guardados en microtubos), etiquetados D. surucucho pa­ ratipo ÓDVela det. 2003 / Pasochoa, Pichincha, Ecuador DVela col., mayo 2001. Dos paratipos Ó (disectados y guardados en microtubos), etiquetados D. surucucho pa­ ratipoÓ DVela det. 2003 / Pasochoa, Pichincha, Ecuador DVela col., jun. 2001. Tres paratipos Ó (disectados y guardados en microtubos), etiquetados D. surucucho pa­ ratipo ÓDVela det. 2003/ Pasochoa, Pichincha, Ecuador DVela col., 1ul. 2001. Holotipo y paratipos depositados en QCAZ. Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vol. XXVII Números l y 2: 31-41, octubre del 2005 l Vela y Rafael 32 18 0.1 mm Terminalia Ó, Placa anal libre. Surestilo con 9 dientes primarios, 10 dientes secunda­ rios, 11-14 cerdas marginales, 8-9 cerdas en la parte inferior, 4 cerdas en la parte media del epandrio. Edeago quitinizado, alargado, ápice con dos proyecciones muy quitinizadas, la par­ te ventral membranosa y micropubescente; go­ nopodio semimembranoso con dos pequeñas cerdas en la parte distal, unido al edeago por te­ jido membranoso. Índice del edeago 0.96. Etimología. "Surucucho" palabra del idioma quichua, que significa lugar o rincón donde crece el suro, planta del género Chuquea. Surucucho es una loma cercana al Pasochoa. 19 Figs. 18-21. Drosophila. surucucho sp. nov. holotipo macho: 18, surestilo, vista posterior, lado izquierdo omitido; 19-21, edeago, apodema del edeago y paráfisis, vista ventral, dorsal y lateral izquierda respectivamente. Localidad tipo. ECUADOR, Pichincha: Volcán Pasochoa, 35 km. al sur de Quito. Descripción. Cabeza de color marrón cla­ ro polinoso. Ocelos de color miel, triángulo ocelar de igual color que cabeza. Ojos rojos. Segunda orbital 1/3 de la primera, todas las cer­ das orbitales a igual distancia del ojo. Carina muy prominente. Dos cerdas orales. Antena de color marrón claro, el tercer segmento más os­ curo, Arista con 5 ramas superiores, 2 ramas in­ feriores más la bifurcación terminal y pequeñas ramificaciones internas. Longitud del cuerpo 3.0mm. Tórax marrón claro polinoso con 8 fi­ las de pelos acrosticales. Patas amarillentas. Alas claras, longitud 3.0rnm. Índice costal 4.1. Abdomen amarillento, del primero al quinto tergitos presencia de línea dorsal media divi­ diendo las bandas marginales que son de forma triangular, en algunas ocasiones estas son muy delgadas y con apariencia de líneas verticales y en otros son triángulos que se extienden hacia los ángulos laterales, el sexto tergito tiene una franja en el centro. D. ichubamba sp. nov. (Figs. 22-25, 66) Material examinado. Holotip Ó (disectado y guardado en microtubo), etiquetado D. iehubamba holotipo Ó OVela det. 2003 / Pasochoa, Pic)lincha, Ecuador OVela col., mayo 2001. Un paratipoÓ (disec­ tado y guardado en microtubos), etiquetado D. iehu­ bamba paratipo Ó OVela del. 2003 / Pasochoa, Pi­ chincha, Ecuador OVela col., mayo 2001. Oos parati­ posc:Xdisectados y guardados en microtubos), etique­ tados D. ichubamba paratipo Ó OVela del. 2003 / Pa­ sochoa, Pichincha, Ecuador OVela col., abr. 2002. Holotipo y paratipos depositados en QCAZ. 22 0.1 mm Figs. 22-25. Drosophila. ichubamba sp. nov. holotipo macho: 22, surestilo, vista posterior; 23-25, edeago y apodema del edea­ go, vista ventral, dorsal y lateral izquierda respectivamente. Revista Ecuatoriana de Medicina y Ciencias Biológicas· Vol. XXVII Números 1 y 2: 32·41, octubre de12005 D Catorce nuevas especies del género Drosophila (Diptera, Drosophilidae) en el Bosque húmedo montano del Volcán....... D. ureu sp. nov. Localidad tipo. ECUADOR, Pichincha: Volcán Pasochoa, 35 km. al sur de Quito. Descripción. Cabeza de color marrón claro polinoso. Ocelos de color amarillento, con los bordes ligeramente más oscuros. Triángulo ocelar del mismo color que la cabe­ za. Ojos de color rojo. Orbital media es 1/2 de la primera, todas a la misma distancia del ojo. Carina prominente. Una cerda oral prominen­ te. Antena de color marrón claro. Arista con 5 ramas superiores y 2 ramas inferiores más la bifurcación terminal y pequeñas ramificacio­ nes internas. Longitud del cuerpo 2.9 mm. Tó­ rax de color marrón claro polinoso, con 8 filas de pelos acrosticales. Escutelo del mismo co­ lor que el tórax, patas de color amarillento li­ geramente polinoso. Alas claras, longitud 3.0mm. Índice costal 3.7. Abdomen amarillen­ to, del primero al cuarto tergitos con una del­ gada banda oscura en el borde inferior; la lí­ nea dorsal media se presenta irregularmente en uno o varios tergitos dividiendo las bandas marginales; en el quinto y sexto tergitos se presenta una banda que cubre la mitad inferior del mismo. (Figs. 26-29, 67) Material examinado. Holotipo (disectado y guar­ dado en microtubo), etiquetado D. urcu holotipo DVe­ la det. 2003 I Pasochoa, Pichincha, Ecuador DVela col., abr. 2002, depositado en QCAZ. Localidad tipo. ECUADOR, Pichincha: Volcán Pasochoa, 35 km. al sur de Quito. Terminalia Ó. Placa anal libre, termina en una prolongación hacia el surestilo. Surestilo con 11-12 dientes primarios, 25-27 dientes se­ cundarios en la parte externa del sures tilo y 9­ 10 dientes en la parte interna del surestilo. 4-5 cerdas marginales, 3-4 cerdas en la parte baja del epandrio. Edeago quitinizado con una pro­ tuberancia quitinizada en la parte ventral, y la parte apical dorsal membranosa, con una hen­ didura muy pronunciada, la parte ventral recu­ bierta por una membrana micropubescente. Ín­ dice del edeago 1.16. Terminalia Ó. Placa anal libre. Epandrio con micropubescencia. Surestilo con una fila de 8-10 dientes primarios, sin dientes secunda­ rios, 8-10 cerdas marginales, 2-3 cerdas en la parte baja del epandrio y una cerda en la parte media. Edeago quitinizado, rama ventral de forma triangular, la parte superior del edeago es de forma redonda, la parte dorsal semimembra­ nosa, en vista dorsal presenta una área transpa­ rente de forma circular; gonopodio con una cer­ da pequeña en la parte central y unido al edea­ go por tejido membranoso. Índice del edeago 1.22. Etimología. "Ichubamba" palabra qui­ chua, que significa pajonal. Ichubamba es un cerro junto al Volcán Pasochoa. 33 0.1 mm 27 Figs. 26-29. Drosophila urcu sp. nov. holotipo macho: 26, su­ restilo, vista posterior, lado izquierdo omitido; 27-29, edeago y apodema del edeago, vista ventral, dorsal y lateral izquierda respectivamente. Revista Ecuatoriana de Medicina y Ciencias Biológicas Vol. xxvn Números 1 y 2: 33-41, octubre del 2005 34 Vela y Rafael 0.1 mm 32. I 0.1 mm Figs. 30-31. Drosophila suni sp. nov. holotipo macho: 30, su­ restilo, vista posterior; 31, hipandrio, vista posterior. Etimología. "Urcu" palabra del idioma quichua que significa cerro, nombre de un cerro ubicado cerca al Volcán Pasochoa. Grupo flavopilosa D. suni sp. nov. (Figs. 30-34, 68) Material examinado. Holotipo (disectado y guar­ dado en microtubo), etiquetado D. suni holotipo DVela det. 2003/ Pasochoa, Pichincha, Ecuador DVela col., mar. 2001, depositado en QCAZ. Localidad tipo. ECUADOR, Pichincha: Volcán Pasochoa, 35 km. al sur de Quito. Terrninaliad. Placa anal fusionada. Epan­ drio pubescente. Surestilo micropubescente con 7-8 dientes primarios, sin dientes secunda­ rios, 20-22 cerdas marginales, en la parte baja del epandrio numerosas cerdas muy largas. Edeago quitinizado, largo, ápice con bordes aserrados y una hendidura en el centro; apode­ ma semiquitinizado y muy corto; gonopodio ovalado y alargado, con una cerda larga en la parte distal interna, unido al edeago por tejido membranoso. Índice del edeago 6.5. Etimología. "Suni" palabra quichua, que significa largo, característica que presenta el falo de esta especie. Figs. 32-34. Drosophila suni sp. nov. holotipo macho: 32-34, edeago, apodema del edeago y paráfisis, vista ventral, dorsal y lateral izquierda respectivamente. D. taxohuaycu sp. nov. (Figs. 35-39,69) Material examinado. Holotipo Ó (disectado y guardado en microtubo), etiquetado D. taxohuaycu holo­ tipoÓDVela det. 2003 / Pasochoa, Pichincha, Ecuador DVela col., mar. 2001. Un paratipo Ó (disectado y guar­ dado en microtubo), etiquetado D. taxohuaycu paratipo DVela det. 2oo3/Pasochoa, Pichincha-Ecuador DVela col., may.200l. Holotipo y paratipo depositados en Q­ CAZ. Localidad tipo. ECUADOR, Pichincha: Volcán Pasochoa, 35 km. al sur de Quito. Descripción. Cabeza de color marrón os­ curo. Ocelos blancos, triángulo ocelar de igual color que cabeza. Ojos rojos. Carina pronuncia­ da, una cerda oral prominente. Tórax de color marrón oscuro polinoso. Patas de color amari­ llento. Alas transparentes. Revista Ecuatoriana de Medicina y Ciencias Biológicas Vol. XXVII Números 1 y 2: 34-41, octubre del 2005 D Catorce nuevas especies del género Drosophila (Diptera, Drosophilidae) en el Bosque húmedo montano del Volcán....... Tenninalia Ó. Placa anal libre. Epandrio micropubescente. Surestilo micropubescente, con 13 dientes primarios, sin dientes secunda­ rios, 11 cerdas marginales y 18 cerdas en la par­ te baja del epandrio. Hipandrio de forma de triángulo invertido, presenta dos cerdas en las paráfisis. Edeago quitinizado, ápice de forma triangular y asimétrico, un lado es más corto, curvado hacia la parte ventral, la región supe­ rior dorsal presenta prominencias de las que emergen microproyecciones, rama ventral lar­ ga; apodema ligeramente más qUÍtinizado y más corto que el edeago; gonopodio de forma triangular con tres cerdas en la parte distal. Ín­ dice del edeago 2.07. Etimología. "Taxohuaycu" palabra qui­ chua que significa quebrada del taxo, fruto del género Passiflora. 35 D. sisa sp. nov. (Figs. 40-44, 70) Material examinado. Holotipo Ó<disectado) guar­ dado en microtubo), etiquetado D. sisa holotipo O DVela det. 2003 I Pasochoa, Pichincha, Ecuador DVela coL, abr. 2002. Un paratipo (disectado y guardado en microtubo), etiquetado D. sisa paratipo ÓDVela del. 2003 I Pasochoa, Pichincha-Ecuador DVela col., abr. 2002. Holotipo y para­ tipo depositados en QCAZ. Localidad tipo. ECUADOR, Pichincha: Volcán Pasochoa, 35 km. al sur de Quito. Descripción. Cabeza de color marrón bri­ llante, ocelos blancos, triángulo ocelar ligera­ mente más oscuro que la cabeza. Primera orbi­ tal 1/3 de la primera, segunda y tercera orbital ligeramente más cerca del ojo. Carina promi­ nente, no surcada, una cerda oral. Arista con 3 ramas superiores, 2 inferiores más la bifurca­ ción tenninal. Longitud del cuerpo 3.0 mm. Tó­ rax marrón polinoso, con 8 filas de pelos acros- I 0.1 mm 6.1 mm 'il 38 Figs. 35-39. Drosophila taxohuaycu sp. nov. holotipo macho: 35, surestilo, vista posterior; 36, hipandrio, vista posterior; 3739, edeago, apodema del edeago y paráfisis, vista ventral, dor­ sal y lateral izquierda respectivamente. Figs. 40-44. Drosophila sisa sp. nov. holotipo macho: 40, su­ restilo, vista posterior; 41, hipandrio, vista posterior; 42-44, edeago, apodema del edeago y paráfisis, vista ventral, dorsal y lateral izquierda respectivamente. Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vol. XXVII Números 1 y 2: 35-41, octubre del 2005 36 Vela y Rafael ticales. Patas de color marrón claro. Punta del ala oscurecida, la sombra cubre la parte distal de las celdas marginal, submarginal, primera posterior y segunda posterior. Longitud de las alas 3.1 mm. Índice costal 3.8. Abdomen ama­ rillento, del primero al cuarto tergitos presencia de línea dorsal media que divide las bandas marginales, el quinto y sexto tergitos totalmen­ te pigmentados. Terminalia Ó . Placa anal libre. Surestilo con 8 dientes primarios, 5 dientes secundarios, 4-5 cerdas marginales, 15 dientes quitinizados y redondeados en el epandrio inferior, una cer­ da pequeña ubicada hacia la parte terminal y 8 cerdas grandes, 2 cerdas en la parte media del epandrio. Edeago largo y quitinizado, la parte superior ancha y curvada hacia la parte dorsal, los bordes de la región superior se doblan hasta juntarse en el parte ventral, ápice con dos pro­ yecciones laterales y más abajo del ápice se presentan cuatro proyecciones quitinizadas for­ mando una especie de corona, la rama ventral del edeago es filiforme y termina en dos forma­ ciones a manera de dedos; apodema membra­ noso y muy corto. Índice del edeago 7.58. Etimología. "Sisa" palabra quichua que significa flor. El ápice del edeago de esta espe­ cie se asemeja a una flor. ja del epandrio y 1 cerda en la parte media. Edeago quitinizado, ensanchado y membrano­ so en la parte dorsal, rama ventral grande, el ápice con dos proyecciones quitinizadas y jun­ to a ellas dos proyecciones membranosas y mi­ cropubescentes; apodema semiquitinizado; go­ nopodio fusionado a las paráfisis con dos cer­ das pequeñas y una larga y gruesa. Índice del edeago 1.47. 45 0.1 mm Figs. 45-48. Drosophila cuscungu sp. nov. holotipo macho: 45, surestilo, vista posterior; 46-48, edeago y apodema del edeago, vista ventral, dorsal y lateral izquierda respectivamente. Grupo guaraní D. cuscungu sp. nov. (Figs. 45-48, 71) Material examinado. Holotipo Ó (disectado y guardado en microtubo), etiquetado D. cuscungu holoti­ po OVela det. 2003/ Pasochoa, Pichincha, Ecuador OVela col.. jun. 2001, depositado en QCAZ. Localidad tipo. ECUADOR, Pichincha: Volcán Pasochoa, 35 km. al sur de Quito. Terminaliaó . Placa anal libre, epandrio superior micropubescente. Surestilo con 10-11 dientes primarios, 3-4 dientes en la parte exter­ na y 1-4 dientes en la parte interna del surestilo. 