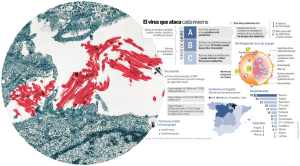
EVALUACIN DE LA CAPACIDAD PROBIOTICA in vitro DE UNA
Anuncio

EVALUACIÓN DE LA CAPACIDAD PROBIOTICA in vitro DE UNA CEPA NATIVA DE Saccharomyces cerevisiae Angela Rocio Ortiz Camargo Joanna Isabella Reuto Andrade Pontificia Universidad Javeriana Facultad de Ciencias Microbiología Industrial y Microbiología Agrícola y Veterinaria Bogota D. C. Julio de 2007 EVALUACIÓN DE LA CAPACIDAD PROBIOTICA in vitro DE UNA CEPA NATIVA DE Saccharomyces cerevisiae Angela Rocio Ortiz Camargo Joanna Isabella Reuto Andrade TRABAJO DE GRADO para optar por el titulo de Microbióloga Industrial y Microbióloga Agrícola y Veterinaria Microbióloga Industrial ANDREA AGUIRRE Bacterióloga, MSc. Director GUSTAVO ARBELAEZ Médico Veterinario, MSc. Ph.D Codirector Pontificia Universidad Javeriana Facultad de Ciencias Microbiología Industrial y Microbiología Agrícola y Veterinaria Bogota D. C. Julio de 2007 EVALUACIÓN DE LA CAPACIDAD PROBIOTICA in vitro DE UNA CEPA NATIVA DE Saccharomyces cerevisiae Angela Rocio Ortiz Camargo Joanna Isabella Reuto Andrade ANDREA AGUIRRE Bacterióloga, MSc. Director GUSTAVO ARBELAEZ Médico Veterinario, MSc. Ph.D Codirector ANA KARINA CARRASCAL Bacterióloga, MSc. Jurado ADRIANA PULIDO Bacterióloga, MSc. Jurado Pontificia Universidad Javeriana Facultad de Ciencias Microbiología Industrial y Microbiología Agrícola y Veterinaria Bogota D. C. Julio de 2007 EVALUACIÓN DE LA CAPACIDAD PROBIOTICA in vitro DE UNA CEPA NATIVA DE Saccharomyces cerevisiae Angela Rocio Ortiz Camargo Joanna Isabella Reuto Andrade APROBADO ANGELA UMAÑA MUÑOZ, M Phill. Decana Académica Facultad de Ciencias LUIS DAVID GOMEZ, MSc. Director de carreras de Microbiología Pontificia Universidad Javeriana Facultad de Ciencias Microbiología Industrial y Microbiología Agrícola y Veterinaria Bogota D. C. Julio de 2007 NOTA DE ADVERTENCIA “La Universidad no se hace responsable por los conceptos emitidos por sus alumnos en sus trabajos de tesis. Solo velará por que no se publique nada contrario al dogma y a la moral católica y por que las tesis no contengan ataques personales contra persona alguna, antes bien se vea en ellas el anhelo de buscar la verdad y la justicia”. Artículo 23 de la Resolución N° 13 de Julio de 1946 AGRADECIMIENTOS A la Doctora Andrea Carolina Aguirre por la confianza y el apoyo incondicional que nos brindo durante la elaboración de este proyecto. A la Doctora Balkys Quevedo por su paciencia, disposición y colaboración. Al laboratorio de Biotecnología Aplicada, a su directora Aura Marina Pedroza por permitirnos llevar a cabo la realización de este proyecto. A nuestros compañeros de laboratorio por su amistad y por la ayuda brindada. Agradecimiento especial a los laboratorios de virología, inmunología, monitoria, contaminación de aguas, microbiología especial, microbiología ambiental y de alimentos por el préstamo de equipos y por la colaboración brindada. A la Vicerrectoría académica de la Pontificia Universidad Javeriana por la financiación de este proyecto. A la Pontificia Universidad Javeriana y sus docentes por la formación integral brindada y por permitirnos vivir una de las experiencias más gratificantes de nuestras vidas. A Dios por haberme bendecido en estos años y por permitirme cumplir con este sueño de mi vida. A mi madre, mi abuelita Amelia y mi tío Jesús por el apoyo incondicional, el cariño, la confianza que depositan en mi cada día y todo lo demás que me han brindado, porque sin ustedes no sería ni la mitad de ser humano que he llegado a ser. A mis amigos por su apoyo, por alegrar mis días, por inspirarme tantas cosas y por su compañía durante estos años maravillosos. Angela Rocío Dedico este trabajo primeramente a Dios, fuente de vida, de sabiduría y de hermosas bendiciones. A mi Madre por ser mi guía, mi sostén incondicional, mi apoyo y mi consejera, a mi hermana por ser mi más fiel amiga, a toda mi familia por estar siempre a mi lado sustentándome y ayudándome a ser el ser humano que hoy soy. A Andrés por enseñarme a amar sinceramente y apoyarme sin condiciones. A mis amigas y amigos por todos los gratos momentos que hemos compartido y por arrancar de mis labios mil sonrisas. Joanna Isabella TABLA DE CONTENIDO Página 1. INTRODUCCION 16 2. MARCO TEORICO 2.1. PROBIÓTICOS 17 2.1.1. Probiosis 17 2.1.2. Mecanismos implicados en la probiosis 18 2.1.3. Composición y administración de probióticos 19 2.1.4. Obtención y conservación de cepas probióticas 20 2.1.5. Pruebas de capacidad probiótica in vitro 21 2.1.5.1. Tolerancia a sales biliares 22 2.1.5.2. Tolerancia a pH 22 2.1.5.3. Resistencia a jugos gástricos 23 2.1.5.4. Adhesión celular 23 2.1.5.4.1. Cultivos celulares 24 2.1.5.4.2. Líneas celulares 25 2.1.5.4.3. Ensayos de adhesión 26 2.1.5.5. Reducción del colesterol en presencia de sales biliares 27 2.1.6. Resistencia del probiótico a condiciones in vivo 28 2.1 7. Procesamiento de probióticos en productos terminados 28 2.2. PROBIÓTICOS PARA AVES DE CORRAL 29 2.2.1. Aves de corral y tracto gastrointestinal 29 2.2.2. La flora normal intestinal de aves de corral 31 2.2.3. Infecciones bacterianas tempranas en polluelos 31 2.2.4. Aplicación y evaluación de probióticos en aves de corral 32 2.3. Saccharomyces cerevisiae 33 2.3.1. Generalidades 33 2.3.2. Ciclo sexual y asexual 35 2.3.3. Cinética de crecimiento 35 2.3.4. Antecedentes de Saccharomyces cerevisiae como probiótico 36 2.4. INDUSTRIA AVÍCOLA EN COLOMBIA 40 3. FORMULACION DEL PROBLEMA Y JUSTIFICION 42 3.1. Formulación del problema 42 3.2. Justificación 43 4. OBJETIVOS 44 ix 4.1. Objetivo general 44 4.2. Objetivos específicos 44 5. MATERIALES Y METODOS 45 5.1. Métodos 45 5.1.1. Obtención y conservación de Cepa A y Cepa B 45 5.1.2. Determinación de capacidad probiótica in vitro 45 5.1.2.1. Tolerancia a sales biliares 45 5.1.2.2. Tolerancia a pH 46 5.1.2.3. Determinación de resistencia a jugos gástricos 46 5.1.2.4. Reducción del colesterol en presencia de sales biliares 47 5.1.2.5. Prueba de adherencia 49 6. RESULTADOS Y DISCUSIÓN DE RESULTADOS 50 6.1. Control de calidad bancos cepa A y cepa B 50 6.2. Tolerancia a sales biliares 52 6.3. Tolerancia a rangos de pH 55 6.4. Resistencia a jugos gástricos 59 6.5. Reducción de colesterol en presencia de sales biliares 63 6.5.1. Determinación de reducción Colesterol en presencia de sales biliares 64 6.6. Ensayo de adhesión a células Caco – 2 69 7. CONCLUSIONES 76 8. RECOMENDACIONES 78 9. BIBLIOGRAFIA 79 10. ANEXOS 94 x INDICE DE FIGURAS Página Figura Nº 1. Células Caco – 2 en monocapa con coloración de Wrigth 26 Figura Nº 2. Análisis de adherencia por microscopio invertido de Kluyveromyces lactis a células Caco-2 diferenciadas con coloración de Giemsa 26 Figura Nº 3. Microscopia electrónica de formación de yemas de Saccharomyces cerevisiae 35 Figura Nº 4. Reproducción sexual y asexual de levaduras 36 Figura Nº 5. Curva de crecimiento de levaduras 36 Figura Nº 6. Consumo per cápita de pollo en Colombia (1970 – 2006) 40 Figura Nº 7. Producción de pollo (toneladas) en 2001 – 2006 41 Figura Nº 8. Fórmula porcentaje de asimilación de colesterol en muestras de medio YPGCHO 48 Figura Nº 9. Fórmula Recuento (Levaduras/mL) en cámara de Newbauer 49 Figura Nº 10. Aspecto microscópico de la cepa nativa (estudio) de Saccharomyces cerevisiae Figura Nº 11. Curva de viabilidad expresada en recuento (UFC/mL) 50 de controles de calidad para cepa A (estudio) y cepa B (control) 50 Figura Nº 12. Recuentos (UFC/mL) en sales biliares para cepa A (estudio) y cepa B (control) 53 Figura Nº 13. Recuentos (UFC/mL) en valores de pH de 2.0 – 5-0 para cepa A (estudio) y cepa B (control). 57 Figura Nº 14. Recuentos (UFC/mL) en Jugos Gástricos de pH 2.0 – 2.3 para cepa A (estudio) y cepa B (control) 60 Figura Nº 15. Recuentos (UFC/mL) en Jugos Gástricos de pH 7.0 – 7.2 para cepa A (estudio) y cepa B (control) 62 Figura Nº 16. Curva patrón de colesterol 63 Figura N. 17. Reacción de Liebermann – Burchard para determinación de Colesterol 67 Figura Nº 18. Recuentos (Levaduras/mL) de cepa A (estudio) expuesta a sonicación 68 Figura Nº 19. Recuentos (Levaduras/mL) de cepa B (control) expuesta a sonicación 69 Figura Nº 20. Células Caco – 2 en monocapa con coloración de Wrigth 70 Figura Nº 21. Análisis de adherencia por microscopio invertido de Saccharomyces cerevisiae a células Caco-2 con colorante de Wright xi 72 Figura Nº 22. Análisis de adherencia por microscopio invertido de S. cerevisiae var. boulardii a células Caco-2 con colorante de Wright 72 Figura Nº 23. Gráfica de barras de Levaduras adheridas/ 100 células Caco -2 de la cepa A (estudio) y Cepa B (control) xii 73 INDICE DE TABLAS Página Tabla Nº 1. Coloración de Gram y valores de Absorbancia620 nm de inóculos de cepa A (estudio) y cepa B (control) 53 Tabla Nº 2. Siembra en superficie en agar YGC de inóculos de cepa A (estudio) y cepa B (control) 53 Tabla Nº 3. Coloración de Gram y valores de Absorbancia620 nm de inóculos de cepa A (estudio) y cepa B (control) 56 Tabla Nº 4. Siembra en superficie en agar YGC de inóculos de cepa A (estudio) y cepa B (control) 57 Tabla Nº 5. Coloración de Gram y valores de Absorbancia620 nm de inóculos de cepa A (estudio) y cepa B (control) 60 Tabla Nº 6. Siembra en superficie en agar YGC de inóculos de cepa A (estudio) y cepa B (control) 60 Tabla Nº 7. Coloración de Gram y valores de Absorbancia 620 nm de inóculos y de cepa A (estudio) y cepa B (control) 64 Tabla Nº 8. Siembra en superficie en agar YGC de inóculos de cepa A (estudio) y cepa B (control) 65 Tabla Nº 9. Valores de colesterol degradado en 12 horas de incubación por la cepa A (estudio) y la cepa B (control) 65 Tabla Nº 10. Porcentajes de asimilación de colesterol/12 horas de incubación por la Cepa A (estudio) y la Cepa B (control) 66 Tabla Nº 11. Coloración de Gram y valores de Absorbancia 620 nm de inóculo y Cepa A (estudio) y cepa B (control) 70 Tabla Nº 12 .Siembra en superficie en agar YGC de inóculos de cepa A (estudio) y cepa B (control) 70 xiii INDICE DE ANEXOS Página ANEXO Nº 1. ASPECTO MACROSCÓPICO DE S. cerevisiae y S. cerevisiae var. boulardii 93 Anexo 1.1. Aspecto macroscópico de S. cerevisiae 93 Anexo 1.2. Aspecto macroscópico de S. cerevisiae var. boulardii 93 Anexo 1.3. Recuento en microgota de S. cerevisiae en Agar YPG 94 ANEXO Nº 2. TABLAS DE RESULTADOS OBTENIDOS PARA CONTROL DE CALIDAD DE BANCOS DE CEPA A Y B 94 Anexo 2.1. Coloración de Gram de inóculos y colonias de cepa A (estudio) y cepa B (control) 94 Anexo 2.2. Técnica: Siembra en superficie en agar YPG de control de calidad de Cepa A (estudio) y Cepa B (control) 95 ANEXO Nº 3. TABLA RESULTADOS DE PRUEBA DE TOLERANCIA A SALES BILIARES 95 ANEXO Nº 4. TABLA RESULTADOS DE PRUEBA DE TOLERANCIA A RANGOS DE pH 96 ANEXO Nº 5. TABLA RESULTADOS DE PRUEBA DE RESISTENCIA A JUGOS GÁSTRICOS 96 ANEXO Nº 6. TABLAS Y FIGURAS DE PRUEBA DE DETERMINACIÓN DE REDUCCION DE COLESTEROL EN PRESENCIA DE SALES BILIARES 97 Anexo 6.1. Protocolo de elaboración de curva patrón de colesterol 97 Anexo 6.2. Curva patrón 97 Anexo 6.3. Valores de Absorbancia (550 nm) obtenidos en la determinación de reducción de colesterol del medio YPGCHO por las cepa A y Cepa B 98 Anexo 6.4. Medio YPGCHO inoculado con la cepa A (estudio) después de 12 horas de incubación 98 Anexo 6.5. Medio YPGCHO inoculado con la cepa B (control) después de 12 Horas de incubación 99 Anexo 6.6. Separación de fases orgánica y acuosa en la extracción de colesterol de las muestras (cepa A y B) 99 Anexo 6.7. Reaccion de Liebermann - Burchard para la determinación de colesterol en medio YPGCHO (cepa A) 100 Anexo 6.8. Reaccion de Liebermann - Burchard para la determinación xiv de colesterol en medio ypgcho (cepa B) 100 ANEXO 7. FIGURAS DE ENSAYO DE ADHESION A CÉLULAS Caco – 2 101 Anexo 7.1. Análisis de adherencia por microscopio invertido de Saccharomyces cerevisiae a células Caco-2 con colorante de Wright 101 Anexo 7.2. Análisis de adherencia por microscopio invertido de Saccharomyces cerevisiae var. boulardii a células Caco-2 con colorante de Wright 101 ANEXO 8. COMPOSICION DE MEDIOS DE CULTIVO Y REACTIVOS 102 Anexo 8.1. AGAR YPG 102 Anexo 8.2. CALDO YGC 102 Anexo 8.3. CALDO YPGCHO 103 Anexo 8.4. Reactivo de extracción de colesterol 103 Anexo 8.5. Medio Mínimo Esencial Eagle 103 Anexo 8.6. PBS 104 ANEXO 9. PROTOCOLO DE TRIPSINIZACION DE CÉLULAS Caco-2 104 ANEXO 10. TINCIÓN DE WRIGHT 104 ANEXO 11. ANÁLISIS ESTADÍSTICO 104 Anexo 11.1. Prueba de tolerancia a sales biliares 104 Anexo 11.2. Prueba de tolerancia a pH 105 Anexo 11.3. Prueba de tolerancia a jugos gástricos 105 Anexo 11.4. Prueba de reducción del colesterol en presencia de sales biliares 106 xv RESUMEN Los probióticos, son microorganismos vivos usados como suplementos alimenticios los cuales afectan positivamente al huésped al mejorar el equilibrio microbiano en el tracto gastrointestinal contribuyendo a la supresión del crecimiento de patógenos por competencia por espacio y sustrato, modulando el sistema inmune, inhibiendo metabolitos tóxicos producidos por otros microorganismos, transformando compuestos carcinogénicos, aumentando la asimilación de nutrientes, entre otros. A través del tiempo se ha comprobado que han desempeñado un papel muy importante en el control de enfermedades sin producir efectos negativos. En el presente estudio se evaluó la capacidad probiotica in vitro de una cepa nativa de Saccharomyces cerevisiae comparada con una cepa comercial utilizada como probiótico. Se realizaron pruebas “in vitro” como resistencia a sales biliares, tolerancia a rangos de pH y jugos gástricos, donde no se observaron diferencias al medir el crecimiento entre la cepa de estudio A y la cepa control B en los diferentes muestreos; reducción del colesterol en presencia de sales biliares en la cual luego de 12 horas de incubación de la levadura en medio con colesterol se obtuvo una reducción del 54% para la cepa A y 58% para cepa B y prueba de adherencia en la que las dos cepas se consideraron adherentes a las células Caco-2. Con los resultados obtenidos se concluye que la cepa A presenta las características apropiadas para ser empleada como probiótico en nutrición animal. 1. INTRODUCCION Los probióticos, son suplementos alimenticios a base de microorganismos vivos que benefician al huésped mejorando el equilibrio microbiano intestinal, contribuyendo a la supresión del crecimiento de patógenos y favoreciendo los procesos metabólicos del mismo. A través del tiempo se ha comprobado que desempeñan un papel muy importante en el control de enfermedades sin producir efectos negativos. Dentro de los microorganismos probióticos más estudiados, se encuentran las Bacterias Ácido Lácticas y Bifidobacterium, ya que se ha comprobado su efecto benéfico en la salud humana y animal, representando el 90% de la flora intestinal. Por esta razón, se han usado ampliamente para reducir los trastornos de origen intestinal en el hombre, como lo reportó Metchnikoff en 1970. Adicionalmente, se han reportado estudios en especies animales como terneros, cerdos, aves y conejos, entre otros, que resaltan la importancia de la administración de estos microorganismos, al obtener resultados positivos y benéficos en los animales en estudio. Saccharomyces cerevisiae ha sido estudiada como integrante de la dieta en pollos de engorde, demostrando que su uso frecuente reduce la carga de enteropátogenos como Salmonella spp y E. coli y en menor proporción de Pseudomonas sp, S. aureus, y de virus, parásitos y hongos. Esto lo consigue, al corregir el balance de la población microbiana, mejorar la asimilación de nutrientes generando una ganancia de peso importante en estos animales de engorde y al tener una acción estimulante de la inmunidad del animal, por lo que su uso como probiótico es útil y prometedor. En el presente estudio se evaluó la capacidad próbiotica de una cepa nativa de Saccharomyces cerevisiae por medio de pruebas in vitro como tolerancia a concentraciones de sales biliares, determinación de resistencia a jugos gástricos, tolerancia a rangos de pH, reducción de colesterol en presencia de sales biliares y pruebas de adherencia en células Caco-2. 16 2. MARCO TEORICO 2.1 PROBIÓTICOS El término probiótico, es derivado del significado griego ¨ Para la vida ¨ y ha tenido diversos significados a través de los años. Primero fue usado por Lilley y Stillwell en 1965 para describir sustancias secretadas por un microorganismo que estimulaba el crecimiento de otro. Esta definición no persistió y fue subsecuentemente usado por Sperti en 1971 para describir extractos de tejido que estimulaban el crecimiento microbiano. No fue, sino hasta 1974 que Parker los definió como: Organismos y sustancias que contribuyen al balance microbiano intestinal. Fuller en 1989 redefinió los probióticos como “Suplemento alimenticio microbiano vivo que afecta benéficamente al animal huésped mejorando su balance microbiano intestinal”. En 1992, Havenaar y Huis In’t Veld consideraron los próbioticos como aquellos que al ser suministrados a animales o humanos, producen efectos benéficos en el hospedero por el incremento de las actividades de la microbiota nativa. Con base en los recientes avances en este campo, Salminen et al (1999) propusieron una definición mas apropiada: Los probióticos son preparaciones o componentes de células microbianas que tienen un efecto benéfico en la salud del hospedero. Sin embargo, considerando las definiciones anteriores, Schrezenmeier y De Vrese (2001) proponen la definición: “ Preparación de un producto que contiene microorganismos viables en suficiente número, los cuales alteran la microflora (por implantación o colonización) en un compartimiento del huésped provocando efectos beneficiosos sobre la salud del mismo" como la más acertada para el término probiótico. 2.1.1 Probiosis La probiosis se encuentra basada en el concepto de exclusión competitiva, designada como la actividad de la flora intestinal normal para limitar la colonización intestinal por varios patógenos entéricos, al colonizar los mismos sitios del intestino que utilizarían los patógenos, mediante la administración de cultivos de microorganismos benéficos para el huésped (Jordan F. y Pattison M, 1998). Animales que eran tratados con antibióticos generalmente mostraban una resistencia reducida a la infección por patógenos intestinales. El factor protector en animales sanos es normalmente atribuido a la flora bacteriana anaerobia. Los factores asociados a la avicultura moderna, como la administración y composición de las vacunas, higiene excesiva y estrés, 17 alteran la flora intestinal normal aumentando la incidencia de infecciones. La higiene excesiva en la cría de animales, resultante por ejemplo del uso exagerado de desinfectantes, puede dificultar el desarrollo de la flora normal, permitiendo así la colonización del intestino por parte de bacterias no deseadas. Durante el estrés el número de Lactobacilos tiende a decrecer y el número de Coliformes, especialmente de Escherichia coli aumenta en las regiones altas del intestino delgado (Acuña, 1995). En estas situaciones, la restauración de la flora normal mediante el uso de probióticos es de gran valor porque minimiza desarreglos digestivos ayudando a mantener la salud, previniendo trastornos intestinales, incluso ayudando a mejorar el crecimiento y conversión de animales domésticos, mejorando el metabolismo del colesterol e inhibiendo la carcinogénesis (Fuller, 1992). 