9-10 cerdas marginales, 8 cerdas en la parte ba­ Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vol. Etimología. "Cuscungu" palabra del idio­ ma quichua con que se nombra a los buhos de plumaje negro. Esta especie presenta colora­ ción oscura. Grupo annuümana D. yana sp. nov. (Figs. 49-52, 72) Material examinado. HoJotipo Ó (disectado y guardado en microtubo), etiquetado D. yana holotipo OVeJa del. 2003/ Pasochoa, Pichincha, Ecuador OVela col., mar. 2001. Un paratipoÓ (disectado y Buardado en microtubo), etiquetado D. yana paratipo Ó OVela del. 2003/ Pasochoa, Pichincha, Ecuador OVeJa col., mayo 2001. Un paratipo Ó (dísectado y guardado en microtu- xxvn Números l y 2: 36-41, octubre del 2005 Catorce nuevas especies del género Drosophila (Diptera, Drosophilidae) en el Bosque húmedo montano del Volcán....... 37 bo), etiquetado D. yana paratipoÓOVela det. 2003/ Pa­ sochoa, Pichincha-Ecuador OVela col., nov. 2001. Holo­ tipo y paratipos depositados en QCAZ. tergito con una banda vertical delgada en el centro, esta banda se adelgaza en los últimos tergitos. Localidad tipo. ECUADOR, Pichincha: Volcán Pasochoa, 35 km al sur de Quito. Terminaliad. Placa anal fusionada. Epan­ 000 pubescente. Surestilo con 10-11 dientes primarios, sin dientes secundarios, 9-10 cerdas marginales, 16-17 cerdas en la parte baja del epanOOo 1-2 cerdas en la parte media y 1-2 cer­ das en la parte superior del epanOOo. Edeago quitinizado con dos proyecciones laterales en la parte dorsal, ápice delgado y bífido; apodema delgado con una pequeña cerda y micropubes­ cencia en la parte central; gonopodio semiova­ lado y unido al edeago por tejido membranoso. Índice del edeago 1.14. Descripción. Cabeza de color marrón os­ curo. Ocelos blancos, triángulo ocelar marrón oscuro. Ojos rojo oscuro. Segunda cerda orbital 213 de la primera y ligeramente más cerca del ojo. Carina prominente y surcada. Una cerda o­ ral. Arista con 5 ramas superiores, 3 ramas infe­ riores más la bifurcación terminal y pequeñas ramificaciones internas. Longitud del cuerpo 3.9mm. Tórax marrón oscuro polinoso con franjas fusionadas que dejan pequeños espacios más claros y más polinosos, ocho filas de pelos acrosticales. Patas de color marrón oscuro. Alas con manchas en r-m (vena transversal anteror) y dm-Cu (vena transversal posterior), las celdas marginal, submarginal y primera posterior, también están sombreadas, longitud 4.1mm. Ín­ dice costal 3.5. Abdomen marrón oscuro, cada Etimología. "Yana" palabra del idioma quichua para nombrar cualquier cosa de colora­ ción negra. Esta especie es de coloración casi negro. Especies no agrupadas D. condormtlChay sp. nov. (Figs. 53-56, 73) 0.1 mm CJ Material examinado. Holotipo Ó (disectado y guardado en microtubo), etiquetado D. condormachay holotipo OVela del. 2003 / Pasochoa, Pichincha, Ecua­ dor OVela col., jun. 2001. Un paratipo Ó (disectado y guardado en microtubo). etiquetado D. condormachay paratipo OVela del. 2003 / Pasochoa, Pichincha, Ecua­ dor OVela col., ago. 2001. Oos paratiposÓ(disectados y guardados en microtubos), etiquetados D. condorma­ chay paratipoÓ OVela det. 2003 / Pasochoa, Pichincha­ Ecuador OVela col., sep. 2001. Oos paratiposÓ(disecta­ dos y guardados en microtubos), etiquetados D. condor­ machay paratipo Ó OVela del. 2003 / Pasochoa, Pichin­ cha-Ecuador OVela col., oct. 2001. Holotipo y paratipos depositados en QCAZ. Localidad tipo. ECUADOR, Pichincha: Volcán Pasochoa, 35 km al sur de Quito. 52 Figs. 49-52. Drosophila yana sp. nov. holotipo macho: 49, su­ restilo, vista posterior; 50-52, edeago, apodema del edeago y paráfisis, vista ventral, dorsal y lateral izquierda respectiva­ mente. Descripción. Cabeza de color marrón cla­ ro polinoso. Ocelos transparentes, triángulo ocelar del mismo color que cabeza. Ojos rojos. Segunda orbital 2/3 de la primera. Carina pro­ minente y surcada, una cerda oral prominente. Antena micropubescente. Tórax marrón claro polinoso, cerdas preescutelares. Patas amari- Revista Ecuatoriana de Medicina y Ciencias Biológicas· Vol. xxvn Números l y 2: 37-41, octubre del 2005 Vela y Rafael 38 llentas. Alas con manchas en r-m (vena trans­ versal anterior) y dm-Cu (vena transversal pos­ terior), las celdas marginales, submarginal y primera posterior bastante sombreadas. Abdo­ men amarillo grisáceo, bandas oscuras en la lí­ nea media dorsal y en los bordes superior e in­ ferior de cada tergito. Etimología. Palabra del idioma quichua, que significa cueva del cóndor. "Condor Ma­ chay" es nombre de un peñasco cercano al Vol­ cán Pasochoa. TerminaliaÓ. Placa anal fusionada. Epan­ drio microopubescente. Surestilo micropubes­ cente, con 12-13 dientes primarios, sin dientes secundarios, 7-8 cerdas marginales, 28 cerdas en la parte baja del epandrio y 3 en la parte su­ perior. Edeago quitinizado, con dos pequeñas proyecciones quitinizadas en la parte superior dorsal, ápice con una ligera hendidura; gonopo­ dio de forma semicircular con una cerda en la parte distal. Índice del edeago 1.05. Material examinado. hOlotipoÓ(disectado y guar­ dado en microtubo), etiquetado D. pugyu holotipo DVe­ la det. 2003 I Pasochoa, Pichincha, Ecuador DVela col., oct. 2001 depositado en QCAZ. D. pugyu sp. nov. (Figs. 57-60, 74) Localidad tipo. ECUADOR, Pichincha: Volcán Pasochoa, 35 km al sur de Quito. Terminalia Ó . Placa anal fusionada. Su­ restilo con15-16 dientes primarios, sin dientes secundarios, 11 cerdas marginales, en la parte baja del epandrio 2 cerdas pequeñas y 8 cerdas grandes, epandrio pubescente. Edeago delga­ do y quinitizado con dos pequeñas proyeccio­ 53 57 0.1 mm 0.1 mm o J 54 Figs. 53-56. Drosophila condonnachay sp. nov. holotipo ma­ cho: 53, surestilo, vista posterior; 54-56, edeago, apodema del edeago y paráfisis, vista ventral, dorsal y lateral izquierda res­ pectivamente. 58 Figs. 57-60. Drosophila pugyu sp. nov. holotipo macho: 57, su­ restilo, vista posterior; 58-60, edeago, apodema del edeago y paráfisis, vista ventral, dorsal y lateral izquierda respectiva­ mente. Revista Ecuatoriana de Medicina y Ciencias Biológicas - VoL XXVII Números 1 y 2: 38-41, octubre del 2005 Catorce nuevas especies del género Drosophila (Díptera, Drosophilidae) en el Bosque húmedo montano del Volcán....... nes quitinizadas en la parte dorsal, rama ven­ trallarga casi de igual longitud que el gonopo­ dio; gonopodio desnudo, Índice del edeago 1. Etimología. "Pugyu" palabra del idioma quichua, que significa fuente de agua, o manan­ tial. Agradecimientos. A nuestros colegas Anita Mafia y Geovani Onore por sus críticas y sugerencias, A IDEA WILD por la donación del material entomólogico. Este trabajo es par­ te del proyecto "Biodiversidad del género Dro­ sophila y detección del impacto ambiental en el Refugio de vida Silvestre Pasochoa" y "Carac­ terización de nuevas especies ecuatorianas de Drosophila" financiado por la Pontificia Uni­ versidad Católica del Ecuador. 39 REFERENCIAS L- VELA, D. YRAFAEL, V. Ocho nuevas especies del grupo tri­ punctata, género Drosophila (Diptera, Drosophilidae) y el registro de D. paraguayensis en el Bosque Protector Paso­ choa, Pichincha-Ecuador. Revista de la Pontificia Uni­ versidad Católica del Ecuador, Quito, 2001; 66:91-120. 2.- VELA, D. YRAFAEL, V. Estudios sobre la diversidad del gé­ nero Drosophila (Díptera, Drosophílidae) en el Bosque Pa­ sochoa de la Provincia de Pichincha-Ecuador. Revista de la Pontificia Universidad Católica del Ecuador, Quito, 2003; 71:117-127. 3.- VELA, D. & RAFAEL, V. Three new Andean specíes of Drosophila (Diptera, Drosophilidae) of the mesophragma­ tica group. Iheringia, Sér. Zool., Porto Alegre, 2004; 94(3):295-299. 4.- VELA, D. Y RAFAEL, V. Dos nuevas especies del grupo flavopilosa, género Drosophila (Díptera, Drosophilídae) en el Bosque Pasochoa. Província de Pichincha. Revista Ecuatoriana de Medicina y Ciencias Biológicas, de la Casa de la Cultura Ecuatoriana, 2004; 26(1,2):7-13. 5.- VELA, D. Y RAFAEL, V. Tres nuevas especies del grupo guarani, género Drosophila (Díptera, Drosophilídae) en el Bosque Pasochoa, Provincia de Pichincha. Revista Ecua­ toriana de Medicina y Ciencias Biológicas, de la Casa de la Cultura Ecuatoriana, 2004; 26 (1,2): 14-21. 6.- VELA, D. Y RAFAEL, V. Nuevas especies de Drosophila (Diptera, Drosophilidae) en el Bosque Pasochoa, Pichin­ cha-Ecuador. Revista de la Pontificia Universidad Cató­ lica del Ecuador, Quito, 2005, 75: 69-80. Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vol. XXVII Números 1 y 2: 39-41, octubre del 2005 Vela y Rafael 40 64 63 62 67 D I 0.1 DDD 68 Figs. 61-68. Edeago y apodema del edeago, vista lateral izquierda: 61, Drosophila shuyu sp. nov.; 62, Drosophila pataeorona sp. nov.; 63, Drosophila quillu sp. nov.; 64, Drosophila ninarumi sp. nov.; 65, Drosophila surueueho sp. nov.; 66, Drosophila iehu­ bamba sp. nov.; 67, Drosophila ureu sp. nov.; 68, Drosophila suni sp. nov. Revista Ecuatoriana de Medicina y Ciencias Biológicas Vol. XXVII Números 1 y 2: 40-41. octubre del 2005 Catorce nuevas especíes del género Drosophíla (Díptera, Drosophilidae) en el Bosque húmedo montano del Volcán....... 41 0.1 mm e 73 Figs. 69-74. Edeago y apodema del edeago, vista Ia.teral izquierda: &J, DrorophiÚl taxohuaycu sp. nov.; 70, DroSOphiÚl siso sp. nov.; 71, Drosophi!a cuscungu sp. nov.; 72, DrosophiÚl yana sp. nov.; 73, Drosophila condorrrtilChay sp. nov. y 74, Drosophila pugyu sp. nov. Revista Ecuatoriana de Medicina Y Ciencias Biológicas - Vol. XXVII Números 1 y 2: 4141, octubre dcl2005 D ............­ D Identificación de islas CpG en el genoma humano a través de las cadenas de Markov: Un modelo matemático basado en probabilidades Quishpe Evelyn2, Sánchez María Eugenia!, Oleas-De la Carrera Gabriela3, Paz-y-Miño César! 'Laboratorio de Genética Molecular y Citogenética Humana-Pontificia Universidad Católica del Ecuador 2 Escuela Politécnica Nacional-Facultad de Ciencias-Ingeniería Matemática 3 Escuela Politécnica del Ejército -Facultad de Ciencias Aplicadas- Ingeniería en Biotecnología [email protected] , [email protected] ' Recibido l abril 2005, aprobado 15 junio 2005 RESUMEN. Los genes de importancia biológica conocidos como genes esenciales de mantenimiento o housekeeping genes sue­ len encontrarse rodeados por regiones denominadas Islas CpG, llamadas así debido a que contienen una cantidad mucho mayor de dinueleótidos CpG que el resto del genoma y, ya que del reconocimiento de tales islas se puede inferir la ubicación de los house­ keeping genes, un modelo matemático de identificación de Islas CpG facilitaría su diferenciación del resto del genoma. El mode­ lo matemático que se presenta en este artículo toma como ejemplo a una secuencia de 60 nuc1eótidos presente en el genoma del parvovirus canino y se basa en las cadenas de Markov para calcular la probabilidad de que un fragmento de dicha secuencia, en re­ lación al resto de ella, corresponda o no a una Isla CpG. Este modelo puede ser utilizado en cualquier secuencia, independiente­ mente de su número de nucleótidos sin embargo, la del parvovirus, escogida en este caso como una pequeña muestra de ejempli­ ficación. sirvió para comparar y confirmar por simple inspección los resultados. PALABRAS CLAVE. Cadenas de Markov, Islas CpG, Hosekeeping genes. ABSTRACT. The biologically important genes known as essential genes or housekeeping genes are usually found surrounded by regions called "CpG Isles" . The CpG isles are named so because they contain a much larger quantity of dionucleotides CpG than the rest of the genome. Since during the recognition of such isles the location of the housekeeping genes can be inferred, a mathe­ matical model of identification of CpG isles will make it easier to teH it apart from the rest of the genome. The mathematical model that is presented in this artiele uses as an example a sequence of 60 nuc1eotides present in the genome of the canine paro virus and is based on the Markov chains to calculate the probability that a fragment of this sequence. in relationship to the rest of it, corresponds or not to a CpG isle. This model can be used in any sequence, independently from its number of nucleotides. However the parovirus sequence, chosen in this case as a small sample. served to compare and confirm the results by simple inspection. KEYWORDS. Markov chains. CpG isles. Housekeeping genes. INTRODUCCIÓN La importancia del estudio de los house­ keeping genes radica en que de su expresión dependen funciones esenciales para la vida de la célula. Sin embargo, y más allá de lo com­ plejo que podría llegar a ser en sí mismo el es­ tudio del gen de nuestro interés, el simple he­ cho de identificar la región en la que se encuen­ tran los housekeeping genes ya implica un gran avance debido a que, de la vastísima extensión del genoma de los eucariontes apenas una pe­ queña proporción corresponde a regiones codi­ ficantes. Entre las herramientas que podemos usar para identificar las regiones en las que se en­ cuentran los housekeeping genes están las Islas CpG. Las Islas CpG son regiones que poseen altas concentraciones de pares de bases CpG y, a menudo, rodean los promotores de los genes expresados constitutivamente, aunque también se encuentran en los promotores de los genes regulados (1). Las Islas CpG son un punto de partida pa­ ra la localización de housekeeping genes; iden­ tificarlas de una forma rápida y precisa, se lo podría hacer con la utilización de modelos ma­ temáticos. Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vol. XXVII Números 1 y 2: 43-46, octubre del 2005 D Quíshpe et al. 44 El presente trabajo relaciona ambos ám­ bitos, las Islas CpG y los modelos matemáticos a partir de que una cadena de Markov definida como una serie de eventos, en la cual la proba­ bilidad de que ocurra un evento depende del evento inmediato anterior (2). Relacionando esta definición con el hecho de que la transición de una base a otra puede considerarse como un evento aleatorio es posible buscar las probabili­ dades de que un fragmento corresponda o no a una Isla CpG (Figura 1). El principio es simple: Dado un nuc1eótido C, cuál será la probabili­ dad de encontrar un nucleótido G a continua­ ción de éste, es decir, la probabilidad buscada dependerá únicamente del estado anterior (C), situación perfectamente compatible con la defi­ nición de la cadena de Markov y aplicable no únicamente a un doblete de bases sino a toda u­ na secuencia. Dicha secuencia fue dividida arbitraria­ mente en dos fragmentos: Fragmento -: ATTCTTTAGAAC CAACTGACCAAGTTCACG Fragmento +: TACGTATGACGTGAT GACCCGCTGCGCGCG Para facilitar la representación de las fór­ mulas, el signo + fue aplicado a todas las se­ cuencias concernientes a las islas CpG y el sig­ no menos corresponderá a aquellas secuencias involucradas en el resto del genoma. Ya que una cadena de Markov es un pro­ ceso aleatorio en el que donde X es el término que representa a los nu­ c1eótidos de la secuencia, n el número de nu­ cleótidos y S las sucesiones posibles de los mismos; fue posible calcular las probabilida­ des de transición (el paso al azar de un nu­ cleótido a otro), tanto para el fragmento co­ mo para el fragmento + a través de las siguien­ tes fórmulas: pij Figura 1.- Posibles cambios de estados (nucleóti­ dos). B indica el punto de partida, una secuencia puede empezar en cualquiera de los nucleótidos, ya elegido un nucleótido, por ejemplo A, el siguien­ te puede ser una e o T o G o una misma A, ahora si el elegido es e, este se convierte en estado ini­ cial y vuelve a iniciarse el proceso. (3) MATERIALES Y MÉTODOS La secuencia que se presenta a continua­ ción corresponde a 60 nuc1eótidos tomados del genoma del parvo virus canino (3): ATTCTTTAGAACCAACTGAC CAAGTTCACGTACGTATGACG TGATGACCCGCTGCGCGCG donde n'ij es el número de veces que el nuc1eó­ tido i sigue al nucleótido j en el fragmento y es la suma de n'u sobre A,C,G y T, es decir, la suma del número de veces que el nucleótido i sigue al nuc1eótido j sobre todas las cuatro ba­ ses. El asterisco (*) representa + o -, depen­ diendo del caso. Las probabilidades de transición fueron colocadas en matrices de transición a medida que se calculaban: Revista Ecuatoriana de Medicina y Ciencias Biológicas Vol. XXVII Números l y 2: 44-46, octubre del 2005 Identificación de islas CpG a través de las cadenas de Markov: Un modelo matemático basado en probabilidades Cuadro 1. Matriz de transición para el fragmento ­ 45 RESULTADOS Y DISCUSIÓN De los dos fragmentos en los que, arbitra­ riamente, fue divida la secuencia original se ob­ tuvo que el primero de estos fragmentos pre­ Océano A­ C­ T­ U G­ senta una probabilidad del 63% de correspon­ A­ 0,333 0,444 0,111 0,111 1 der a la secuencia ordinaria del ADN y no a u­ 0,375 0,245 0,125 0,25 1 C­ i na isla CpG. Del segundo fragmento se obtuvo 1 I G­ 0,666 o o 0,333 una probabilidad del 85% de que dicho frag­ T­ 0,142 0,285 0,142 0,428 1 mento corresponde a una Isla CpG y no a la se­ cuencia regular del genoma. Tales resultados, I:c 1,516 0,979 0,378 1,122 comparados con los de una simple inspección a (4) ambos fragmentos de la secuencia original de 60 nucleótidos permiten confirmar que, efecti­ Cuadro 2. Matriz de transición vamente, el primer fragmento de la secuencia para el fragmento + usada posee apenas un dinucleótido CG en comparación a los seis que saltan a la vista en el -+Sj+l segundo fragmento. Si bien es cierto, la sección del genoma del parvovirus usada en este artícu­ Océano A­ C­ G­ T­ U lo como modelo de ejemplificación es suma­ 0,6 0,41 1 A­ O O mente pequeña, por este motivo el hecho de 1 C­ O 0,125 0,75 0,125 que la primera parte de la secuencia no sea una 0,25 0,375 0,375 1 G­ O isla CpG y la segunda sí 10 sea puede resultar T­ 0,333 0,666 evidente a simple vista, sin embargo tales con­ O O 1 sideraciones se dificultarían de manera propor­ 0,708 1,1 1,416 0,775 I:c cional al número de nucleótidos de la secuencia (4) y, siendo este modelo perfectamente aplicable a En ambas matrices de transición el sím­ n número de nucleótidos, las probabilidades basadas en este tipo de cálculos proporciona­ bolo!.f es la sumatoria de los valores de las fi­ las, correspondientes a las probabilidades de rían una forma más objetiva de evaluar cuál se­ transición para cada uno de los nucleótidos cuencia puede ser considerada como una Isla CpG y cuál no, en el caso de ser destinado a se­ que, de ser correctas, siempre será 1. El Símbolo!.c corresponde en cambio a cuencias mucho más grandes. la sumatoria de las columnas, cálculo necesario Ahora bien, pese a la enorme ventaja que para determinar si el fragmento corresponde a ofrece la flexibilidad de este modelo, siendo una isla o al resto del genoma, para lo cual se aplicable para una secuencia con cualquier nú­ calculó la probabilidad del fragmento + sobre mero de nucleótidos, presenta en este mismo hecho su desventaja ya que, aunque los cálcu­ el - y viceversa. P (fragmento+ I fragmento - ) = = 0,854655 ~ 85% los son relativamente simples, resultaría más laborioso el realizarlos para una secuencia de IIPij (i,j)eK+ cientos o miles de nucleótidos, sin embargo, P (fragmento - I fragmento +) = =0,629458 ~ 63% siendo en nuestros días tan sencillo preparar un IIPij programa que efectúe operaciones tales como Aquí TI representa el producto de la suma las explicadas anteriormente, tal dificultad se­ de las probabilidades de transición para cada ría solventada a través de la elaboración de un programa a base del modelo matemático des­ columna de la matriz de transición. crito en este artículo. -+Sj+l Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vol. XXVII Números 1 y 2: 45-46, octubre del 2005 D Quishpe et al. 46 REFERENCIAS 1. 2. 3. LEWIN, B. GENES vn Ed. Marbán, Madrid 200 1. página 682. http://www.itlp.edu.mxIpublicaltutoriales/in­ vestoper2/tema43.htm 4. NCBI. Nucleotide sequence and genome or­ ganization of canine parvovirus (Banco de Datos-secuencias de AND). www.ncbi.hlm­ .nih. gov/entreziqueIY.fcgi? KARLIN, S. A First Course in Stochastres Processes. USA. 1975 Academic. Press. Pags. 58-59. o Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vol. xxvn Números 1 y 2: 46-46. octubre del 2005 ---~ ... __ Elementos Alu en el análisis filogenético molecular del infraorden Platyrrhini, Primates Bertha Ludeña, Juan Carlos Escobar, Jean-Christophe Pintaud Laboratorio de Genética Molecular de Eucariotes Pontificia Universidad Católica del Ecuador 17-01-2184 Quito-Ecuador. Recibido 1 abril 2005, aprobado 15 de junio 2005 RESUMEN. Los marcadores moleculares Alu: STKI, VP Y HBGF fueron utilizados en el presente trabajo para analizar la filo­ genia de algunas especies de platirrinos ecuatorianos. Los resultados positivos para la amplificación genómica de estos marcado­ res en las especies estudiadas: Cebus albifrons , Saimiri sciureus, Ateles belzebuth, Ateles fusciceps, Alouatta palliata y Lagoth­ rix lagotricha revelan que eventos derivados de transposición de elementos Alu han tenido relación con la filogenia de los prima­ tes del Nuevo Mundo. El análisis mediante Neighbor-joining de la información obtenida ratifica el posicionamiento de los géneros Alouatta, Ate­ les y Lagothrix en la familia Atelidae y sugiere una estrecha cercanía entre los géneros Cebus y Saimiri apoyando la inclusión de éstos en un cIado. PALABRAS CLAVE. Análisis filogenético, evolución, marcadores genómicos Alu, platirrinos. ABSTRACf. Molecular markers Alu: STKI ,VP and HBGF were applied to the phylogenetic analysis of sorne ecuadorian platyrr­ hines. PCR amplification for all three markers was positive in the analysed species: Cebus albifrons , Saimiri sciureus, Ateles bel­ zebuth, Ateles fusciceps, Allouata palliata and Lagothrix lagotricha . This shows that derived Alu- transpositions originated du­ ring New World monkey evolution. Neighbor- joining analysis of this information supports the positioning of genera Alouatta, Ateles and Lagotricha in Atelidae family and a close affiliatíon between genera Cebus and Saimiri which could be considered as a cIade. KEYWORDS. Phylogenetic analysis, evolution, Alu generic markers, Platyrrhines. INTRODUCCIÓN Existe un consenso general respecto a la monofilia del grupo de los primates antropoi­ deos, es decir que tanto Catarrhini (monos del Viejo Mundo) como Platyrrhini (monos del Nuevo Mundo) descienden de un ancestro co­ mún. Respecto a la aparición de los Monos del Nuevo Mundo, los primeros registros fósiles datan del Deseadánico y Colhuehuagiano en Bolivia y Argentina (edades anteriores al Oli­ goceno). En aquella época Sud América era u­ na isla separada de otros continentes por gran­ des expansiones de océano. Se sugiere que en los límites entre Eoceno y Oligoceno, tuvieron lugar cambios climáticos drásticos afectando el nivel del mar. En este ambiente, las dorsales del sur del océano Atlántico quedaron al descubier­ to formando islas, creando así vías que pudie­ ron haber favorecido la migración de fauna desde Africa hacia el aislado continente de Sud América (1,2). Los monos del Nuevo Mundo así como otros mamíferos tienen relaciones de taxones hermanos con grupos africanos, por tanto la existencia de una relación faunística entre Afri­ ca y Sud América en la transición Eoceno, Oli­ goceno es fuertemente corroborada. (3). Los platirrinos actuales se encuentran asignados a 16 géneros o subgéneros, 12 de los cuales pertenecen a tres grupos monofiléti­ cos: los Pitheciidae/Pitheciinae, los Atelí· daelAtelinae y los Callitrichidae/Callitrichinae. Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vol. XXVII Números 1 y 2: 47-51, octubre del 2005 ._­ 48 Ludeña et al. De acuerdo a la clasificación taxonómica de Goodman et al. (4), estos grupos pueden ser considerados como subfamilias. Respecto a las relaciones filogenéticas de los géneros Cebus, Saimiri, Callicebus y Aotus hasta el momento resulta difícil definirlas. Es importante señalar que existe una fuerte evidencia que indicaría un origen mono­ filético de los primates de América Central y América del Sur, sustentada en el análisis de se­ cuencias de ADN (5, 6, 7,8,9). En el Ecuador han sido descritas 19 espe­ cies de platirrinos (10) representadas por los géneros: Callithrix (Cebuella), Saguinus, Sai­ mirl, Cebus, Aotus, Callicebus, Pithecia, Alouatta, Ateles y Lagothrix. En el presente trabajo se aborda la pro­ blemática filogenética en platirrinos ecuato­ rianos mediante el análisis de los elementos Alu: en el gen de la Tirosína kinasa de Células Madre (STK1), en el intrón 4 del pigmento vi­ sual (VP) y en el precursor del factor del cre­ cimiento similar a EGF de uni6n a la heparina (HBGF). Los mencionados marcadores genó­ micos hacen parte de una familia de elemen­ tos nucleares cortos interdispersos en el geno­ ma de primates (SINES). Los SINES constitu­ yen marcadores cladísticos que revelan sina­ pomorfías moleculares y son útiles para esta­ blecer relaciones de taxa en términos genera­ les (11). El presente análisis de elementos nuclea­ res específicos de primates (SINES) ha sido efectuado en el genoma de individuos de las si­ guientes especies: Cebus albifrons, Saimiri sciureus, Ateles belzebuth, Ateles fusciceps, Alouatta palliata y Lagothrix lagotricha MATERIALES Y MÉTODOS Muestras biológicas El ADN para este estudio ha sido extraí­ do a partir de sangre venosa periférica (alrede­ dor de lml), tomada de individuos en cautive­ rio o cazados por indígenas. De la sangre obtenida se ha procedido a realizar la extracción de DNA mediante shock hipotónico y digestión en presencia de protei­ nasa K. Los ácidos nucleicos han sido separa­ dos utilizando cloroformo: alcohol isoarru1ico y precipitados con etanol. El material obtenido ha sido mantenido en Robert's buffer y su concentración y calidad han sido verificadas en gel de agarosa al 1% (Figura 1). Figura l. Resultados de extracción de ADN El ADN extraído a partir de sangre periférica de primates del Nuevo Mundo es cuantificado y calificado en geles de agarosa al 1%. En la figura se ilustra los resultados obtenidos para dos individuos de Ateles fusciceps y dos individuos de Cebus albifrons. Amplificación mediante PCR Para la amplificación de los elementos Alu, se utilizó los oligonucle6tidos diseñados por Singer et al, 2003. La reacción de PCR se efectuó en 12,5ul de volumen final incluyendo 30 TJg de ADN, oligonucleótidos forward y re­ verse en concentraci6n 20 mM, buffer FailSafe premix IX (Epicentre, Madisson,USA)y 0,5U de Platinum Taq polimerasa (Invitrogen). Los programas de amplificación correspondieron a 2 minutos de pre-denaturación a 92° C y 30 ciclos de 45 segundos de denaturaci6n a 92° C, 60 se­ gundos de acoplamiento a la temperatura corres­ pondiente a cada par de oligonucleótidos y 60 segundos a 72° C para la extensión. Los produc­ tos de PCR fueron visualizados en geles de aga­ rosa junto a un marcador de peso molecular y registrados mediante fotografía (Figura 2). El análisis de las dimensiones de los amplificados Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vol. XXVII Números 1 y 2: 48-51, octubre del 2005 D Elementos Alu en el análisis filogenético molecular del infraorden Platyrrhini, Primates obtenidos fue hecho con el programa PhotoCapt MW V.10.01. Posterionnente la infonnación ob­ tenida fue tratada en términos de matriz y some­ tida al análisis con PAUP 4.0b.1O (12). 