2.1.2 Mecanismos implicados en la probiosis Los probióticos pueden afectar de forma benéfica al animal modificando las interacciones metabólicas que tienen lugar en el intestino. Esto puede suceder por medio de la supresión de reacciones generadoras de metabolitos tóxicos; ejemplo de esto es la presencia de proteína no digerida o con aminoácidos no absorbidos o mal absorbidos en el intestino grueso, resultando en un cambio de la flora intestinal. Microorganismos como Enterobacterias o Clostridium sp, comienzan a dominar y a romper las proteínas generando cadaverina, putrescina, tiramina e histamina. Estos compuestos, pueden generar una irritación de la pared intestinal, produciendo un daño en el epitelio llevando a una mala absorción por lo que el valor osmótico del contenido intestinal aumenta, resultando en la atracción de líquidos por el intestino que conduce a diarrea. Adicional a lo anterior, la histamina entra a la corriente sanguínea y actúa como un tipo de veneno, ya que dosis elevadas de este compuesto en la sangre causa edemas (Otero, 1997). Cuando el probiótico se encuentra presente en el sistema digestivo se controla la presencia de esta flora dañina, evitando así la producción de estos metabolitos. Adicionalmente, los microorganismos probióticos puede potenciar reacciones de detoxificación, estimulando la digestión mediante sustitución de deficiencia de enzimas digestivas o sintetizando vitaminas u otros nutrientes no disponibles en la dieta, mejorando la absorción de los nutrientes, resultante en un aumento de peso, especialmente benéfico en animales de engorde (Fuller, 1992). Los probióticos pueden actuar benéficamente al alterar el metabolismo bacteriano intestinal directamente a través de sus propias actividades metabólicas o bien de forma indirecta desplazando o influenciando las actividades metabólicas de otros grupos microbianos, así como la propiedad de influenciar en el sistema inmune del animal, ya que el uso de los 18 probióticos es una manera de reforzar la capacidad de defensa natural de la flora bacteriana normal contra los patógenos (Fuller, 1992). 2.1.3 Composición y administración de probióticos Los probióticos pueden contener una o más cepas bacterianas. Se han utilizado Lactobacilos, Bifidobacterias, Estreptococos, Enterococos, Escherichia coli no patógenos, Pediococos, Propionibacterias, levaduras y especies de Bacillus y Leuconostoc. Tissier (1906) aisló por primera vez en el Instituto Pasteur de París, Bífidobacterias en las deposiciones de los lactantes alimentados con leche materna y estableció una relación con el hecho de que los lactantes alimentados con leche materna sólo padecían diarrea en raras ocasiones. Por ello recomendó la ingestión oral de Bífidobacterias al suponer que éstas eran capaces de eliminar las bacterias responsables de las diarreas. En cuanto a las levaduras, una de las mas estudiadas es Saccharomyces boulardii, debido a que tiene varias ventajas, como su crecimiento a una temperatura relativamente alta (37 ºC), sin que se vea afectado su metabolismo y la resistencia a varios antibióticos (Buts, 2005). Al ser administrada en forma liofilizada, se reactiva en el tracto gastrointestinal, generando una respuesta benéfica en la salud y fisiología del huésped (McFarland et al, 1993), Entre otras ventajas de este microorganismo, se encuentra el no ser tan sensible al calor y poder sobrevivir a exposiciones cortas de calor o largas de enfriamiento (Toma et al, 2005). El estudio de la capacidad probiótica de levaduras ha sido más limitado y en menor proporción (Guslandi et al, 2003), ya que Lactobacillus sp. y Bifidobacterium son parte predominante de la flora normal del tracto gastrointestinal de humanos y otros vertebrados (Kumura et al, 2004), lo que ha generado que se realicen amplios estudios sobre estos dos géneros (Boriello et al, 2003). Saccharomyces cerevisiae ha sido estudiada como un suplemento en la dieta de animales, debido a que mejora la digestibilidad de nutrientes (Agarwal et al, 2000). Newbold et al (1995), observó que diferentes cepas de esta levadura tenían efectos benéficos en terneros actuando en las células de epitelio intestinal, al resistir factores como presencia de lisosimas, enzimas pancreáticas, bajo pH, ácidos orgánicos y sales biliares presentes en estos animales, manteniéndose metabólicamente activas. Mathew et al (1998), observó que la adición de levaduras en la dieta basal de cerdos, tendía a generar un aumento de peso en estos animales, comparado con aquellos a los que no se les suministraba. 19 Un probiótico debe ser capaz de ejercer un efecto benéfico sobre el huésped y no ser patógeno ni toxico, debe normalmente estar presente en forma viable o por lo menos como células metabólicamente activas, capaces de sobrevivir en el intestino, además debe permanecer viable y estable por largos periodos de almacenamiento. Los probióticos deben actuar produciendo compuestos antibacterianos, incluidos ácidos que reducen el pH intestinal, compitiendo por nutrientes o lugares de adhesión, alterando el metabolismo microbiano o estimulando el sistema inmune. Para que se establezcan de forma permanente, habría que administrarlos al poco tiempo de nacer. En el animal adulto los efectos tienden a durar tanto como el tratamiento, por eso el mejor método de administración es el continuo. La implantación de probióticos depende de la duración de la fase de crecimiento, la cual puede ser influenciada por la capacidad de asociación a la pared intestinal y de utilización de nutrientes disponibles. Los probióticos se pueden administrar junto con prebióticos, a menudo oligosacáridos, los cuales se cree ayudan a su crecimiento y establecimiento en el intestino (Fuller, 1992). 2.1.4. Obtención y conservación de cepas probióticas Cuando se realiza un estudio sobre obtención de nuevas cepas con capacidad probiótica, el primer paso es el muestreo, el cual debe realizarse en lugares en lo que probablemente se encuentre el microorganismo de acuerdo a su ecología y metabolismo. Una vez obtenidas las cepas aisladas del medio donde se encuentran de acuerdo a la ecología y características de crecimiento, se procede por un lado a realizar pruebas bioquímicas e incluso moleculares, que permitan caracterizar el microorganismo y por otro, realizar pases en medios adecuados según el tipo de microorganismo, que permitan obtener un cultivo axénico (Thomas, 2002). Una vez se ha obtenido el microorganismo, se debe realizar la conservación de la cepa. Los denominados bancos de cepas son un conjunto de alícuotas homogéneas de un cultivo microbiológicamente puro que se almacenan bajo condiciones que garanticen su viabilidad, pureza, actividad y características fenotípicas y genotípicas. La finalidad de estos bancos es mantener un registro detallado y actualizado de la cepa, conservar microorganismos de producción, mantener colección para estudios comparativos y disponibilidad para otros laboratorios (Cameotra, 2007) Existen varios métodos para la conservación de cepas entre los que se encuentran los métodos a corto, mediano y largo plazo. El primer método permite la conservación de la cepa por un lapso máximo de 15 días; se realizan subcultivos o pasajes celulares periódicos en caja, siendo una herramienta que garantiza un acceso inmediato al microorganismo. Sin 20 embargo, se presentan problemas como la variabilidad genética que pueden sufrir los microorganismos y contaminación, lo que aumenta el riesgo de perder la cepa. El segundo método, permite una conservación por espacio de 3 a 6 meses; los medios líquidos son preferibles a los sólidos, ya que en microorganismos como las levaduras, evitan la formación de ascosporas, que disminuyen la estabilidad de la cepa. En bacterias y levaduras, son comúnmente usados la solución salina, agua peptonada, buffer fosfato, entre otros; para hongos filamentosos y actinos, se usa compuestos como avena, arena, agua y aceite mineral (Chang et al, 1986). El tercer método permite la conservación de la cepa por un intervalo de tiempo más amplio, son los más recomendados, debido a que detiene el crecimiento de las células microbianas garantizando así al máximo su estabilidad genética. Dentro de este se encuentran los métodos de congelación o criopreservación, método simple y usado para la preservación de las cepas. Debe ser añadido un agente crioprotector (anticongelante biológico), que debe ser adicionado al cultivo y cuya finalidad es reducir la injuria de las células al no ser tóxico, ser altamente permeable, de fácil penetración en la membrana celular y ayudar a disminuir los efectos nocivos de la congelación (Marín, 2003). Las temperaturas de almacenamiento deben encontrarse por debajo de los – 20 ºC. Las células deben ser centrifugadas y posteriormente se les debe adicionar glicerol al 20 ó 30 % (v/v) o Dimetil Sulfóxido (DMSO) al 5% (v/v), para ser dispensadas en viales y refrigeradas; mediante este método la viabilidad de los microorganismos puede mantenerse de 3 a 5 años (Marín, 2003). Otro método de conservación a largo plazo es la liofilización, que implica la remoción de agua de las células por sublimación bajo presión. Es uno de los métodos más efectivos de preservación y necesita criopreservantes como leche en polvo al 20 % (p/v) o sacarosa al 12% (p/v). Las cepas pueden ser preservadas por un período de 10 a 20 años. Este método es muy usado cuando se necesita transportar constantemente los microorganismos (Chang et al, 1986) 2.1.5. Pruebas de capacidad probiótica in vitro Cuando el microorganismo ya se ha obtenido, aislado y se han realizado procesos de conservación y de caracterización que permitan su identificación, es necesario realizar pruebas de capacidad probiótica in vitro, que permitan evaluar la resistencia del microorganismo a condiciones características del tracto gastrointestinal, ya que los probióticos deben ser tolerantes a la presencia de ácido y de bilis (Valerio, 2006). Estas pruebas pueden ser cualitativas o cuantitativas (Kumura et al, 2004) y permitirán correlacionar la actividad de las cepas con su capacidad probiótica in vivo. 21 2.1.5.1. Tolerancia a sales biliares La bilis, es una solución acuosa verde – amarillenta cuyos mayores componentes son ácidos biliares, colesterol, fosfolípidos y el pigmento biliverdina. Es sintetizada en los hepatocitos pericentrales del hígado, almacenada y concentrada en la vésicula biliar y liberada al duodeno después de la ingesta de comida. La bilis es un detergente biológico que emulsiona y solubiliza los ácidos grasos de cadena larga y los mono y diacilglicéridos resultantes de la acción de las lipasas intestinales, formando micelas y por lo tanto juega un rol esencial en la digestión de grasas. Esta propiedad de detergente, también le confiere actividad antimicrobiana, al generar la disolución de las membranas bacterianas (Beagley, 2006). Los ácidos biliares primarios, el ácido cólico y quenodeoxicólico, son sintetizados del colesterol de novo en el hígado. Los ácidos biliares son eficientemente conservados bajo condiciones normales, por un proceso de recirculación enteropática. Microorganismos como Lactobacillus spp, poseen enzimas hidrolasas (BSH) que generan la deconjugación de las sales biliares, que hidrolizan el enlace amida y liberan glicina y taurina de la base esteroide, mecanismo por el cual pueden resistir concentraciones de sales biliares en pruebas in vitro y el paso por el TGI en animales (Moser, 2001). Para evaluar la tolerancia a esta sales, se suplementa el medio de cultivo, según el tipo de microorganismo a evaluar, con diferentes concentraciones de sales biliares, generalmente desde 0.1% a 3.5% (p/v) y se incuba por períodos de 24 horas, realizando posteriormente determinaciones de viabilidad mediante turbidez o recuento (Gillilland et al, 1990). 