49 Con esta infonnación se procedió a reali­ zar el análisis filogenético mediante Neighbor­ joining. El árbol obtenido (Figura 3) permitió dilucidar relaciones de parentezco entre las es­ pecies analizadas. RESULTADOS DISCUSIÓN La amplificación mediante PCR de los elementos Alu: STK1, HBGF YVP permitió ob­ tener productos ort610gos para cada especie con variación de relativamente pocos pares de bases dentro de cada una (Figura 2 y Tabla 1). Figura 2. Amplificación del marcador Alu HBGF para Saimiri sciureus, Lagothrix lagotricha y Alouatta palliata Los productos de amplificación mediante PCR para el marcador HBGF oscillan entre 1073, 1080 Y 1093 pb pa­ ra Saimiri. En Lagothrix se amplificó un producto de 948 pb, mientras que para Alouatta se obtuvo un pro­ ducto de 968pb El hecho de haber amplificado median­ te PCR tres tipos de elementos Alu en el ge­ noma de las especies de platirrinos analizadas en este trabajo respalda fuertemente la mono­ filia propuesta para el infraorden ( 5, 6, 7, 8, 9). Por otra parte la presencia de productos de amplificación de diferentes dimensiones pa­ ra cada especie estudiada sugiere que las trans­ posiciones derivadas de este tipo de elementos Alu han tenido lugar durante la evolución de los monos del Nuevo Mundo. Las relaciones inferidas mediante el análisis de Neighbor-joining evidencian por u­ na parte, mayor cercanía entre Cebus y Saimi­ ri, sosteniendo el cIado Cebus - Saimiri pro­ puesto por evidencias citogenéticas (13) y mo­ leculares (6, 7, 8, 14) Y por otra entre Lagoth­ rix, Ateles y Alouatta respaldando el posicio­ namiento de estos tres géneros en la familia Atelidae. Considerando que los monos del Nuevo Mundo podrían con seguridad ser los protago­ nistas de un importante evento de radiación que o Tabla l. Productos de amplificación mediante PCR para los elementos Alu STK1, HBGF YVP en varias especies de platirrinos ecuatorianos Especie Elemento Alu STKl Elemento Alu HBGF Elemento Alu VP Cebus albifrons 1660 - 1686 pb 1040 - 1052 pb 870 - 880 pb Saimiri sciureus 1514 -1577- 1595 pb 1073 - 1080 - 1093 pb 818 - 858 - 880 pb Ateles belzebuth 1215 pb 877pb 890pb Ateles fusciceps 1200pb 866pb 890pb Lagotrhrix lagotricha 1159 - 1200 pb 948pb 800- 807 pb Alouatta palliata 1277 pb 968pb 882pb Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vol. XXVII Números 1 y 2: 49-51, octume del 2005 - Ludeña et al. 50 Figura 3. Arbol de Neigbor-Joining para los elementos Alu: HBGF, VP y STKl analizados en las especies: Cebus alb­ ifrons, Saimiri sciureus, Ateles belzebuth, Ateles fusciceps, Alouatta palliata y Lagothrix lagotricha. Saimiri .---- ........ .......... Sal sci23 • •• . . .. .. .. Sai scil9 •• • .. • \ •• Sal sci2I I I •• •• • \ •.. Sai scl12 .. . . .. . ........ Sai sci15 ~~---"¡i-..... -­ Cebus les pennitió ocupar nichos ecológicos muy va­ riados y divergir dramáticamente desde hace aproximadamente 40 millones de años, (11) es necesario abordar el análisis de su filogenia a la luz de varias fuentes de información genómica, así como de otros parámetros biológicos. La información obtenida en el presente trabajo ha sido sin duda de interés para verificar la presencia de una dinámica transposicional con valor filogenético en el genoma de prima­ tes y para entender mejor las relaciones entre especies. No obstante esta información será ampliada a un mayor número de especies y se­ rá integrada a aquella proveniente de otros mar­ cadores genómicos, que están siendo utilizados ya al momento, con el fin de aclarar la intrinca­ da filogenia de los platirrrinos. Agradecimientos Se agradece al Dr. Andrés Ortega de la Fundación Zoológica de Guay llabamba por su colaboración en la obtención del material bio­ lógico utilizado en esta investigación. Revista Ecuatoriana de Medicina y Ciencias Biológicas Vol. XXVII Números l y 2: 50-51, octubre del 2005 Elementos Alu en el análisis filogenético molecular del infraorden Platyrrhini, Primates REFERENCIAS L- TRTKOVA, K.; MAYER, W.; O'HUIGIN, C. & JAN K­ LEIN. Mhc - DRB genes and the origin of New World Monkeys. Molecular Phylogenetics and Evolution, 1995; 64: 408 -419. 2.- HOULE, A. The origin of Platyrrhines: an evaluation of the Antartic scenario and the floating island model. Ame­ rican Journal oC Physical Antropology, 1999; 109:541-559. 3.- SCHRAGO, C. & RUSSO, C. Timing the origin of New World Monkeys. Molecular Biology and Evolution, 2003; 20 (10):1620-1625. 4.- GOODMAN, M.; PORTER,C.; CZELUSNIAK, J.; PA­ GE, S.; SCHNEIDER, H.;SHOSHANI, J.; GUNNELL, G. & GROVES, C. Towards a phylogenetic classification of primates based on DNA evidence complemented by fossil evidence. Molecular Phylogenetics Evolution, 1998; 9.585-598. 5.- HOROVITZ, 1. & MEYER, A. Sistematics of New World Monkeys (Platyrrhini, Primates) based on 16S mitochon­ drial DNA sequences: a comparative anaIysis of different weighting methods in cladistic analysis. Molecular Phy­ logenetics Evolution, 1995; 4: 448-456. 6.- HOROVITZ, l.; ZARDOYA, R. & MEYER, A. Platyrr­ hine systematics: a simultaneous analysis of molecular and morphological data. American Joumal oC Physical Antropology, 1998; 106: 261-268. 7.- PORTER, C.; SAMPAIO, l.; SCHNEIDER, H.; SCH­ NEIDER, M.; CZELUSNIAK,J. & GOODMAN, M. Evidence on primate phylogeny from epsilon-globin ge­ ne sequences and flanking regions. Journal oC Molecu­ lar Evolution, 1995; 40:30-55. 51 8.- PORTER, C.; CZRLUSNIAK, J.; SCHNEIDER, H.; SCHNEIDER, M.; SAMPAIO, L. & GOODMAN, M. Se­ quences of epsilon globin gene: implication for systema­ tics of the marmosets and other New World primates.Ge­ ne, 1997; 205:59-71.. 9.- SHCHNEIDER, H.; SHCNEIDER, M. P. C.; SAMPAIO, l.; HARADA., M. L.; STANHOPE, M.; CZELUSNIAK, J. & GOODMAN, M. Molecular phylogeny of the New World Monkeys (Platyrrhini, Primates). Molecular Phy­ logenetics and Evolution, 1993; 2: 225. 10.- TIRIRA, D. (1999) Manúferos del Ecuador. Publicación especial N" 2. Museo de Zoología de la PontificiaUniver­ sidad Católica del Ecuador. Quito. pp. 67-72 11.- SINGER, S. S.; SCHMITZ, J.; SCHWIEGK, C. & ZISCHLER, H. Molecular cladistic markers in New World monkeys phylogeny (Platyrrhini, Primates). Molecular Phylogenetics and Evolution, 2002; 26: 490-501 12.- SWOFFORD D., PAUP 4.0.b.1O. Phylogenetic Analysis using Parsimony and other methods. Computer Program distributed by Sinauer Associates. Sunderland, Massachu­ setts. 200 l. 13.- CHIARELLI, A. B., The karyology of South American Primates and their relationship to African and Asian spe­ cies. In Ciochon, R.L., Chiarelli,A.B. (Eds.), Evolutionary Biology of the New World Monkeys and Continental Drift. Plenum Press. New York, 1980; pp 387-398. 14. HARADA. M.; SCHNEIDER. H.; SCHNEIDER, M.; SAMPAIO, l.; CZELUSNIAK & J.; GOODMAN, M. D­ NA evidence on the phylogenetic systematics of New World Monkeys :support for the sister-grouping of Cebus and Saimiri from two unlinked nuclear genes. Molecular Phylogenetics Evolution, 1995; 4: 331-349. D Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vol. XXVII Números l y 2: 49-51, octubre del 2005 D TRABAJOS DE REVISIÓN ----------------------------------------Jf---­ La originalidad del desarrollo de la anestesia en el Ecuador.- Siglo XX - 1960 - 2000 Oswaldo Morán Sección Ciencias Biológicas y Naturales Casa de la Cultura Ecuatoriana Recibido 16 marzo, 2005, aprobado 28 marzo 2005-06-28 INICIACIÓN DE LA ANESTESIA EN EL ECUADOR D Existen algunos temas universales que por interés de su contenido, han sido aprove­ chados como sentimiento de reacción de los hombres, esa es la Anestesia aquel arte de la medicina a la cual se consideraba como nueva y que nada había que de ella escribir, pero que en realidad tiene una dualidad, es vieja y joven. Tan vieja como la misma vida, si recordamos la primera anestesia que según el Génesis, "Adán fue sumido en un profundo sueño, para extraér­ sele una costilla y llenar de carne aquel vacío". Joven si se piensa que a mediados del siglo x­ IX Alfred Velpou gran cirujano francés decía "el suprimir el dolor en las intervenciones qui­ rúrgicas es una quimera..." en cirugía, bisturí y dolor son dos palabras indesligables la una de la otra y, es necesario admitir y aceptar esta rea­ lidad". Ya poco antes de nacer la vida del futu­ ro ciudadano comienza a estar influenciada por la Anestesia. Durante el trabajo de parto, casi todas las mujeres solicitan les sea admi­ nistrada alguna forma de anestesia y, unas bas­ tante más, otras bastante menos, toda aneste­ sia administrada a la madre tiene repercusión en el niño. En adelante, se supone que un ciudadano de vida normal será sometido a algún tipo de anestesia por lo menos tres veces en su vida pa­ ra fines quirúrgicos. La palabra anestesia deriva del griego ANAISTHESIA que significa abolición total o parcial de la sensibilidad del cuerpo o de una de sus regiones provocado por enfermedad o bien por anestésicos. El empleo de las propiedades anestésicas de ciertas substancias como el alco­ hol, las inhalaciones de mandrágora, etc., para reducir el dolor en las intervenciones quirúrgi­ cas, se conoce desde la antigüedad, pero solo a mediados del siglo XIX se iniciaron las técni­ cas científicas de la anestesia. El accidente anestésico es habitualmente llamativo y trasciende al público con facilidad debido a que por lo general, sucede como re­ lámpago en el cielo sereno. El accidente anestésico le ha dado una falsa jerarquía a la especialidad sencillamente, la gente le tiene miedo a la anestesia. La historia de la medicina nos relata que el dolor es tan antiguo como el hombre y muestra los esfuerzos que se han hecho para combatirlo. Existió un triángulo de actores humanos que tomaron parte en uno de los actos mas tras­ cendentales y valientes en la Historia de la Me­ dicina como fue la primera demostración públi- Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vol. XXVII Nl1meros l y 2: 53-56. octubre del 2005 o Morán 54 ca intencional y exitosa de la Anestesia para fi­ nes quirúrgicos en el Hostpital de Boston el 16 de octubre de 1846: Gilberth Abbot fue el pa­ ciente, Willians G. Morthon el anestesista y Jhon Collins el cirujano, fue algo que dio la vuelta al mundo y que a través de los años lle­ gó a nosotros. El objetivo fundamental de la anestesia como especialidad Médica es prevenir y aliviar el dolor, su primera responsabilidad es mante­ ner la vida del paciente y esto se lo consigue no con una simple técnica de anestesia, sino con el conocimiento y dominio farmacológico de los agentes anestésicos y analgésicos, sus variadas formas de asociación, uso y administración de dichos fármacos. En los tiempos de inicio de la Cirugía en Quito, la anestesia se nutrió con los conoci­ mientos del Dr. Alejandro Schibbye, de nacio­ nalidad sueca, quien en el año de 1870 introdu­ jo la anestesia en la Capital del Ecuador, acon­ sejado por Willis "Usó en las experiencias sin dolor el tan afamado anestésico protóxido de nitrógeno que se preparaba diariamente fresco y puro, tal como lo aconsejaba su descubridor", Joseph Priestley. En el año de 1874 Gayraut hace uso de la anestesia clorofórmica de aplicación con pañuelo o compresa en el Hospital San Juan de Dios, el, junto con el Doctor Domeck, fue­ ron los que introdujeron el cloroformo en el País. Dato curioso es aquel que nos da a cono­ cer que en el año de 1872, según los archivos del Hospital San Juan de Dios, esa botica esta­ ba bien provista. El Doctor Antonio Barahona fue su boticario, hace constar en el inventario de febrero de ese año, abundante lista de droga útiles y aparatos de anestesia. En 1889, se usa por primera vez la anes­ tesia local por cocaína, en cirugía dental por el guayaquileño Doctor Manuel G. Ramos. En 1894, a fines de este año, el profesor de la Facultad de Medicina de Guayaquil, Francisco Martínez Aguirre operó con aneste­ sia al éter, atribuyéndole el mérito de introdu­ cir este tipo de anestesia, y que hasta aquel en­ tonces, se 10 hacía con el cloroformo o la com­ presa. En 1901, la primera laparatomía con anestesia al eter, en el San Juan de Dios, lo hi­ zo el médico guayaquileño Dr. Julio Vásconez. En 1903, en Guayaquil el Doctor Miguel Achig aplica la primera raquianestesia. En 1906, el Doctor Ezequiel Cevallos Zambrano, Profesor de Fisiología de la Facul­ tad de Quito, practica la anestesia local en ope­ raciones de fimosis por primera vez en Quito con procaína. Posteriormente en la segunda década del novecientos en el único hospital existente en Quito, el Hospital San Juan de Dios, se admi­ nistraba el cloroformo con el aparato de Ri­ card y el éter se suministraba con el aparato de Ombredanne. La Medicina y Cirugía nacional marcha­ ron paralelas con la Escuela Francesa. Los sis­ temas de enseñanza, textos, plan organizativo de sus hospitales dependían en gran parte de la influencia de ese País. La concepción de las sa­ las de cirugía, los equipos quirúrgicos, los apa­ ratos y las técnicas anestésicas eran inspirados en la Escuela Gala. Entre nosotros se sistematizó el que la anestesia era parte del conocimiento del ciruja­ no, y no constituía aún especialidad, es mas es­ taba a cargo de enfermeros o hermanas de la ca­ ridad, hasta que en 1924 el Dr. Isridro Ayora, Director de la Maternidad, consideró que la Anestesia debe ser administrada por estudian­ tes de sétimo año de medicina o por residentes. Para la década de los treinta, existían otros 2 hospitales en Quito, el Eugenio Espejo y el Militar, la anestesia general era administra­ da bajo cloroformo o éter. En 1938, se realizó la primera anestesia peridural en la práctica anestésica de Quito. por el Doctor Elías Gallegos Anda, así como la Avertina y el Evipán Sódico en Riobamba son usados por primera vez. En 1950, el Doctor Rafael Compte en Guayaquil, realiza la primera intubación naso­ traqueal y orotraqueal, así como introduce el u­ so de los relajantes musculares como coadyu­ vantes anestésicos. Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vol. XXVII Números 1 y 2: 54-56, octubre del 2005 D La originalidad del desarrollo de la anestesia en el Ecuador.- Siglo XX - 1960 - 2000 Para los años cuarenta y siete ya existía la Clínica del Seguro Social en Quito, para obre­ ros y empleados, la mejor equipada para aquel entonces, con capacidad para 85 enfermos, ser­ vicios modernos tales como: Radioterapia y Anestesia de Gases. Esta se administraba con un aparato MC. Kensson, con ciclopropano óxido nitroso y etileno. 1960-2000 Es a partir de la década de 1960, en que la anestesia toma verdadero cuerpo como espe­ cialidad autónoma en el País. Llegamos los primeros graduados en las Universidades de Iso Estados Unidos y se instalan los primeros ser­ vicios anestésicos de esa escuela, con su des­ pliegue de aparatos de administración múltiple de gases, Resucitadores, Monitores, etc, predo­ minó en el Quirófano la sistematización ameri­ cana. Se desarrolla una acción que bien pudo llamarse renovadora. En 1962 se funda la Sociedad de Aneste­ siología de Pichincha y dos años después reco­ nocida oficialmente adquiere personería jurídi­ ca con sede en Quito y se afilia a la Confedera­ ción Latinoamericana de Sociedades de Anes­ tesia (CLASA) y luego a la Federación Mun­ dial de Sociedades de Anestesia (WPSA). En el mismo año participamos en el cuarto Congreso Mundial de Anestesia, en Sao-Paulo-Brasil. Luego de un cursillo de preparación, trajimos el aparato de TAKA-OKA, nombre de su inven­ tor, conocido como de bolsillo y aplicable para cirugía de corazón abierto, tiene un peso de 2.5 kilogramos y un sistema abierto con respirador automático. En 1966 en la Clínica del Seguro Social de Quito, iniciamos con todo éxito por primera vez en el País, luego de estudios en el Catholic Hospital de Bruselas - Bélgica; Catholieke Uni­ versiti, Nimegen - Holanta; Stadtesche-HR­ NEN-Nanstalten; Bremen - Alemania, la Neu­ rolepto-Analesia (N.L.A.), que produce una analgesia absoluta mediante el Thalamonal con sus dos componentes: Fentanyl y el Dihidro 55 Benzoperidol nueva droga en investigación que actúa selectivamente sobre el Tálamo, Hipotála­ mo, Sistema Reticular y Gama Neuronas sin provocar una depresión general del organismo. Es la llamada "Anestesia General sin Gases". En el mismo año, los primeros trabajos de la especialidad son presentados en el VIII Con­ greso Latinoamericano, en Caracas. Estos tra­ bajos sirvieron para el establecimiento de nor­ mas precisas de la presentación de experiencias científicas de la de especialidad en el País, para su época. En febrero de 1966 en Quito se inicia en la Clínica del Seguro Social el Servicio de Recuperación Post-anestesia. Igual se organiza la cátedra de Anestesiología, primera en el País. Iniciamos con 32 estudiantes que posteriormen­ te un buen número de ellos se titularon como anestesiólogos luego de obtener su Diploma en el primer curso de Post-grado. Año 1967, en la Clínica del Seguro So­ cial de Quito, primera operación de corazón abierto. La Facultad de Ciencias Médicas, de acuerdo con la Sociedad Ecuatoriana de Anes­ tesia, Núcleo de Pichincha, otorga los primeros certificados de Anestesistas a quienes hayan ejercido la práctica por cinco años en hospitales docentes. La especialidad se nutrió con la orga­ nización de cursos y cursillos didácticos para médicos y estudiantes internos, el shock es es­ tudiado con dominio casi total en el campo de la Anestesiología; se planifican servicios de re­ cuperación mejor dotados y también Cuidados Intensivos. Se elaboran normas para comportamien­ to en áreas quirúrgicas, como aquellas que pa­ ra entrar en el Quirófano, debían hacerlo con un mandil, cubierta la cabeza con gorra, usar mas­ carillas, además cubrir los zapatos con botas de tela que posteriormente se las confeccionó para el grupo operatorio con plantilla anti-estática. Se organizan jornadas médicas y luego el 1ero Congreso Nacional de Anestesia. En Quito, la Facultad de Ciencias Médi­ cas inicia el primer curso de Post-grado de Anestesia en el País; me acompañaron un equi­ po de prestigiosos profesionales como docen­ tes. 1968, por disposición Ministerial, se prohi- o Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vol. XXVII Números 1 y 2: 55·56, ocmbre del 2005 _ .. __ ... _._­ - Morán 56 be ejercer la especialidad en las Instituciones Estatales y privadas a quienes no tengan el títu­ lo de Anestesiólogo; además se oficializa a tra­ vés del Ministerio de Salud el uso del registro o protocolo único en el curso de la anestesia, de­ sechando de esta manera las hojitas o recetarios donde las graficaban. Nuevas técnicas y fármacos utilizamos: Inhalatorios; Fluhotano y Pentrane, locales, la Xilocaina, el La c-43, etc., y publicadas sus ex­ periencias en revistas nacionales y extranjeras. El bloqueo del Plexo-Braquial por vía axilar o endovenosa con torniquete, reempla­ zan al riesgoso supraclavicular que entrañaba el peligro de herir la pleura; se reinicia la técnica conductiva Epidural que por su eficacia y dura­ bilidad supera a la raquídea. En el tratamiento del shock se usa un nuevo sustituto del plasma, Haemaccel y la nueva droga anti-shock, Beta Adrenérgica Me­ taproterenol. OTRAS ALTERNATIVAS PARA LA SALUD.­ ACUPUNTURA, con estudios en Cen­ tro Acupunturistas Akirainamoto de Kyoto ­ Japón, S.T.LEE, Seúl- Korea del Sur en 1972; TERAPIA NEURAL o ACUPUNTU­ RAALEMANA, estudios en la Universidad de Gutemburg, Alemania Occidental, 1976; CLÍNICA DEL DOLOR, término usa­ do para estas nuevas terapias; se nos prohibió usar en el Hospital, ahí valía la primacía del concepto que el dolor debía ser tratado de la séptima vértebra cervical hacia arriba por el neurólogo y de la séptima cervical para abajo, por el traumatólogo; nosotros usábamos en la práctica privada, nuestras técnicas aprendi­ das. El Hospital Andrade Marín a la fecha tie­ ne un moderno servicio de la "Clínica del Do­ lor" con especialistas. La excelencia en el pro­ greso de la anestesia en el País, permitió al ci­ rujano afrontar con éxito los riesgos ce las ope­ raciones en el paciente especialmente los del corazón y el cerebro. Desde el Quirófano y la Docencia Uni­ versitaria se formó un nuevo tipo de médico. Mi respeto y admiración a lo logrado por estos profesionales que me acompañaron y per­ mitieron hacer histórica la evolución de esta ra­ ma médica que hoy la consideran de alta espe­ cialidad. BffiLlOGRAFÍA: ARCOS GUALBERTO.- Evolución de la Medicina en el Ecuador, 3ra. Edición. Casa de la Cultura Ecuato­ riana 1979. ASTUDILLO CELIN.- Profesor Dr. Isidro Ayora, Médi­ co Innovador y Presidente de la República. Quito 1983. FALCONÍ VILLAGÓMEZ J.A ..- Asclepio y Cromos, 1961. GARCÉS CABRERA ENRIQUE.- Perfil del doctor Eu­ genio Espejo, 1974. HOWARD RILEY RAPER.- Man Against Pain, New Y­ orle, Prentice-Hall Inc, 1945. HERMIDA CESAR PlliDRA.- Resumen de la Historia de la Medicina Ecuatoriana, Segunda Edición. Cuenca 1979. LARCO NICOLÁS.- Historia de la Medicina Ecuatoria­ na, ler. Tomo 1990. REVISTA "TERHAPIA".- Life. 1974 MADERO MAURO.- Historia de la Medicina en la Pro­ vincia del Guayas. 1955. MORAN PINTO OSWALDO.- Síntesis Cronológica de la Anestesia en el Ecuador. Editorial Universita­ ria.l%9. MORAN PINTO OSWALOO.- Bibliografía, Revista Ecuatoriana de Medicina y Ciencias Biológicas. Vol. XVII. No. 2 Julio Diciembre 1981. MANAKA YOSHIO M.O. and lANA URQUHART. PH.D.- The Layman's Guide To Acupunture. Tokyo. 1972. M.MIYAZAKI - K.IWTSUKL M. FUJITA.- Excerta Medica. Fifth World Congress Of Anaesthesiolo­ gits. 1972. MONTERO JOSE A..- Maestros de ayer y de hoy. 1957 NARANJO PLUTARCO VARGAS.- Contribuciones Aborígenes a la Medicina Ecuatoriana. Revista, Medicina y Ciencias Biológicas Nos. 1 y 2 a Ju­ nio 1970. PAREDES BORJA VIRGILIO.- Historia de la Medicina Ecuatoriana. 1%9. SAMANlliGO JUAN JOSÉ.- Cronología Médica Ecua­ toriana. 1957. Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vol. XXVII Números 1 y 2: 56-56, octubre de12005 o Bioética y práctica médica Oswaldo Cbaves Sociedad Ecuatoriana de Bioética Presentado 5 abril 2005, aprobado 19 abril 2005 o El ejercicio profesional de la medicina demanda, de manera muy especial, profunda responsabilidad en cuanto a su competencia científica y, a la vez, la más elevada sensibili­ dad humana y compromiso ético. No es suficiente poseer los conocimien­ tos y el arte necesarios para saber diagnosticar y tratar enfermedades, por sobre toda conside­ ración de experticia se requiere de la calidad moral para entender al ser humano a quien atiende, respetar su dignidad, sensibilidad, do­ lor, minusvalía, necesidades y sentimientos. Junto a él, considerar a su familia y, en todo momento, apreciar que el estado de salud no es la simple ausencia de enfermedad, que el pro­ ceso salud-enfermedad es multicausal, con fac­ tores que no solamente son de carácter biológi­ co, físico o químico, que también son ambien­ tales y sociales, económicos y emocionales. Aceptar que la miseria, el estrés, la exclu­ sión, las guerras y la humillación son motivos masivos de ausencia de salud, al privar a hom­ bres y mujeres de todo bienestar y de posibili­ dades de trabajo, a los niños de educación y alegría, a los adultos mayores, de paz, amor y comprensión y, a todos, de su desarrollo inte­ gral y de un ambiente saludable. Esta consideración, impone la necesidad de entender que la definición de salud que la OMS adoptara en 1948, ya no es suficiente, que la inhumana globalización, la deuda externa, la pobreza extrema, la ciencia sin conciencia, la depredación de la naturaleza y las imposiciones imperiales del poderoso, reclaman volver a la poética y geocéntrica definición quechua de sa­ lud, que al traducirla al español dice: "Salud es la relación armónica con uno mismo, con los otros seres humanos, con la Pachamama en la búsqueda de una tranqui­ lidad espiritual, individual y social". Con esta interpretación de salud que im­ plica la armonía del ser humano con su hábitat, la sociedad, su familia, su bienestar integral, su conciencia y sus valores morales, es posible abordar el significado, características y com­ promiso de la práctica médica, independiente­ mente de si es pública o privada, de su especia­ lidad, del nivel de atención a que corresponda o si se trate de la promoción o prevención en sa­ lud, porque en todo caso estará subordinada a los principios fundamentales de la Bioética. Igual si, en el ejercicio profesional, realiza in­ vestigación clínica, ejerce la docencia o cumple funciones relacionadas con la gestión en salud, invariablemente, sus valores éticos y morales deberán guiar su desempeño en todos los roles que le corresponda asumir. Es oportuno, por tanto, referirnos esque­ máticamente a esos diferentes roles de la prác­ tica médica, para analizar la vigencia de la Bioética en los mismos. Así, en la función de servicio, tanto a nivel asistencial como de pro­ moción y prevención en salud, su relación fun­ damentalmente será con el paciente, su familia y la comunidad, con sus colegas y demás miembros del equipo de salud, en escenarios donde tendrá particular importancia: • La competencia profesional y su honesti­ dad para referir al paciente a quien pueda atenderle con mayor idoneidad. Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vol. :xxvn Números 1 y 2: 57-59. octubre del 2005 o 58 Chaves El respeto permanente a: • • • • • • La vida La dignidad del ser humano La autonomía del paciente La confidencialidad La veracidad Las creencias convicciones y cultu­ ra de los pacientes, familiares y co­ munidad • El consentimiento informado • La relación médico/ paciente/ familia! comunidad, que deberá ser: humana, cá­ lida, ética y sin distingo de naturaleza al­ guna, social, económica, política, religio­ sa, étnica o cultural. • El cumplimiento permanente e irrenun­ ciable de los principios de beneficencia y no maleficencia • Una relación consecuente y solidaria en­ tre colegas • Discreción, prudencia, mesura y ética en la publicidad y honorarios profesionales • El sentido de justicia, equidad y solidari­ dad en todos los servicios de salud, sin exclusión alguna, particularmente en los que se preste atención integral en salud. Orientada la atención integral en salud fundamentalmente hacia la promoción y pre­ vención en salud, a los espacios y maternidad saludables, a la nutrición y seguridad alimenta­ ria, al tratamiento de enfermedades infecciosas de la infancia, emergentes y re-emergentes, a la atención al adulto mayor y a los minusválidos. Esta obligación de la atención en salud debe ca­ racterizarse por tener cobertura universal y por ser responsabilidad indelegable del Estado, an­ te la cual al médico le corresponde exigir y con­ tribuir a garantizar la eficiencia y eficacia de los servicios de salud, para así cumplir los prin­ cipios bioéticos de justicia, equidad y solida­ ridad, con calidad y atención prioritaria a los más desprotegidos. ¡ En el desempeño de la función de inves­ tigación, epidemiológica o clínica, que puede corresponderle al médico, cuando cumple su práctica profesional, deberá respetar, además, el consentimiento informado y las normas y convenios internacionales en cuanto a la inves­ tigación con seres humanos, manteniendo siempre una concepción axiológica y teleológi­ ca de una investigación que priorice la búsque­ da y aplicación de soluciones a las necesidades primarias de salud, a la vez que no subordine la experimentación a la trascendencia del éxito académico, publicitario o económico y que mantenga, por sobre todo interés, la vigencia del legado de Potter cuando al crear la Bioéti­ ca nos dijo: "La bioética une las ciencias biológi­ cas y la filosoña para tender un puente ha­ cia el futuro y promover la supervivencia humana".l La función de educación que, en los dife­ rentes escenarios y oportunidades de servicio, debe cumplir el médico, le impone la misión de modelaje frente a los estudiantes a quienes debe servir de ejemplo y la de ser un símbolo de credibilidad y confianza para la población, en general. La otra función que frecuentemente le corresponde al médico, en su desempeño públi­ co o privado, es la de gestión, en la cual adicio­ nalmente a lo expuesto deberá mantener, con celo, entre otros valores los de: Honestidad, disciplina, tolerancia y efi­ ciencia Protección a sus colegas en los casos in­ justos de acusación por mala praxis médica y sanción cuando se justifique En adición a todo lo dicho, es necesario considerar que en la práctica profesional exis­ ten aspectos particulares de especial trascen­ dencia y que pueden significar motivo de con­ flicto, por lo cual amerita una referencia parti­ cular a ellos, especialmente porque no han sido Potter, Van Rensselaer. Bioethics, Bridge to the future. Englewood Cliffs, N.J.:Prentiee - Hall, Ine. (1971 J. Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vol. xxvn Números 1 y 2: 58-59, octubre del 2005 D ------~ ..- _ . __.. .. _--~.-_._--_ _---~.-------_ Bioética y práctica médica ..- _ . __..------.- -_.~_._~ .. ­ 59 motivo de una seria preocupación en el proceso de formación del médico. Tal el caso de la con­ ducta a seguir frente a la muerte, al paciente terminal, al minusválido el enajenado mental o la terapia del dolor. Igualmente, tampoco se analiza la actitud del médico ante la prescrip­ ción de medicamentos no justificados, el tras­ plante de órganos, la reproducción asistida, el aborto, la eutanasia o la eugenesia. Ante la muerte o un paciente terminal, el moribundo ni sus familiares esperan nuevas prescripciones farmacológicas o sofisticados y costosos auxilios tecnológicos, que prolonguen penosamente su existencia, anhelan un acom­ pañamiento cálido, compresivo, veraz y respe­ tuoso de su dignidad humana. El minusválido y el enajenado mental, igualmente, merecen la mayor consideración y apoyo, de la misma manera que quien espera ayuda para mitigar su dolor. Prescribir medicamentos sin la seguridad de su necesidad o de su efecto, es atentar con­ tra los principios de beneficencia y no malefi­ cencia' en tanto que silenciar o propiciar la compra o venta de órganos implica mercantili­ zar un hecho tan noble y generoso como su do­ nación. No existe justificación alguna para que un médico sugiera o asuma complicidad en una decisión que signifique atentar contra la inte­ gridad de la vida de un ser humano; y, por otra parte, concederse la potestad de adoptar deci­ siones o aconsejar paternal o ligeramente res­ pecto a la reproducción asistida, el aborto, la eutanasia o la eugenesia, implica irrespetar el principio bioético de autonomía, oponerse a su voluntad e interferir en su capacidad de razona­ miento y reflexión. Alternativas muy difíciles que, en ningún caso, deberían aceptarse como responsabilidad de un solo médico, deberían compartirse con otros colegas y, de preferencia, cuando sea posible, consultarse al Comité Hos­ pitalario de Bioética o su equivalente. Es por todo lo expuesto, que valga la oportunidad para insistir en la necesidad de que las Sociedades Nacionales de Bioética estimu­ len y ayuden a la constitución de los Comités Institucionales de Bioética, en hospitales y cen­ tros de investigación, en sociedades científicas y gremiales, en universidades y organizaciones locales y nacionales de bienestar social. Su creación y funcionamiento continuo y apropiado, constituirán garantía para el de­ sarrollo de una investigación al servicio de la humanidad y la naturaleza, para que disminu­ ya la mala praxis médica, se combata el mer­ cantilismo en el ejercicio profesional y el de­ recho universal a la atención en salud no sea letra muerta de las constituciones o recurso demagógico de los políticos en las campañas electorales y constituya un beneficio bioético auténtico para todos los seres humanos, sin distingo de naturaleza alguna, en todos los pueblos de la tierra. D Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vol. XXVII Números 1 y 2: 59-59, octubre del 2005 D D EI denominador "nacidos vivos" en investigación de salud oública Estuardo Albán, César Hermida, Jorge Hermida, juan Vásconez, Mario Vergara, Miguel IDnojasa, Miguel Machuca, Nelson Oviedo REDSISMIL', Ecuador 'Réét)'il" ~ ffijri:o 1005, AM"l,).Iii" 1¡ ÜW'11005 RESUMEN: Con el objetivo de ajustar los datos nacionales y mostrar la metodología para ajustes provinciales y locales, se cal­ culan las inscripciones tardías de nacidos vivos a partir de la información del Instituto de Estadísticas y Censos. Se obtiene un de­ nominador más apropiado para los cálculos de mortalidad infantil y materna, y las estimaciones de coberturas del programa de va­ cunación y otros. Se compara este dato con el número total de inscripciones en el año, para destacar la inconveniencia de su uso, así como la inconveniencia de optar por las proyecciones del número de menores de un año como alternativa para la estimación de nacidos vivos. PALABRAS CLAVES: Nacidos vivos, mortalidad infantil, mortalidad materna, coberturas de vacunación. INTRODUCCIÓN: En el año 2000 los 189 Estados Miembros de las Naciones Unidas se reunieron en la Cum­ bre del Milenio y aprobaron ocho objetivos y 18 metas para combatir la pobreza, el hambre, las enfermedades. la discriminación contra la mu­ jer, la degradación de las tierras y el analfabetis­ mo. El Objetivo 4 propone "reducir la mortali­ dad en la infancia", y la meta 5 "reducir en dos terceras partes, entre 1990 y 2015, la tasa de mortalidad de los niños menores de cinco años". Dentro de esta última se encuentra el indicador de Mortalidad Infantil medido por la tasa de muertes de niños menores de un año por cada mil nacidos vivos, en el año. El Objetivo 5 pro­ pone "mejorar la salud materna", y la meta 6 "reducir en tres cuartas partes, entre 1990 y 2015, la tasa de mortalidad derivada de la ma­ ternidad". Esta tasa o razón se refiere a las muertes debidas a causas del embarazo, parto o puerperio por cada mil nacidos vivos en el año. El denominador de nacidos vivos tiene, enton­ ces, un carácter crucial, porque además permite calcular las coberturas de vacunación y otras ta­ sas de morbi mortalidad. Si bien las Encuestas han contribuido de manera notable al conocimiento de los indicado­ res (muchos de los cuales no se los puede cono­ cer de otra manera), y deben ser mantenidas en el tiempo, se propone fortalecer las Estadísticas Vitales del Registro Civil, todas procesadas por el INEC, en el entendido de que es indispensable disponer de las estadísticas oficiales públicas y de los resultados de las encuestas2• 1.- REDSISMIL, Red del Sistema de Indicadores de Salud para los Objetivos del Milenio, Ecuador. Estuardo Albán [email protected].¡:ov.ec César Hermida [email protected].¡: Jorge Hermida [email protected] Juan Vásconez [email protected]¡: Mario Vergara mario.ve([email protected] Miguel Hinojosa [email protected] Miguel Machuca [email protected].¡: Nelson Oviedo [email protected] 2.- Desde hace dos décadas, en el marco internacional de la "disminución del tamaño del Estado" se ha dado, en el caso del INEC y el MSP. una notable reducción de personal (desde las parroquias para el Registro Civil y las Estadísticas Vitales y del personal para procesamiento en cantones, provincias y nivel central), aparte de una disminución progresiva del valor real de los salarios. El propio Censo de 2001 tuvo dificultades de financiamiento y retraso. Esta tendencia implica el riesgo de provocar inefi­ ciencia en las responsabilidades del Estado, deterioro de sus productos, y por lo tanto una baja calidad de la información ofi­ cial. Al mismo tiempo se ha dado una corriente que busca obtener todos los indicadores mediante encuestas apoyadas con fon­ dos de cooperación internacional. Se trata de defender y robustecer tanto las funciones públicas como aquellas privadas, su­ perando la tendencia a descuidar las primeras en beneficio de las segundas. Un fortalecimiento de 10 público y lo privado y u­ na fuerte interrelación de los dos parece lo más razonable. Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vol. XXVII Números 1 y 2: 61-65, octubre del 2005 D A l1lin et al. 62 Método: Se analiza la consistencia de los datos del denominador utilizando las estadísticas pú­ blicas del INEC para los nacidos vivos, así co­ rno la información del último Censo Nacional (2001). En el primer caso se suman los regis­ tros tardíos de cada año para calcular el total re­ al, y luego se compara el número, tanto con el total de inscritos en el año, corno con los datos de menores de un año del censo y sus proyec­ CIOnes. Corno el 59.63 % de promedio de inscri­ tos en el mismo año del período, es aproximado al 57.60% del promedio de las inscripciones del mismo año sobre el total de inscripciones an­ uales, Cuadro 3, se podría pensar que el dato de inscripciones totales puede suplir, a nivel na­ cional, al dato de nacidos vivos en el año (yen provincias y cantones "sólo si sucediera lo mis­ mo"), sin embargo, es menester recordar que se trata de un promedio, pues cuando se compara los números de cada año, que son los que se uti­ lizarían, las diferencias resultan notables, Cua­ dr04. Resultados: Discusión: Los nacidos vivos inscritos a partir de El denominador de "nacidos vivos" es 1995 constan en el Cuadro 1, con aquellos re­ crucial. Lamentablemente, ante la falta de ajus­ gistrados en el mismo año y en los años subsi­ guientes. El Cuadro 2 muestra los porcentajes. te y uso de las estadísticas locales, en el país se ha utilizado, a partir del Censo de 1990, un gro­ Se observa que alrededor de un 60% dna­ sero estimado nacional para las necesidades lo­ cimientos se inscriben en el mismo año, mien­ tras un 28% lo hace en el año siguiente, y un cales, basado en el porcentaje de menores de un año de la población total. Así, la población na­ 12% lo hace en años ulteriores (5.5% en el se­ gundo y el resto a partir del tercer año). Hay, cional se redondeó a 10 millones y los menores de un año a 300 mil, dando un porcentaje de 3 por lo tanto, alrededor de un 40% de inscrip­ % para el estimado del denominador de nacidos ción tardía en el nivel nacional. Cuadro l. Nacidos vivos inscritos en el mismo año y en años subsiguientes Ecuador, 1995 a 2003 Nad­ i I Inscri­ tosen dos en: 1995 1996 1997 1998 1999 2000 2001 1995 181268 90072 13004 10798 8351 5605 3023 182242 88336 26142 12561 7265 4746 2773 169869 101889 20883 9765 5474 4506 3408 199079 76876 13740 6583 4467 4775 218108 87179 14317 7780 7774 202257 93892 16865 12426 192786 85384 1996 1997 1998 1999 2000 2001 ! I 183792 2002 2003 Fuente: lNEC, Estadísticas Vitales (Cuadro 1: Nacidos vivos inscritos por años). Elaboración: UNFPA Ecuador, 2004. xxvn Números 1 y 2: 62-65, octubre del 2005 I D 23243 91508 178549 2003 Revista Ecuatoriana de Medicina y Ciencias Biológicas· Vol. 2002 63 El denominador "nacidos vivos" en investigación de salud pública Cuadro 2. Inscripción de nacidos vivos en el mismo año y en años subsiguientes Porcentajes. Ecuador, 1995 a 2000 Ins.5 años + tarde Ins.6 años + tarde Total nacidos en el año 2.67% 1.79% 0.96% 312 121 3.87% 2.24% 1.46% 0.85% 324065 6.62% 3.09% 1.73% 1.42% 0.91% 315 284 24.94% 4.45% 2.13% 1.44% 1.50% 0.91% 308 206 64.18% 25.65% 4.21% 2.28% 1.46% 1.34% 0.84% 339 786 60.85% 28.25% 5.07% 2.16% 1.41% 1.38% 0.85% 332 359 Media 59.63% 27.87% 5.42% 2.83% 1.82% 1.48% 0.88% 323287 Ins.2 años + tarde Ins.3 años + tarde Año: Inseri­ tos, mismo año Inseri­ tos, año siguiente 1995 58.07% 28.85% 4.16% 3.45% • 1996 56.23% 27.25% 8.06% 1997 53.87% 32.31% 1998 64.59% 1999 ·2000 Ins.4 años + tarde ! Fuente: lNEC, Estadísticas Vitales (Cuadro 1: Nacidos vivos inscritos por años). Elaboración: UNFPA Ecuador, 2004. NOTA: Estimados para 6 años del 98,5 y 6 del 99 y 4,5 Y 6 del 2000, calculando el promedio de los últimos ños. Se puede concluir que un 60% se inscribe el mismo año, 28% al año siguiente y 12 en los años ulteriores vivos. Dicha estimación se ha utilizado en los servicios locales del Ministerio de Salud por más de una década. Estimar el denominador con el número de menores de un año tiene la ventaja de obtener fácilmente el dato a partir de las proyecciones de la población, pero tiene dos graves inconvenientes: primero que el estima­ do no corresponde al registro de nacimientos, y segundo que las proyecciones sólo se ajustan cada diez años con los censos. Por ejemplo a partir de 1990 la proyección anual de menores de un año mostraba un paulatino crecimiento, pero como se hallaba sobre estimado, al llegar a 294 645 como dato censal en el 2001, se ini­ cia, en la proyección, su declinación. Para di­ cho año, teniendo en cuenta la población censal total de 12479 924, el porcentaje de menores de un año corresponde al 2.36 %, mientras en el 2004 (con 289 789 menores de un año y una población de 13 026 891, ambos datos proyec­ tados) el porcentaje es de 2.22 %. El Cuadro 5 muestra que la población re­ gistrada en el Censo del 2001 como menor de un año constituye sólo el 73.37 % de aquella calculada con los registros tardíos para el mis­ mo año, y es el 80.50 % de aquella estimada con las proyecciones censales. La proyección censal es incluso sólo el 91.13 % de aquella calculada con los registros tardíos, pero este dato resulta clave, pues con el 8.87 % de dife­ rencia se puede estimar el número de nacidos vivos. Efectivamente, el Cuadro 6 muestra la proyección de la población de menores de un año, y la estimación con el cálculo de registros tardíos, señalando para el año 2004, por ejem­ plo, que el número de nacidos vivos es 315 493 (es decir sumando 25704 más nacidos, 8.87 % de incremento, al total de 289 789 de la pro­ yección). Conclusiones: Se debe calcular el registro tardío tanto para el nivel nacional como para el provincial, cantonal y parroquial, con estos ajustes es posi­ ble estimar el número de nacidos vivos para los años subsiguientes, a partir de las proyecciones censales. Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vol. XXVII Números 1 y 2: 63-65, octubre del 2005 A lLlin et al. 64 Es recomendable efectuar un ejercicio si­ milar al propuesto para los niveles provinciales y cantonales, a fin de obtener indicadores más consistentes. En el nivel local, es decir en las ci6n del registro civil, para utilizarla, ajustarla y mejorarla, a fin de que la consistencia se for­ talezca desde la base. Cuadro 3. Total de inscripciones en el año y número de nacidos en el mismos año. Porcentajes. Ecuador, 1996 a 2003 Años Inscripciones totales Porcentaje Inscripciones en el mismo año 1996 302 217 182 246 60.30 % 1997 288 803 169 869 58.81 % 1998 364684 199 079 54.58 % 1999 368 659 218 108 59.16 % 2000 345 715 202 257 58.50 % 2001 332 776 192 786 57.93 % 2002 314 577 183 792 58.42 % 2003 332870 178549 53.63% Total 2650301 1526686 57.60% Media Fuente: INEC, Estadísticas Vitales (Cuadro 1: Nacidos vivos inscritos por años). Elaboración: UNFPA Ecuador, 2004. parroquias, es conveniente conocer la informaCuadro 4. Número total de nacidos, con inscripción tardía, e inscripciones totales en el año. Ecuador, 1996 a 2003 i I I i i I i i Año: Total nacidos en el año· Inscripciones totales 1995 312 121 1996 324065 302217 1997 315284 288803 1998 308206 CJ 364684 ! 1999 339786 2000 332359 345715 2001 323303 332776 2002 257308** 314577 2003 249968** 332870 * Con inscripción tardía ** Estimado agregado el 40% a los nacidos e inscritos en el mismo año. Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vol. XXVII Números 1 y 2: 64-65, octubre del 2005 368659 j 65 El denominador "nacidos vivos" en investigación de salud pública Cuadro 5. Nacidos vivos totales comparados con los menores de un año del Censo del 2001 y de la Proyección Censal Ecuador, 2001. I I Total de Nacidos Vivos, estimados con el registro tardlo, 2001 323303 100% Estimación de la población de menores de un año para 2001, en base y a partir del Censo del 2001 294645 91.13 % deltotal de nacidos vivos (incluyendo registro tardío). VI Censo de Población, 2001. Población de menores de un año, Total 237209 80.50 % de la estimación de la población a partir del censo. 73.37 % del total de nacidos vivos (incluyendo registro tardío). Fuente: INEC, Estadísticas Vitales (Cuadro 1: Nacidos vivos inscritos por años). INEC, Estimaciones y Proyecciones de Población 1950 2025, Serie 01 No. 205, Octubre 2003. lNEC, VI Censo de Población 2001. Población de Menores de laño. Elaboración: UNFPA Ecuador, 2004. Cuadro 6 Proyección de población menor de un año y estimación de nacidos vivos. Ecuador 2004 a 2010 Año Proyección Censal (1) Estimación (2) 2004 289 789 315 493 2005 288 485 314 073 2006 287 679 313 196 2007 287 082 312 546 2008 286 616 312 038 2009 286 204 311 590 2010 285 786 311 135 CJ i J I Fuentes: (1) INEC Estimaciones y proyecciones de Población, 2003. (20 Estimaciones con 8.87 % de incremento en base al cálculo de registros tardíos, UNFPA, 2004. No es recomendable tomar el número de menores de un año y su proyección, peor el porcentaje de éstos de la población total, para reemplazarlo por el dato o estimación de naci- dos vivos. Tampoco es recomendable utilizar el número total de inscritos en el año como al­ ternativa del número de nacidos vivos en el año. Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vol. XXVII Números l y 2: 65-65, octubre del 2005 ------------------ ~ ........ CJ L-..... "Deshumanización de la medicina y bioética" Guillermo Álvares Domínguez Instituto de Radiología S.e. Av. Eloy Alfaro N29-214 y Alemania Recibido 31 marzo 2005, aprobado 21 abril 2005-06-28 En los ensalmos primitivos de la mágica medicina shamánica y en los procedimientos de la medicina hipocrática, se destacaban en for­ ma magistral el buen uso del lenguaje y la ade­ cuada y serena comunicación entre el enfermo y quien pretende aliviarlo, sanarlo o curarlo. Para comunicarnos - escribe un académico de la Lengua - "debemos descubrir que dice la melodía ajena, que contenido estricto oculta en la expresión fluída o entrecortada por la emo­ ción, o en la desarticulada por falta de destreza o por un pobre respaldo sociocultural. Sin ese puente obligado no hay comunicación , ni me­ dicina efectiva ni interlocutor posible". El examen clínico siempre fue un acto sa­ grado y solemne, en el que se comunican el co­ nocimiento, la pericia y la pulcritud del exami­ nador con la confianza y la buena fe del exami­ nado. Los progresos tecnológicos de la medici­ na moderna y la aparición de los profesionales especialistas, parece ser que modificaron los esquemas de diagnóstico y tratamiento alter­ nando además la relación personal médico-pa­ ciente. Si bien es verdad que la bondad diag­ nóstica de la tecnología electrónica despertó u­ na inusitada confianza en los actores del exa­ men clínico, no es menos cierto que ellos se ro­ botizaron, el primero accionando botones y lu­ ces en su pantalla, y el segundo cambiando su nombre por un número. Sin embargo la medicina próxima deberá ser cada vez más tecnológica y cada vez más clínica. Habida cuenta que la herramienta elec­ trónica aun no puede reemplazar al cerebro hu­ mano, se adoptarán criterios y protocolos que sepan discernir lo significativo de lo no signifi­ cativo, a fin de no derivar de inmediato hacia el especialista a personas que pueden ser tratadas por un buen clínico general, lo que además de crear un entendimiento más personalizado con el enfermo evita que éste rote por diversos pro­ fesionales, para cada uno de los cuales el pa­ ciente es un simple anónimo. Los procedi­ mientos tecnológicos de diagnóstico y trata­ miento deberán ser cada vez más simples, mas baratos y menos agresivos, respetando la inte­ gridad corporal del enfermo. Con Irwin y colaboradores se investigó la relación entre el mundo subjetivo del hombre, su sistema inmunológico y la posibilidad de contraer enfermedades debido a la baja inusita­ da de sus defensas. La reducción de las células T y NK Yla respuesta linfocítica insuficiente en personas sufridas y agobiadas, nos induce a tratar con especial interés las enfermedades llamadas psico-somáticas, ofreciendo al pa­ ciente mayor atención y ayuda con el propósito de robustecer su fe y su esperanza. Esta actitud humanizante es percibida fácilmente por quie­ nes están enfermos del alma. Para la Medicina próxima es pues, res­ ponsabilidad imperativa de las Facultades de Medicina modificar sus curricula pasados para formar buenos clínicos generales y especialis­ tas capacitados, ambos con idéntica calidad y rango, procurándoles primero una formación humanística adecuada con asignaturas básicas como historia y filosofía de la medicina, antro­ pología y bioética... Sin esta formación básica Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vol. XXVII Números l y 2: 67·(1,}, octubre del 2005 68 o el médico quedará cada vez mas despersonali­ zado faltando a los preceptos de su juramento de dignidad, honestidad y conciencia; sin el in­ terés de ser primero un amigo confidente del enfermo y compartir luego juntos la lucha con­ tra la enfermedad. Es imperativo de las instituciones em­ pleadoras considerar para sus colaboradores médicos, ambientes adecuados y salarios justos que se compadezcan con sus exigencias socia­ les. Establecer legislaciones justas de la llama­ da "mala practica" , muchas de las cuales se en­ cuentran al arbitrio radical de las leyes y en las mas de las veces, lejos de controlar la idonei­ dad de la práctica médica se dejan seducir por intereses exclusivamente económicos. Nunca será edificante ni positiva la actividad de un profesional que labore con horarios reducidos y bajo la presión de amenazas jurídicas. Desde Lían Entralgo has.ta Werner Jaeger sabemos que la medicina comprende al hombre entero, es pues un saber antropológico amplio y complicado, un conocimiento del hombre que integra la anatomía, la biofísica y la bioquími­ ca: un conocimiento científico y filosófico del hombre. La medicina - se dice - es la podero­ sa fuerza formativa del individuo y la colectivi­ dad, que intenta extraer de ellos, por la educa­ ción, su contenido verdadero, estimulando los elementos fundamentalmente del ser humano. El médico aparece, según Jaeber, "como repre­ sentante de una cultura especial del mas alto re­ finamiento metódico y es, al propio tiempo, la encamación de una ética profesional ejemplar por la proyección del saber sobre un fin ético de carácter práctico.." "El neoliberalismo ha atentado contra la humanidad.." Se mencionó en la Declaración del 11 En­ cuentro Mundial por la Humanidad, en Puerto Serrano. Se concluyó que la flexibilización del mercado laboral no crea mas trabajo, mas bien crea el desempleo e incrementa la precariza­ cion laboral, una situación cercana al caos que involucra a todas las fuerzas económicas y so­ ciales incluidos los servidores de la salud. El capital monopólico de los países del Norte li­ mito la circulación internacional de la fuerza del trabajo de los inmigrantes del Sur, creando además su exclusión social. Se deshumaniza­ ron así no solo la Medicina sino todos los estra­ tos de la comunidad, por eso se habló también sobre la devastación de la tierra, el exterminio de los campesinos y de las poblaciones indíge­ nas, el mercado de menores, la segregación de los viejos y el menosprecio a los enfermos. El dudoso matrimonio de la oferta y la demanda de un mercado libre sirve a los poderosos, cas­ tiga los pobres y genera una economía de es­ peculación - menciona Eduardo Galeano-. El mercado libre, humilladero de los débiles, es el mas exitoso producto de exportación de los fuertes; se fabrica para el consumo de los países pobres y ningún país rico la ha usado jamás. Los países del Sur han sido hasta ahora incapa­ ces de crear una tecnología de los poderosos que castiga y desplaza a las materias primas na­ turales - afirma a este pensador sureño. Es indudable que estas nuevas normas de libre mercado trastocaron los conceptos del ju­ ramento medico enunciados hasta 2.500 años allá en el mar Egeo. Y es evidente el cambio que estas nuevas fuerzas económicas ejercieron sobre los enunciados del humanismo renacen­ tista de Erasmo, Maquiavelo y Moro. Si se pre­ tende someter al médico como actor principal del la "deshumanización de la medicina" , debe­ ría considerarse su origen y su formación uni­ versitaria, las oportunidades de trabajo y las condiciones en que desarrollo su actividad pro­ fesional, las relaciones con el Estado o la em­ presa empleadora y, lo que es mas importante, el sistema socioeconómico que en estas últimas décadas prevaleció a manera de mordaza en la relación humana médico - paciente. Resulta comprensible y las leyes de libre mercado pueden dar cuenta, de los progresos de la quimioterapia para prolongar la vida y de la cirugía cosmética para embellecerla, pues son producto de su creación. En cambio permane­ cen como hechos inefables y nos preguntamos, que diría Hipócrates sobre las fronteras tecno­ lógicas de la fertilización in vitro y el alquiler de úteros, cuál sería la apreciación de Galeno Revista Ecuatoriana de Medicina y Ciencias Biológicas· Vol. XXVD Números 1 y 2: 68-69. octubre del 2005 Deshumanización de la medicina y bioética sobre los transplantes de órganos, bancos de es­ permas y compra-venta de partes del cuerpo; y cuál será la respuesta del Creador ante los pri­ meros experimentos de la clonación humana y el afán de perpetuidad de los hombres. Oportu­ no e recordar el mensaje de Confucio a sus dis­ cípulos: 69 "Si todavía no sabemos servir a los hom­ bres, no sabemos aún de la vida. Si desconoce­ mos la manera de servir a los espíritus, desco­ nocemos aún de la muerte". o Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vo\. XXVII Números 1 y 2: 69-69. octubre del 2005 D HOJAS DE VIDA ....................................---............ ...