2.1.5.2. Tolerancia a pH En cuanto al pH, microorganismos como Saccharomyces cerevisiae bajo condiciones aeróbicas, pueden usar ácidos orgánicos de cadena corta como ácido acético y láctico como fuentes de carbono, mediante el uso de vías anabólicas, enzimas y mecanismos específicos de transporte (Casal et al., 1996). Una vez en el interior de la célula, estos compuestos generan una acidificación de citoplasma. En general, las células eucarióticas mantienen el pH intracelular a pesar de las variaciones de pH extracelular. Para mantener los valores adecuados dentro de un rango óptimo del metabolismo, las células expulsan protones a expensas de ATP, lo que resulta en la disminución del rendimiento de consumo con respecto a la glucosa, generándose un aumento del pH intracelular, permitiéndole a la levadura resistir rangos amplios (2.4 a 8.6) de pH ácidos (Rose, 1987). La prueba de resistencia a pH, 22 se realiza virando hacia la acidez el medio mediante el uso de HCl, para obtener valores de 1.0 a 5.0, incubando por períodos de 24 horas y realizando la determinación de la viabilidad del microorganismo (Thomas et al 2002). 2.1.5.3. Resistencia a Jugos gástricos Los jugos gástricos son producidos en el estómago, son sustancias muy ácidas cuya función es la de degradar el alimento. Los jugos gástricos están compuestos por agua (98%), sales, ácido clorhídrico, mucoproteínas, enzimas proteolíticas, factor intrínseco o glucoproteína, secreciones endocrinas e inmunoglobulinas. Dentro de estas sustancias se destaca el HCl (ácido clorhídrico), secretado por las células gástricas parietales, que mantiene el pH necesario, ablanda la fibrina y el colágeno, controla el paso de bacterias al intestino y estimula la secreción de secretina, estimulador a su vez de la secreción pancreática y biliar. Contiene enzimas como la pepsina que en medio ácido inicia la digestión de las proteínas, la renina que sirve para cuajar la leche, y la lipasa que inicia la digestión de grasas. Estas son características comunes para animales monogástricos. (Lesson et al, 1990). La evaluación in vitro se realiza elaborando un jugo gástrico artificial, incubando por 24 horas y realizando determinación de la viabilidad del microorganismo (Spencer et al, 2001). 2.1.5.4. Adhesión celular Los cultivos de tejidos celulares se han desarrollado desde los últimos años del siglo XX, con el objetivo de reproducir las condiciones in vivo que permitan el crecimiento de las células, manteniendo al máximo sus propiedades fisiológicas, bioquímicas y genéticas. Los cultivos celulares permiten evaluar diferentes tópicos o mecanismos celulares, tales como actividad intracelular, interacciones celulares (Fresney, 1990) y la adherencia celular en la selección de nuevas cepas con potencial probiótico. La importancia de la adhesión a células epiteliales del intestino para el establecimiento de microorganismos probióticos en el ecosistema del tracto gastrointestinal ha sido demostrada en estudios clínicos (Alander et al, 1997). La adhesión a la mucosa intestinal es importante para la modulación de la respuesta inmune del huésped y la exclusión de patógenos tales como Helycobacter pilori, Salmonella sp, Listeria sp o Clostridium sp. (Alander et al, 1999). La habilidad de adherencia al tracto gastrointestinal de niños y adultos, ha sido estudiada en microorganismos como Lactobacillus rhamnosus (Goldin et al, 1992, Millar et al, 1993). Además in vitro, se han realizado estudios de la inhibición de la adherencia de patógenos 23 como E. coli, Salmonella typhimurium y Enterococcus faecalis, por bacterias ácido lácticas (Todokiri et al, 2001). McFarland et al (1994) reportó la disminución en la incidencia de diarrea causada por Clostridium sp. al administrar levaduras (Saccharomyces cerevisiae var. boulardii;, sin embargo para este año no se había dilucidado si el mecanismo de adhesión y por lo tanto exclusión era usado por el probiótico para evitar la colonización del patógeno. Sin embargo, Gedek (1999) reportó que cepas de E.coli enteropatogénica y de Salmonella typhimurium y Salmonella enteritidis se unían a S. cerevisiae por receptores tipo lectina dependiente de manosas o derivados de estas presentes en la pared celular de la levadura, resultados comparables con los hallados por Pérez et al (2005) en que se observó la adherencia de cepas de Salmonella sp. a levaduras, verificando que este es un mecanismo usado por algunos microorganismos probióticos para evitar la colonización de la mucosa del intestino por parte de patógenos. 2.1.5.4.1. Cultivos celulares Para poder realizar cultivos celulares es importante tener en cuenta la curva de crecimiento celular en la que se evidencia la adaptación a cultivos, a condiciones ambientales y a la viabilidad física del substrato o del suplemento necesario para poder generarse nuevas células. Las fases de crecimiento celular que se presentan en los cultivos son: - Fase de latencia (lag): en esta fase no hay división celular, su duración depende del tiempo y la densidad celular. - Fase logarítmica (log): esta es una fase de proliferación activa, el número de células se eleva exponencialmente. El porcentaje de viabilidad de las células puede llegar a se de un 90 a 100%. - Fase estacionaria: se da división, pero mucha más lenta. El porcentaje de división tiende a balancearse con la tasa de muerte y la viabilidad baja a un 0 a 10%. - Fase declinativa: es un periodo de caída reflejado en el número de células compensadas por la muerte celular (Fresney, 1990). Dentro de los medios de cultivo mas utilizados para la realización de cultivos celulares, se encuentra el Medio Mínimo Esencial de Eagle (MEM), que provee nutrientes básicos a las células, ya sea que se en encuentren en solución o ligados a proteínas, favoreciendo o 24 estimulando el crecimiento celular, los factores de adhesión y propagación en los cuales se proporciona el transporte de hormonas, vitaminas, minerales y lípidos (Sambuy et al., 2005). Se emplea de manera generalizada una adición al 10% (v/v) de suero fetal bovino inactivado (SFB), con el fin de destruir inmunoglobulinas que puedan llevar a la lisis celular; este SFB es una mezcla de biomoléculas pequeñas que promueven el crecimiento celular al contener glucosa, nitrógeno, albúmina, fibronectina, ácido búrico, creatina, hemoglobina, entre otros (Fresney, 1990). Adicionalmente, el medio debe ser suplementado con L-Glutamina, que es un aminoácido esencial para las células en cultivo, debido a que actúa como la principal fuente de carbono, al ser precursor para posteriores biosíntesis y como fuente de energía al participar en el ciclo de Krebs (Mathews, 2003). Además se debe suplementar el medio con antibióticos como Penicilina y Estreptomicina, que eviten contaminación microbiana, pero que no inhiban el crecimiento celular, ni se vean envueltos en el metabolismo de las células (Fresney, 1990). 2.1.5.4.2. Líneas celulares Existen diversas líneas celulares con las que se puede trabajar según el tipo de estudio que se quiera realizar. Para estudios de adherencia celular de probióticos, una de las líneas mas usadas en probióticos es la Caco-2 aisladas por J. Fogh a partir de un tumor primario de colón de humano, en medio mínimo esencial de Eagle en presencia de 5 – 10% de CO2 y un pH de 7.4 (ATCC, 1992) ya que, al cultivarse por un proceso de diferenciación espontánea se permite la formación de la monocapa como se observa en la Figura 1, expresando varias características morfológicas y funcionales típicas de los entericitos maduros (Sambuy et al, 2005), como la similitud en la organización de microvellos en el lado apical de estas células, blanco donde actúan estos microorganismos (Zweibaum et al, 1991), uniones tight entre células adyacentes y actividad de enzimas hidrolasas presentes en el intestino delgado. Las células en edad temprana muestran un efecto citopático, tornándose estas más redondeadas y en forma de racimos (Willicocks, 1990). En la membrana de estas células se encuentran receptores de membrana polarizadas para factores de crecimiento y actividades de transporte de azúcares, vitaminas, ácidos biliares, micronutrientes como metales pesados y nucleósidos (Sambuy et al, 2005) 25 Figura Nº 1. Células Caco – 2 en monocapa con coloración de Wrigth. Fuente (Autores, 2007) 2.1.5.4.3. Ensayos de adhesión Estos ensayos se llevan a cabo para verificar la exclusión competitiva de patógenos y bacterias indeseables, exponiendo cultivos celulares a los microorganismos y verificando la adhesión por microscopía usando la coloración de Giemsa, ya que permite observar a los microorganismos (probióticos) frente a las células como se observa en la Figura 2, al ser una coloración diferencial que evidencia la adherencia y hace posible realizar un conteo expresado en número de microorganismos adheridos a 100 células (Kumura et al, 2004). Figura 2. Análisis de adherencia por microscopio invertido de Kluyveromyces lactis a células Caco-2 diferenciadas con coloración de Giemsa. Fuente (Kumura et al, 2004) 26 2.1.5.5. Reducción del colesterol en presencia de sales biliares Se pueden realizar pruebas de reducción de colesterol, ya que microorganismos como Lactobacillus sp, llevan a cabo una conversión metabólica del colesterol a una forma más asimilable y menos dañina. Esta es una propiedad importante, debido a que en condiciones de estrés se puede afectar el metabolismo de lípidos, afectando la calidad del pollo y sus derivados (Endo, 1999). Estudios relevantes de la asimilación de colesterol por levaduras en condiciones típicamente encontradas en el tracto gastrointestinal no han sido aún reportados; sin embargo, varios estudios han indicado que durante su crecimiento pueden remover colesterol de medios suplementados con micelios de colesterol (Taylor and Parks, 1981). El método de extracción que se utiliza es el propuesto por Fleutoris et al (1998) para la determinación de colesterol en leches y sus derivados con algunas modificaciones propuestas por Spencer (2001). Este protocolo se utiliza ya que la precisión y correlación de los datos, han demostrado que es adecuado, debido a que los resultados obtenidos demuestran un 98.6% de recuperación del colesterol de las muestras analizadas, adicional a la pequeña cantidad de analito que se necesita y al corto tiempo de determinación comparado con otros métodos (Fleutoris et al, 1998). La primera parte de la determinación consiste en la saponificación directa, en la cual el colesterol esterificado debe ser liberado del complejo de lipoproteínas e hidrolizado. La adición de solventes, es importante, ya que el colesterol en la solución de saponificación no se emulsiona, por lo que la adición de estos compuestos, libera la muestra a analizar, evitando resultados erróneos. Inicialmente, se debe aplicar una solución de KOH metanólica o etanol al 95% seguido de KOH al 50%, seguido de calor (60 ºC a 80 ºC) de 10 a 15 minutos. Una vez se ha llevado a cabo esta parte del protocolo, sigue la extracción, en la que se adicionan solventes polares como el hexano que ha mostrado ser excelente, debido a que es menos tóxico que otros compuestos y no forma emulsiones ni peróxidos que podrían interactuar con el colesterol , alterando los resultados (Fenton, 1992). La adición de agua en este proceso es importante, debido a que en presencia de este compuesto se puede hacer una extracción cuantitativa, eliminando la necesidad de una segunda extracción, al producir un aumento de la polaridad de la fase donde se encuentra el KOH metanólico, por lo que el colesterol es forzado a transferirse a la fase menos polar, donde se encuentra el hexano 27 (Anexo Nº 6.6). Posteriormente, se analiza la muestra por cromatografía de gas (Fleutoris et al, 1998) o por la reacción de Liebermann – Burchard (Skoog, 1988). Dentro de la determinación de la reducción de colesterol es recomendable realizar sonicación que es un mecanismo de ruptura celular que genera la lisis de la membrana o pared por la intensa agitación producida al transmitir corrientes eléctricas a un sistema mecánico que genera vibraciones de alta energía produciendo ondas de ultrasonido que forman burbujas microscópicas que se expanden y colapsan contra las células causando la ruptura de la membrana, fenómeno conocido como cavitación (Ruiz et al, 1999); con el fin de determinar si el microorganismo probiótico asimila el colesterol a su estructura confiriéndole mayor resistencia a condiciones adversas que puedan llegar a generar la lisis de la célula, comparado con aquellos que no lo incorporan a su membrana (Psomas et al, 2005). 2.1.6. Resistencia del probiótico a condiciones in vivo Después de la administración del probiótico, los microorganismos no deben morir debido a los mecanismos de defensa del huésped (Netherwood, 1999). Dependiendo de los sitios de administración del probiótico, deben resistir condiciones específicas: significa, por ejemplo que aquellos que son suministrados por vía oral deben resistir enzimas de la cavidad bucal e intestinal como amilasas, lisosimas o lipasas, así como pH bajo debido a la alta concentración de acido clorhídrico en el estómago, a la bilis, jugo pancreático y la mucosidad del intestino delgado (García, 1995). Para realizar la selección de una cepa apropiada para ser usada como probióticos se deben tener en cuenta los ensayos realizados previamente con el microorganismo in vitro, como los mencionados anteriormente, para poder correlacionar los resultados con su capacidad probiótica in vivo. (Fuller, 1992). 2.1.7. Procesamiento de probióticos en productos terminados Al determinar que un microorganismo tiene capacidad probiótica in vitro e in vivo, es importante estudiar la viabilidad y resistencia de los microorganismos durante el procesamiento al que serán sometidos, para su comercialización. El propósito para el cual los probióticos van a ser utilizados determina el método de administración. Para cada método de administración se debe considerar el procesamiento del producto así como las propiedades de los microorganismos probióticos. Durante el proceso de selección se deben determinar las características específicas de cada cepa de microorganismo. 28 Uno de los primeros pasos para la producción de probióticos es el cultivo, que debe realizarse en medios que sean específicos para el tipo de microorganismo a trabajar y que deben favorecer su crecimiento, teniendo en cuenta la concentración que se desea obtener para posteriormente realizar un lavado y secado de los mismos. Muchas de la bacterias ácido lácticas comúnmente usadas como probióticos pueden ser cultivadas a gran escala en fermentadores en condiciones aerobias y luego ser sometidas a un proceso de centrifugación. Los probióticos para animales pueden ser incluidos en el pienso o dosificados directamente en forma de pasta, polvos, gránulos o capsulas (Peña, 2000). Se han desarrollado varios procesos industriales para mantener los probióticos en un estado estable y viable (Fuller, 1992), tales como la microencapsulación o inmovilización (Ferreira et al, 2004), que consiste en introducir las células en una matriz o sistema de pared, con el objetivo de protegerlos de la luz o el oxígeno, que pueden afectarlos negativamente disminuyendo su viabilidad. Al estar encapsulado, las células se van liberando gradualmente, obteniendo mejores resultados al extender su vida útil y facilitando su uso (Chang, 2000). Dentro de las técnicas se encuentran aquellas que son físicas como la extrusión o emulsión en materiales de soporte como alginato de sodio, aceite de girasol (Ré, 1996). El método de la extrusión es el más común y consiste en la formación de cápsulas con hidrocoloides. El polisacárido alginato de sodio ha sido una de las herramientas de microencapsulación mas usadas (Park et al., 2000). Este forma un gel al entrar en contacto con calcio y cationes multivalentes (Fennema, 2000). Las micropartículas de alginato son estables en condiciones de pH bajo. Cuando los probióticos se incluyen en algunos alimentos, el alginato de calcio ofrece una buena protección para microorganismos como Lactobacillus bulgaricus durante la refrigeración en helados, para Bifidobacteria que inmovilizado en este compuesto es más resistente a valores de pH ácidos comparado con células libres en alimentos como mayonesa (Khalil et al., 1998). Una técnica comúnmente usada en la industria de alimentos es la compresión en pellets del probiótico, pero esto hace que durante este proceso la viabilidad baje dramáticamente. Otro aspecto a tener en cuenta son los aditivos que son incorporados al alimento y que puedan afectar a los microorganismos probióticos, sustancias que se incluyen al alimento para evitar contaminación o suplementos como sulfato de hierro, óxido de hierro, sulfato de cobalto, que deben ser adicionadas con cuidado teniendo en cuenta las cantidades que pueden ser perjudiciales para los probióticos (Fuller, 1992). 29 2.2 PROBIÓTICOS PARA AVES DE CORRAL 2.2.1. Aves de corral y el tracto gastrointestinal La fisiología del sistema digestivo de las aves es similar en términos generales al de los cerdos y los humanos. Todos ellos, animales monogástricos con poca capacidad para digerir la celulosa y otros carbohidratos complejos en comparación con los animales herbívoros rumiantes. El aparato digestivo de las aves es más ligero, más corto y el alimento lo recorre con mucha mayor rapidez que en especies no rumiantes (Rose, 1997). - En la boca presenta un pico córneo característico que suele ser cortante y puntiagudo, dispone de una lengua no flexible que solamente se mueve hacia atrás y pasa las partículas del alimento rápidamente hacia la faringe. La saliva escasa, produce poca descomposición enzimática del alimento en la boca. - El buche es un saco dilatable que funciona almacenando alimentos. Dispone de una población de bacterias residentes, principalmente Lactobacillus, que fermentan parte de los carbohidratos a ácido láctico, por lo que desciende el pH del alimento durante su retención en el buche. - El proventrículo o verdadero estómago, se distingue por su gruesa membrana mucosa glandular. Las glándulas segregan ácido clorhídrico y pepsinógeno. La acidez del alimento es reducida hasta un pH que permite la formación de pepsina que cataliza la hidrólisis de las proteínas. - En la molleja se realiza la mayor parte de la hidrólisis de la proteína catalizada por la pepsina. Debido a su forma de esfera aplastada rodeada de músculos potentes, se provoca una presión intensa en su interior. Adicional, ya que se retienen partículas de arena, se genera una superficie abrasiva, que ayuda en la molturación de los alimentos. - En el intestino delgado las partículas penetran al duodeno donde se vierte la bilis y las enzimas pancreáticas. El antiperistaltismo resulta particularmente importante para que sea máxima la hidrólisis y la absorción de lípidos. La acidez de la molleja no aporta el pH adecuado para que actúe la lipasa, aunque las sales biliares no son dependientes el pH y continuarán emulsionando los glóbulos de grasa. - El colón y el recto son muy cortos. Los ciegos son los órganos en que se produce la fermentación bacteriana del material alimenticio no digerido en la mayoría de los animales rumiantes. 