----------if---------------------------------------------------------------------­ Se presentarán las hojas de vida de los miembros de la sección, empezando por la directiva. Dr. Plutarco Naranjo Vargas DATOS PERSONALES: NOMBRES Y APELLIDOS: LUGAR y FECHA DE NACIMIENTO: ESTADO CIVIL: PROFESION: PLUTARCO NARANJO VARGAS Ambato Provincia del Tungurahua, Ecuador, 18 de junio de 1921. Casado 3 hijos. Doctor en Medicina, Universidad Central del Ecuador 1949. DATOS PROFESIONALES y DOCENTES: Estudios de Post-grado en la Universidad de U­ tah, Estados Unidos. Profesor agregado de Botánica y Materia Médi­ ca, 1945-1948 Universidad Central. Profesor Titular de Farmacología, 1948-1970 Universidad Central. Profesor contratado por la Universidad del Va­ lle (Cali-Colombia) para organizar el La­ boratorio y dirigir la cátedra de Farmaco­ logía y asesor del Departamento de Fisio­ logía, 1952-1956. Subdirector de la Universidad Popular 1948 1949, Universidad Central. Director del Instituto de Ciencias Naturales, 1957 - 1969, Universidad Central. Director del Centro de Biología, 1962 1963, Universidad Central. Profesor de los Cursos de Post-Grado de Medi­ cina, Universidad Central, desde 1977 en temas de Alergología. Jefe del Departamento de Ciencias Fisiológicas de la Facultad de Medicina, Universidad Central,1965-1970. Director del área de Salud, Universidad Andina Simón Bolívar, 1996. Profesor de Historia de la Medicina. Universi­ dad Politécnica Equinoccial 2002-2004. POSICIONES OFICIALES: Director General de los Servicios Médicos del Seguro Social Ecuatoriano, 1963-1966. Ministerio de Salud Pública, 1988-1992. Presidente del Comité Ejecutivo de la Organi­ zación Panamericana de la Salud 1988. Presidente de la XIII Reunión de Ministros de Salud del Área Andina, 1988. Presidente de la XLIII Asamblea Mundial de la Salud, 1990. POSICIONES ACADEMICAS. Presidente de la Academia Ecuatoriana de Me­ dicina, 1981-1983. Presidente de la Asociación Latinoamericana de Academia de Medicina, 1982-1984. Miembro del Número de de la Academia de Medicina de Chile. Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vol. XXVII Números 1 y 2: 71-73, octubre del 2005 D 72 Miembro Correspondiente de la Academia de Medicina de Venezuela, 1989. Miembro de Número de la Academia Ecuato­ riana de Historia, 1990. Miembro de la Academia de Ciencia de New York. Vicepresidente de la Casa de la Cultura Ecuato­ riana 1962-1966. Miembro del "Grupo América" 1998. Presidente del Grupo América 2002-2003. Director de la Academia Nacional de Historia del Ecuador. 1999-200 l. Director Vitalicio Honorario de la Academia N acional de Historia del Ecuador, 2002. Profesor Honorario de la Universidad Técnica Equinoccial (Quito).2oo3. Además, Miembro Honorario de varias Socie­ dades Científicas Médicas y de Alergia. Premio Interamericano de Salud Pública "Abraham Horwitz",1993. Condecoración al Mérito en el grado "Gran­ cruz" Convenio Andino Hipólito Una­ nue,1993. Placa de la Sociedad Latinoamericana de Aler­ gia e Inmunología México 1997. Placa de homenaje de la Sociedad Latinoameri­ cana de Farmacología. Montevideo-Uru­ guay. 1990. Declarado "Héroe de la Salud Pública" por la OPS/OMS.2ool. Condecoración al ejercicio médico ético. Club Rotario del Ecuador.2oo3. Condecoración al mérito. Gobierno Provincial de Pichincha.2004. OTRAS POSICIONES: POSICIONES DIPLOMATICAS: Embajador ante la Unión Soviética y Embaja­ dor Concurrente ante la República Popu­ lar de Polonia y de la República Popular de Alemania, 1977-1978. Miembro de la Junta Consultiva del Ministerio de Relaciones Exteriores, 1986-1988. PREMIOS Y CONDECORACIONES: D Premio Universidad Central por obras de inves­ tigación científica (cuatro ocasiones). Condecoración "Pluma de Oro" del Municipio de Ambato. Condecoración del Gobierno de Italia, 1972. Condecoración del Gobierno de Rumania, 1976. Premio Nacional de Ciencias, 1976. Premio "Tobar", Municipio de Quito, 1977. Condecoración Nacional "Eugenio Espe­ jo",1987. Condecoración al Mérito Nacional en Sa­ lud.1987. Condecoración "Daniel Carrión" del Gobierno del Perú, 1990. Miembro del Comité Ejecutivo de la Fundación Internacional Darwin. Miembro de la Comisión Nacional de UNES­ CO, 1969-1973. Miembro del Comité Asesor de la Oficina Sani­ taria Panamericana, para la selección de textos de enseñanza médica. Vicepresidente de la Sociedad Internacional de Quimioterapia, 1962-1972. Vicepresidente del Congreso Internacional so­ bre Quimioterapia Stuttgart (Alemania), 1963. Presidente del 11 Congreso Latinoamericano de Farmacología, 1965-1969. Presidente del 11 Congreso Latinoamericano de Alergología. Quito 1967. Presidente de la Sociedad Latinoamericana de Alergología 1967-1969. Presidente del IV Congreso Italo-Latinoameri­ cano de Etnomedicina. Quito, 1995. Presidente del V Congreso Italo-Latinoameri­ cano de Etnomedicina. Roma, 1996. Conferenciante de varias Universidades Lati­ noamericanas y Norteamericanas y Euro­ peas. Relator y participante de numerosos congresos internacionales. Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vol. XXVII Números 1 y 2: 72-73, octubre del 2005 D 73 PUBLICACIONES: IDIOMAS: 37 libros o folletos, como único autor. 59 como coautor, en libros de varios autores. Más de 300 estudios o artículos en revis­ tas nacionales, latino, norteamericanas y euro­ peas; en español, inglés, francés, italiano y ru­ so. Centenares de artículos periodísticos en "El Comercio" de Quito, y "El Universo" de Guayaquil. Castellano (Lengua materna) Inglés (fluido) Italiano (fluido). Francés (lectura y traducción). Portugués (lectura y traducción) Ruso (lectura y traducción restringida). Quichua (lectura y traducción restringida). Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vol. xxvn Números 1 y 2: 73-73, octubre del 2005 el INSTRUCTIVO PARA LOS AUTORES ----------------------------------------if---­ Revista ecuatoriana de MEDICINA y CIENCIAS BIOLÓGICAS DE LA CASA DE LA CULTURA ECUATORIANA Instructivo para los autores Hojas de Vida de los miembros de la Sección de Ciencias Naturales y Ciencias Ex­ actas de la Casa de La Cultura Ecuatoriana y de la Sociedad Ecuatoriana de Biología Núcleo de Pichincha. ! Formato del manuscrito L a revista Medicina y Ciencias Bi­ ológicas es el órgano oficial de divulgación científica de la Sección de Ciencias Naturales, Biológicas y Ciencias Exactas de la Casa de la Cultura Ecuatoriana y de la Sociedad Ecuatori­ ana de Biología, Núcleo de Pichincha. Sus ob­ jetivos son: divulgar artículos científicos, pre­ sentar casos médicos y publicar notas científi­ cas en las áreas de ciencias biológicas puras, aplicadas y biomédicas; esta revista tiene tres secciones: A) Artículos Originales, B) Traba­ jos de Revisión y Notas Científicas y C) Hojas de Vida. Los artículos originales provienen de trabajos de investigación científica realizados por los autores, es responsabilidad de ellos el uso de datos de terceros. Los artículos origi­ nales constarán de las siguientes partes: Títu­ lo, Autor(es), Afiliación Institucional, Re­ sumen, Palabras Clave, Abstract, Key- word, Introducción, Material y Métodos, Resulta­ dos, Discusión, Conclusiones, Agradecimien­ tos y Referencias Bibliográficas. Los Trabajos de revisión y notas científicas, tendrán un formato personalizado por sus autores, pero es obligatorio el uso de las siguientes secciones: Título, Autor(es), Afiliación Institucional, Texto y Referencias Bibliográficas. I 1. Los manuscritos deben ser presentados, en idioma españolo inglés, en papel A4, con los cuatro márgenes de 2.5cm, tipo de letra ''Times'' tamaño 12, y páginas numeradas. 2. Título en minúsculas, los nombres científi­ cos mantendrán estrictamente las normas de nomenclatura biológica, con una exten­ sión ideal de 15 palabras, negrita, centra­ do, 18 puntos. Ejemplo: Preferencia de estratos boscosos en la alimentación de Turdus fuscater del Pasochoa 3. Autor o Autores: nombre(s), apellido(s), negrita, centrado, 12 puntos. Ejemplo: Juan Eduardo Pueblo P.I & Estelita Estrella2 4. Afiliación institucional: Institución, Ciu­ dad-País, correo electrónico del autor en­ cargado de la correspondencia, si los au­ tores representan a diferentes institu­ ciones indicar con un numeral, 9 puntos. Ejemplo: Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vol. XXVII Números l y 2: 75-77, octubre del 2005 D 76 1. Centro de Investigaciones Científi­ cas del lASA, Facultad de Ciencias Agropecuarias, ESPE, Sangolquí­ Ecuador. [email protected] 2. Departamento de Zoología, Escuela de Biología, Universidad Amazónica del Uruguay. D 5. RESUMEN con una extensión máxima de 200 palabras. Palabras clave: cinco palabras separadas por coma, la primera letra en mayúscula, en orden alfabético. 6. ABSTRACT (Título en inglés) traducción del resumen, Keywords: palabras clave traducidas al idioma inglés. 7. Como se nota de los puntos cinco y seis los títulos de las secciones van en mayúscu­ las, negrita y centrados, 12 puntos. 8. Texto con letra normal de 12 puntos. Si se requiere un subtítulo este va en el mismo párrafo con negrita, separado del texto por punto y raya. 9. Las figuras (esquemas, diagramas, dibu­ jos, gráficos, fotografías y mapas) deben s­ er preparadas en Photoshop, y presentase en hoja aparte y numeradas, con el título al pie de la figura en 9 puntos. La ubicación de las figuras deben señalarse con lápiz en el margen derecho del manuscrito, el área útil de la página es de 16.5 x 24 cm. El Comité Editorial se reserva el derecho de ubicar las figuras, en función del espacio disponible, o solicitar a los autores hacer los cambios correspondientes. 10. Las tablas deben presentarse en Excel, con el título en la parte superior, de un mo­ do conciso y explicativo, en 9 puntos. 11. REFERENCIAS BIBLIOGRÁFICAS se prefieren revisiones que provengan de artículos publicados en revistas científicas indexadas, libros reconocidos, tesis de doc­ torado, maestría o pre-grado que reposen en las bibliotecas de las instituciones otor­ gantes. Los artículos en revisión o en pren­ sa, así como las comunicaciones person­ ales se citarán en el texto, pero no formarán parte de la lista bibliográfica. Las referen­ cias, deben constar en el texto numeradas entre paréntesis y debeb aparecer en la bibliografía de acuerdo a la secuencia de citación. Use sangría especial francesa, 9 puntos. Ejemplos de referencias bibliográ­ ficas: a) ARTÍCULO DE REVISTA CIENTÍFICA DUCHICELA, E. J. l. 2004. Diversidad funcional de las micorrizas arbuscu­ lares asociadas a Brosimun utile en condiciones naturales. Ciencia 7(8): 65-77. STAIKOU, A. & LAZARIDOU-DIMI­ TRlADOU, M. 1991. The life cy­ cle,population dynarnics, growth and secondary production of the snail Xe­ ropicta arenosa Ziegler (gasteropo­ da:Pulmonata) in northem Greece. Zoological Joumal of Linnean Soci­ ety, London, 101: 179-188. DYKE, F. G. V.; BROKE, R. H. & SHAW, H. G. 1986. Use of road track count as indices of mountains lion presence. J ournal of wildlife Management, Bethesda, 50(1):102-109. b) CAPíTULO DE LIBRO EISENBERG, J. F. 1979. Habitat, eco no­ my and society: some correlations, and hypotheses for the Neotropical primates. En: Bemsteind, I.S and O.E. SMITH. eds. Primates ecology and human origins: ecologycal influ­ ences on social organization: 215­ 262. Granland STPM Press. NY y London. c) TESIS DE GRADO JIMÉNEZ P., P. J. 1995. Área de vida y u­ so preferencial del hábitat de Cebus albifrons (Primates: Cebidae) en Cuyabeno, Amazonía Ecuatoriana. Tesis de Licenciatura, Pontificia Uni­ versidad Católica del Ecuador. Quito, Ecuador. Revista Ecuatoriana de Medicina yCiencias Biológicas - Vol. xxvn Números 1 y2: 76-77, octubre del 2005 D 77 d) LIBROS ENTEROS KREBS, C. H. 1985. Ecología: estudio de la distribución y abundancia. Segun­ da edición. Hada. México. e) INFORMACION PROVERNIENTE DE INTERNET Ciertas páginas que tengan infonnación científica relevante pueden citarse así: DI flORE, A. 2001. Proyecto Primates E­ cuador. Página de Internet: www.nyu.edulprojects/difiorelYasun. Consultada 13-Enero-2005. En la presentación de casos, las referen­ cias bibliográficas deben ir de acuerdo a la con­ notación editorial internacional que consta en la página web: http://www.icmje.org/. El autor principal deberá enviar el manu­ scrito original (más dos impresiones y una copia electrónica del documento en CD bajo el fonnato Word 3000 o inferior), acompañado de una carta dirigida al Editor de la Revista, in­ dicando la originalidad del trabajo. Los manu­ scritos serán revisados por dos árbitros, su aprobación estará sujeto al contenido científi­ co, respaldado por el parecer de los árbitros y del comité Editorial. Se solicitará a los autores cambios mediante la devolución de los manu­ scritos, acompañados de las sugerencias. Para la presentación de casos y notas científicas y hojas de vida dirigirse al Dr. Claudio Cañizares, Editor de la Revista Medicina y Ciencias Biológicas de la Casa de la Cultura E­ cuatoriana. Apartado 17-08-8339 Quito. Telé­ fonos 2464261, 2520572, 098747558, correo electrónico: [email protected] y para los tra­ bajos científicos a la Dra. Violeta Rafael Presi­ denta de la SEB-PICHINCHA, Escuela de Ciencias Biológicas, Pontificia Universidad Católica del Ecuador. Apartado Postal 17-01­ 2184, Quito, Ecuador, correo electrónico: vrafael @puce.edu.ec :l Revista Ecuatoriana de Medicina y Ciencias Biológicas - Vol. XXVII Números 1 y 2: 77-77, octubre del 2005 el Re'IIista Ecuatoriana de MEDICINA y CIENCIAS BIOLÓGICAS VOLUMEN XXVI - NÚMEROS 1 Y 2 se tenninó de imprimir en Quito, el mes de octubre del 2005, en la Editorial "Pedro Jorge Vera" de la Casa de la Cultura Ecuatoriana "Benjamín Camón ", siendo Presidente el Dr. Marco Antonio Rodríguez, Director de Publicaciones el Lic. Marcelo Cevallos R. y Jefe Técnico General el Sr. Guillermo Arias G. Publicación del Fondo Editorial de la CASA DE LA CULTURA ECUATORIANA