30 - En la cloaca los residuos alimenticios no digeridos se acumulan en esta parte desde son excretados. La orina se acumula también en la cloaca y se mezclará con las heces antes de ser eliminadas del cuerpo de las aves. 2.2.2. La flora intestinal normal de aves de corral La microflora intestinal de aves de corral es compleja y las interacciones entre los diferentes tipos de organismo son complicadas. Las interacciones que son mejor comprendidas son probablemente las más simples. Sin embargo el conocimiento de la flora intestinal de pollos aumenta continuamente. Los microorganismos predominantes son Lactobacilos que producen principalmente acido láctico y acético de manera que el buche de un pollo saludable presenta valores de pH entre 4 y 5; de esta manera microorganismos menos acidúricos no crecen en la misma cantidad. El pH del proventrículo y la molleja es mas bajo (de 1 a 2) (Fuller, 1992) y la sobrevivencia microbiana depende de la resistencia a ácidos. En el ciego se encuentran anaerobios facultativos que incluyen enterobacterias como E. coli, Citrobacter sp., Salmonella sp., Proteus sp. y Klebsiella sp. y otros microorganismos en menor proporción como Pseudomonas sp. y levaduras. En polluelos el tracto alimentario es estéril por lo que es rápidamente colonizado por anaerobios facultativos principalmente Coliformes y Estreptococos, incluso algunos Clostridios pueden estar presentes (Fuller, 1992). 2.2.3. Infecciones bacterianas tempranas en polluelos Las bacterias amenazan a los pollitos tanto en la incubación natural o en la artificial. Cuanto mayor es el grupo, con más facilidad se diseminan los microorganismos en la población. Estos microorganismos patógenos pueden proceder de la madre o bien, tener su origen en deficientes condiciones del entorno. Determinadas acciones ambientales, como la existencia de corrientes de aire o de temperaturas inadecuadas en los gallineros, favorecen la multiplicación de microorganismos en las aves y aumentan la propensión de estas a sufrir enfermedades. Entre los patógenos, se debe tener en cuenta principalmente a las Salmonelas (Salmonella pullorum, Salmonella enteritidis). Por eliminarse en las heces, ingresan rápidamente por el pico hacia el tracto gastrointestinal de otros pollitos, ya en la 31 incubadora, pero también por vía respiratoria con el aire que contiene partículas de excrementos (Woernle, 1996). 2.2.4 Aplicación y evaluación de probióticos en aves de corral. Los probióticos para pollos son diseñados para reemplazar organismos benéficos que no se encuentran en el tracto alimenticio o para proveer al pollo de los efectos benéficos de los mismos (Abdudrahim, 1999). Estos microorganismos pueden estar ausentes porque los métodos actuales de crianza previenen el contacto entre el polluelo y su progenitor previniendo la transferencia vertical rápida de bacterias benéficas o por prácticas de manejo que pueden alterar la microecologia intestinal; adicionalmente, la producción de corticoides endógenos por estrés, deprimen la respuesta inmune del animal, desencadenando un desequilibro en la flora luminar del tubo digestivo. En el caso de los pollitos que son criados lejos de sus madres y adquieren del medio ambiente su flora, trae como consecuencia que la flora intestinal de los polluelos recién eclosionados “modernos” sea deficiente comparado con la de los pollos incubados por sus madres biológicas (Fuller, 1992). Existen dos grupos grandes de preparaciones de probióticos: aquellas que primariamente son efectivas en el buche (en esta parte del TGI se encuentra gobernado el mecanismo de colonización, ya que allí se lleva a cabo la multiplicación de la flora bacteriana y posteriormente esta migra para colonizar las partes bajas del sistema digestivo) y regiones anteriores del tracto alimenticio y aquellas cuyo efecto es principalmente en el intestino ciego. Sin embargo ambas preparaciones pueden ser efectivas a través de todo el sistema digestivo (Fuller, 1992). Si se proporciona microflora a las aves, por lo común de contenido de ciegos, se produce una alta protección estable 32 horas después, lo cual limita con eficiencia la prevalencia de la infección de Salmonella sp. La exclusión competitiva funciona contra la exposición animal continua elevada y con frecuencia, previene las infecciones cuando existe contaminación con Salmonella sp. en bajas cantidades (Jordan F. y Pattison M, 1998). Aunque un número de observaciones empíricas ha sugerido que, algunas preparaciones que contienen bacterias son probióticos efectivos, el uso de organismos vivos es enfatizado para muchos productos debido a que la colonización intestinal es esencial para la eficacia del mismo (Fuller, 1992). De la misma manera, se ha estudiado la posibilidad de proporcionar a los pollitos un cultivo de heces proveniente de pollos adultos, como 32 alternativa al uso de antibióticos para controlar Salmonella sp, ya que mediante este método el patógeno puede no ser eliminado y generar resistencia (Oliveira et al, 2000). Según Tortuero (1973), en pollos de engorde, la administración de probióticos fue tan benéfica como la de un promotor de crecimiento convencional en lo que hace referencia al incremento de peso y al índice de conversión. Dilworth y Day en 1978 condujeron experimentos evaluando probióticos en la dieta de pollos de engorde. Se obtuvo un significativo mejoramiento en el peso y la eficiencia del alimento. Para 1979, Crawford encontró que un cultivo probiótico en la proporción de 454 g por tonelada de alimento, mejoró la conversión alimenticia en pollos sometidos a la prueba desde el nacimiento hasta el día del sacrificio. En 1978 Couch agregó un probiótico a un alimento para pollos de engorde obteniendo un incremento en los promedios de peso, decreciendo la mortalidad en un 4% (Fuller, 1992). Para asegurar la habilidad de colonizar, el probiótico debe adherirse al epitelio del buche, crecer en el medio ambiente del intestino y resistir mecanismos inhibitorios naturales o innatos. La tolerancia a bajos rangos de pH también es importante para permitir la colonización del intestino delgado y la sobrevivencia al medio ambiente de la molleja. Factores adicionales tales como temperatura óptima de crecimiento y resistencia a ácidos grasos insaturados pueden ser también significantes en la colonización (Fuller, 1992). 2.3. Saccharomyces cerevisiae 2.3.1. Generalidades Levadura es un nombre genérico que agrupa a una variedad de hongos, incluyendo tanto especies patógenas para plantas y animales, como especies no solamente inocuas sino de gran utilidad. Se encuentran clasificadas dentro de los Ascomicetes y Basidiomicetes pertenecientes al orden Endomicetales. Las levaduras constituyen el grupo de microorganismos mas íntimamente asociado al progreso y bienestar de la humanidad. La membrana celular consta de polisacáridos y muy poca quítina. Tienen glucógeno como sustancia de reserva y contienen también numerosas vitaminas (Barnett, 1990). Saccharomyces cerevisiae es un anaerobio facultativo: transformando azúcar a la misma velocidad, la levadura aeróbica produce dióxido de carbono, agua y una producción relativamente alta de nueva levadura, mientras que crecida anaeróbicamente tiene una 33 velocidad relativamente lenta de crecimiento, que se acopla a una alta conversión de azúcar en alcohol y dióxido de carbono. Dentro de sus requerimientos nutricionales se encuentra el carbono, como el mayor compuesto encontrado en la célula, utilizando para su metabolismo glúcidos como hexosas, disacáridos y trisacáridos. El nitrógeno, es utilizado en forma de ión amonio y es utilizado en aminoácidos, vitaminas y nucleótidos. Otros elementos como fósforo, azufre, potasio, magnesio, calcio, zinc y manganeso son importantes dentro de su metabolismo (Bouix, 2000). Algunas especies de levaduras del género Saccharomyces son capaces de llevar a cabo el proceso de fermentación, propiedad que se ha explotado desde hace muchos años en la producción de pan y de bebidas alcohólicas. El estudio de las levaduras como modelo biológico ha contribuido de manera muy importante a dilucidar los procesos básicos de la fisiología celular. (González, 2000). En años recientes se ha implementado su uso como probiótico, entre las que se incluyen Saccharomyces cerevisiae, Saccharomyces uvarum y Saccharomyces cerevisiae var. boulardii (Arora, 1991). Dentro del género Saccharomyces, la especie cerevisiae constituye la levadura y el microorganismo eucariote más estudiado. Así, desde fines del siglo XIX se reconocieron algunas de las ventajas que ofrece el uso de esta levadura como biológico en la investigación: por un lado, la facilidad con la que se obtienen grandes cantidades de este microorganismo debido a su facilidad de cultivo y su velocidad de división celular (aproximadamente dos horas); y por otro, el hecho de que S. cerevisiae posee un ciclo de vida que al incluir una fase sexual, permite abordar estudios con las herramientas que provee la genética formal (González, 2000). Por medio de receptores anclados a la membrana celular, esta levadura puede responder a la presencia de moléculas que actúan como señales en una condición fisiológica determinada, estas señales se traducen por un sistema de señalización en el que participan cinasas denominadas “proteín-cinasas”, que le permiten resistir exposiciones a alta o baja osmolaridad, la presencia de una alta o baja concentración de sal en el medio de cultivo o de pH, por lo que se genera una señal, que al interaccionar con un receptor específico, pasa a través de una serie de moléculas intermediarias hasta modificar un determinado regulador transcripcional, que es capaz de activar o reprimir la expresión de un gen o grupos de genes. Esto le permite a la célula transcribir de manera selectiva un grupo de genes específicos, cuya traducción resultará en la síntesis de los productos que le permitan contender de manera eficiente con la condición fisiológica en la que se encuentra, y por tanto mantener la 34 viabilidad, esporular, crecer en forma de filamentos, etc. Así se han descrito en S. cerevisiae distintas cascadas que responden a estímulos ambientales diferentes y específicos (González, 2000) 2.3.2. Ciclo asexual y sexual de Saccharomyces cerevisiae S. cerevisiae se divide por gemación y puede tener una reproducción asexual cuando se encuentra en su forma haploide. Durante la fase vegetativa, la levadura se divide por gemación como se observa en la Figura 2. En la parte asexual, la célula hija inicia su crecimiento formando una yema en la célula madre, posteriormente ocurre la división nuclear, la síntesis de la pared y finalmente la separación de las dos células. Durante muchos años se consideró que a diferencia de los hongos denominados filamentosos, S. cerevisiae era incapaz de formar hifas o filamentos y que su crecimiento vegetativo, a través de la gemación, únicamente resultaba en la formación de células esféricas denominadas levaduras. Sin embargo, en estudios recientes se encontró que Saccharomyces era capaz de formar hifas (pseudohifas), denominadas así debido a que por la fisión binaria se generan cilindros que terminarán en elongaciones y la formación de un septo, por donde se dividirá la célula (González, 2000). Figura Nº 3. Microscopia electrónica de formación de yemas de Saccharomyces cerevisiae. Fuente (www.bath.ac.uk/bio-sci/wheals2.htm) En la parte del ciclo sexual, a partir de un cigoto se forma un asca que contiene cuatro ascosporas haploides, generadas cuando la célula diploide se divide por meiosis. S. cerevisiae posee dos tipo sexuales: a y α, determinados por un par de alelos heterocigos: MATa y MATα, que al combinarse, dan origen a las nuevas células diploides (González, 2000) que se observan en la Figura 4. 35 Figura Nº 4. Reproducción sexual y asexual de Saccharomyces cerevisiae. Fuente (Tuité, 2001) 2.3.3. Cinética de crecimiento de Saccharomyces cerevisiae Cuando se adicionan levaduras a un medio adecuado que supla los requerimientos nutricionales y metabólicos del microorganismo, se puede observar la cinética del crecimiento, representado en una curva, con diferentes fases como se observa en la Figura 5. Esta curva es de gran utilidad, ya que permite obtener una población de células de número conocido, incubando durante un tiempo determinado (Barnnet, 1990). Figura Nº 5. Curva de crecimiento de levaduras. Fuente((http://www.ugr.es/~eianez/Microbiologia/12crecimiento.htm) 36 - Fase de latencia: al inocular la levadura a medio fresco, el crecimiento no se da inmediatamente, sino después de cierto tiempo. Es una fase de adaptación al medio, ya sea porque deben sintetizar enzimas que les permitan degradar los componentes del mismo, o porque han sufrido algún tipo de injuria o proceso como conservación. - Fase exponencial: la velocidad de crecimiento es lenta inicialmente, pero después se incrementa constantemente. Debido a la rápida reproducción de la células, los nutrientes son consumidos rápidamente y es importante que las condiciones de crecimiento referentes a temperatura y agitación, para intercambio de gases, se encuentre controlada. - Fase estacionaria: no hay incremento neto (o decremento) del número de células, sin embargo todavía ocurren muchas funciones celulares, incluyendo el metabolismo energético y algunos procesos sintéticos. En esta fase, se da un agotamiento de los nutrientes presentes en el medio o por producción de sustancias tóxicas. - Fase de muerte: el número de células disminuye, por la ausencia total de nutrientes, puede incluso generarse lisis celular, la disminución es exponencial, pero la velocidad de muerte no es tan rápida como el crecimiento exponencial. 2.3.4. Antecedentes de Saccharomyces cerevisiae como probiótico Al encontrarse activa o viable con un conteo de 10 mil a 20 mil millones de células vivas por gramo, esta levadura se utiliza principalmente como probiótico, algunas de sus funciones son: - Promotor de crecimiento - Aumenta la producción de vitamina B. - Mayor ganancia de peso. - Rápida digestión de algunos alimentos. - Acción estimulante de la inmunidad. - Mejora la asimilación de nutrientes. - Corrige el balance de la población microbiana (Agarwal et al, 2000). Dentro de las levaduras, la especie S. cerevisiae var. boulardii ha sido la más ampliamente estudiada en ensayos clínicos, ya que ha mostrado estar asociada a la prevención de diarrea, aunque el mecanismo aun no se encuentra bien definido. Este microorganismo fue aislado inicialmente de frutas que eran usadas en la medicina popular. En Francia, en 1950 37 se introdujo un producto que contenía la levadura y en la actualidad es comercializada de manera liofilizada en Europa, Africa y Suramérica. No es encontrada como parte de la flora habitual del tracto gastrointestinal y debe ser administrada continuamente, para asegurar su permanencia y colonización de enterocitos (Toma et al, 2005). Esta levadura puede secretar una proteasa que digiere dos exotoxinas, toxina A y toxina B, que están implicadas en la diarrea y la colitis causada Clostridium difficile. (Surawicz et al, 1989). Basados en estudios experimentales se han propuesto 3 posibles mecanismos de acción por el que esta levadura puede controlar la diarrea i) actividad antagónica in vitro contra varias bacterias patógenas (Brugier and Patte, 1975) y especies de Candida (Ducluzeau and Bensaada, 1982); ii) estimulación de una secreción de inmunoglobulinas intestinales, defensa ante infecciones (Machado-Caetano et al, 1986; Buts et al, 1990); iii) reducción de la producción o acción de varias toxinas bacterianas (Massot et al, 1982); y iv) aumento de la actividad de disacáridos en la mucosa intestinal en ratas y humanos (Buts et al, 1986). Según Rodrigues et al (1996) la actividad antagónica de la levadura se observó in vivo contra Candida sp., pero no contra Salmonella typhimurium, Shigella flexneri y Escherichia coli. Otro mecanismo posiblemente envuelve el sistema inmune como responsable de la protección conferida por S. cerevisiae var. boulardii contra estos enteropatógenos. Toma et al (2005), encontró que probablemente S. boulardii puede estar implicada en la conversión de diferentes compuestos tóxicos tales como agentes mutágenicos en productos no reactivos, característica que junto la resistencia a temperatura y acidez (Fietto et al, 2004) y la resistencia a antibióticos, genera un panorama muy prometedor para el uso de este microorganismo comparado con otros probióticos. S. cerevisiae es un microorganismo probiótico que ha sido estudiado en especies como cerdos, bovinos y caballos en los que se ha utilizado como promotor del crecimiento (Medina et al, 2002). En el estudio de Dann et al (2003), se observó que se mejoraba la producción de leche cuando se suministraba la levadura en la dieta de bovinos, En animales como terneros se ha mostrado que se mejora la absorción de nutrientes y se mejora la producción de los animales a los que se suministra en la dieta (Kung et al, 1997). Saccharomyces contiene grandes cantidades de ß-glucanos en su pared celular, que actúan como promotores de la activación inespecífica del sistema inmune. Estos compuestos, son polímeros de glucosa con uniones beta-(1-3) que se pueden encontrar en forma de 38 partículas o en forma soluble y tienen capacidad inmunoestimulante, mediante la estimulación de macrófagos y neutrófilos y de esta manera protegiendo al huésped de infecciones, ya que en situaciones de estrés se producen y liberan corticoides endógenos, que deprimen la respuesta inmune y se genera un desequilibrio en la flora intestinal, situación propicia para la colonización por patógenos (Pelizon et al, 2003). El valor nutritivo de la levadura varía dependiendo del sustrato utilizado para su crecimiento y también del proceso industrial al cual es sometida (Alvarez y Valdivia, 1980). Existen reportes que indican que cantidades del 10 al 15 % de levadura, como sustituto parcial de la harina de soya en raciones de pollos de engorde, no afecta el comportamiento productivo de las aves (Murakami et al., 1993). Las levaduras no son parte de la flora normal de los monogástricos, sin embargo al estar situadas en bajo ciertas condiciones anaerobias pueden aportar enzimas como lipasas, proteasas, vitamina B, aminoácidos, minerales, iones metálicos y otros cofactores (Pardio, 1996). Al consumir el oxígeno debido a su metabolismo, pueden estimular el crecimiento de microorganismos anaerobios característicos del tracto gastrointestinal. Al encontrarse adheridas a enterocitos, y debido la absorción celular que tiene en su pared, esta levadura puede ser fuente y reserva de zinc, potasio y cobre. Pueden aportar factores de crecimiento, vitaminas del grupo B, inhiben toxinas y debido a la producción de ácido glutámico aumentan el gusto de los alimentos (Dawson, 1987). En Cuba se han realizado estudios que incluyen S. cerevisiae, con bagazo de caña de azúcar en concentraciones de 10%, 15% y 20 % (p/p) a la dieta tradicional suministrada a los polluelos para evidenciar los beneficios que genera en el animal, encontrando que después del período de evaluación, la levadura puede constituir portadores de nutrientes con características favorables, aumentando el peso del pollo en un período de tiempo aceptable y a la vez disminuyendo los costos de producción (Solano et al, 2005). Los beneficios que ofrece este microorganismo para los pollos radica en lo enumerado anteriormente y además porque se señala que su utilización como probiótico, reduce algunos enteropatógenos, produce cambios favorables en la mucosa intestinal y mejora el comportamiento productivo con raciones bajas en proteína (Kumprechtová et al., 2000) por lo que la evaluación de este microorganismo se hace muy importante como mecanismo para verificar que las cepas en estudio pueden ser suministradas al polluelo, y resisten las condiciones del tracto gastrointestinal. 39 2.4. INDUSTRIA AVÍCOLA EN COLOMBIA La crianza de aves útiles al hombre se origina en la antigüedad, por lo que la avicultura moderna no es más que la modernización de los métodos, ya que las aves son animales cuyas necesidades nutricionales y técnicas han alcanzado un alto grado de demanda, lo que ha generado un mayor perfeccionamiento, generando mejores niveles de calidad, pero asimismo aumentando los costos de mantenimiento En Colombia, los años 80’s generaron que el desarrollo tecnológico en esta área se disparara y ya para el año 1991, la avicultura generaba un 2 % del PIB total del país (Molina, 2002). En la actualidad, la industria avícola ha aumentado notablemente la producción al registrarse un crecimiento de 10.6% para el 2006, el doble del alcanzado en el 2005 (5.4%), con lo cual este sector sigue consolidándose como el más dinámico de la economía nacional. En el 2006 se presentó un consumo anual de 849.557 toneladas de pollo, siendo el consumo per cápita de 19.9 Kg/año (Figura 6), aumentando notoriamente sobre el consumo per cápita de carne bovina o porcícola (FENAVI, 2006). CONSUMO PER CÁPITA DE POLLO EN COLOMBIA (1970 -2006) (kilogramos / año) 25,0 19,8 20,0 18,1 15,6 15,2 14,8 13,8 13,3 12,9 12,0 12,0 11,7 11,2 10,6 9,7 15,0 10,0 7,7 8,1 7,6 7,9 7,8 8,6 6,9 4,7 5,0 2,8 2,9 2,4 2,6 1,9 2,2 1,6 1,8 1,3 1,2 1,0 5,2 5,3 5,8 3,6 2006 2005 2004 2003 2002 2001 2000 1999 1998 1997 1996 1995 1994 1993 1992 1991 1990 1989 1988 1987 1986 1985 1984 1983 1982 1981 1980 1979 1978 1977 1976 1975 1974 1973 1972 1971 1970 0,0 Figura Nº 6. Consumo per cápita de pollo en Colombia (1970 – 2006). Fuente (FENAVI, 2006) La oferta total de pollo en canal registró en el primer semestre del año un crecimiento de 8%: de una producción en igual periodo del 2005 de 367.709 toneladas, se pasó a 397.197 40 (Figura 7). Esto quiere decir que desde el 2000 se ha mantenido un crecimiento de 5.1% promedio anual (FENAVI, 2006). La incidencia económica de la industria avícola en Colombia es importante, ya que es uno de los sectores con mayor crecimiento, la exportación para 2006 en pollitos de 1 día generó ingresos de U$ 105’110.000 y con una tendencia al aumento de la demanda, condiciones que crean el escenario perfecto, para que el uso de probióticos disminuya las pérdidas de polluelos, reduciendo los costos de producción, ya que para el año 2006 estos costos presentaron un aumento del 8.7%; beneficiando en primer lugar a la empresa y es segundo al consumidor, que deberá pagar un valor menor por un producto de calidad. Figura Nº 7. Producción de pollo (toneladas) en 2001 – 2006. Fuente (FENAVI, 2006) 41 3. FORMULACIÓN DEL PROBLEMA Y JUSTIFICACION 3.1. Formulación del problema Las levaduras utilizadas como probióticos ofrecen beneficios tales como la capacidad de resistir el pasaje a través del tracto gastrointestinal sin que presenten pérdidas significativas de viabilidad debido a que, activan mecanismos para tolerar condiciones de estrés como la exposición a sales biliares, jugos gástricos, ácidos orgánicos y enzimas digestivas, así como la competencia con microbiota propia del huésped (Holzapfel et al, 1998). Adicionalmente, se han reconocido ventajas de su uso ya que en especies como bovinos, aves, cunícola y porcícola ayuda a la promoción del crecimiento al mejorar la digestión del animal, al aumentar la absorción de nutrientes, producir cambios favorables en la mucosa intestinal, así como disminuir o reducir notablemente la presencia de microorganismos patógenos (Perdomo et al, 2004) como Salmonella sp. (Mantilla y Suá 2000), Pseudomonas sp, S. aureus, Clostridium perfringes y Clostridium botulinum (Woernle, 1996); generar efectos anticarcinogénicos, anticolesterémicos e inmunoestimulatorios (Oyetayo et al, 2005), así como el crecimiento a una temperatura relativamente alta (37 ºC) y la resistencia a varios antibióticos (Buts, 2005). A pesar de todas las ventajas que ofrece el uso de levaduras como probióticos, los microorganismos más estudiados y utilizados para tal fin pertenecen a los géneros Lactobacillus s.p, Bifidobacterium y en menor proporción levaduras debido a que estas no hacen parte de la flora intestinal de animales monogástricos (Otero, 1997). Aunque se han realizado estudios preliminares de capacidad probiótica especialmente de Saccharomyces boulardii como los mencionados anteriormente, aún existen interrogantes sobre los mecanismos de tolerancia de este microorganismo a condiciones características del tracto gastrointestinal, que le permitirán sobrevivir para llegar a las células blanco y afectar benéficamente al huésped. Debido a lo anterior, en este proyecto se pretende evaluar la capacidad probiótica in vitro de una cepa nativa de Saccharomyces cerevisiae como futuro suplemento probiotico en la dieta de polluelos u otras especies de uso zootécnico, con el fin de comprobar que este microorganismo cumple con los requerimientos necesarios para su administracion por via oral para animales monogástricos. 42 3.2. Justificación En Colombia, se han producido grandes cambios tecnológicos en la industria avícola, los cuales se han ido materializando en aves genéticamente más resistentes. Nuevas formas de manejo y de cría han llevado a obtener aves precoces desarrolladas en incubadoras con ambientes controlados y limpios. Estas aves, son criadas en ambientes más tecnificados, eficientes y aislados que permiten mayor concentración de aves por metro cuadrado. En consecuencia, se están criando aves cada vez más susceptibles a estrés que presentan un establecimiento tardío en la flora intestinal normal, generando como resultado una defensa natural disminuida contra las enfermedades bacterianas de origen intestinal (Barrow, 1992). A pesar de que en países desarrollados existen varias formulaciones con microorganismos probióticos para mejorar e incrementar la tasa de supervivencia en pollos u otros animales monogástricos, en Colombia aún no existe un biopreparado a partir de cepas de levaduras nativas útil en la alimentación animal. En trabajos previos se ha demostrado in vitro la capacidad probiótica de levaduras nativas como una alternativa biotecnológica válida ya que haría la producción a gran escala menos dependiente de productos importados que generan mayor inversión (Aguirre, 2003). En el presente estudio se ha planteado la utilización de una cepa nativa, evaluando la capacidad probiótica in vitro de Saccharomyces cerevisiae, mediante pruebas de tolerancia a sales biliares, tolerancia a rangos de pH, resistencia a jugos gástricos, reducción del colesterol en presencia de sales biliares y ensayos de adhesión celular a células Caco-2; comparada con una cepa de Saccharomyces cerevisiae var. boulardii utilizada actualmente como un probiotico comercial, con el fin de cumplir con la etapa preliminar de formulación de un producto probiotico destinado a ser un suplemento para dietas del sector avícola 43 4. OBJETIVOS • 4.1 Objetivo General Evaluar la capacidad probiótica in vitro de una cepa nativa de Saccharomyces cerevisiae. • 4.2 Objetivos Específicos • Determinar la capacidad probiótica de la cepa estudio (cepa A) y de la cepa control (cepa B) por medio de pruebas de tolerancia a sales biliares en concentraciones de 0.5% a 3.0% (p/v). • Comprobar la capacidad probiótica de la cepa estudio (cepa A) y de la cepa control (cepa B) por medio de pruebas de tolerancia a rangos de pH de 2.0 a 5.0 y resistencia a jugos gástricos. • Determinar la capacidad in vitro de reducción de colesterol en presencia de sales biliares de la cepa estudio en comparación con la cepa control. • Evaluar la adhesión de las levaduras (Cepa A y B) a las células Caco-2. 44 5. MATERIALES Y MÉTODOS 5.1. Métodos 5.1.1. Obtención y conservación de Cepa A y Cepa B La Cepa A (Saccharomyces cerevisiae) fue previamente aislada a partir de cultivos de vid y la Cepa B (Saccharomyces cerevisiae var. boulardii) fue obtenida a partir de un probiótico comercial. Ambas cepas fueron conservadas en un banco de células en medio YPG adicionado de glicerol 30% (v/v) y mantenidas a -70 ºC. Para cada banco se realizó un control de calidad cada dos meses, por medio de la técnica de recuento en placa. 5.1.2. Determinación de capacidad probiótica in vitro Cada una de las pruebas de capacidad probiótica se realizó por triplicado, tanto para la cepa en estudio como para la cepa control. 5.1.2.1. Tolerancia a Sales Biliares • Producción del inóculo En erlenmeyer relación 1/5 se realizó el inóculo de la cepa A y cepa B, incubándose en condiciones de operación de 30ºC, 150 rpm durante 12 horas como mínimo hasta obtener un valor de absorbancia620nm de 0,80. Finalizado el tiempo se verificó la pureza mediante coloración de Gram (Aguirre, 2003). El caldo YPG fue suplementado con Bile Oxgall (Difco®) hasta obtener una concentración final de 0.05%, 0.1%, 0.15%, 0.2%, 0.25% y 0.3% respectivamente. Posteriormente, el mdeio fue esterilizado a 121 ºC durante 20 min (Spencer, 2001). 1.0 mL de la suspensión de las levaduras previamente obtenidas fue inoculado en 9 mL de caldo YPG suplementado con sales biliares. Posteriormente, fueron incubados a 30 ºC durante 24 h. Finalizado el tiempo de incubación, se realizaron recuentos en placa de la siguiente manera: de cada concentración de caldo YPG adicionado de sales biliares se realizaron diluciones 45 seriadas en solución salina al 0.85% (p/v). Las diluciones fueron sembradas por el método de microgota tomando 20 ųl de cada muestra y depositándolos en agar YPG, desde una altura no mayor de 2 cm, dividiendo la caja en cuatro cuadrantes y sembrando en cada uno dos réplicas de las diluciones realizadas. Después de la incubación se escogieron las placas que tenían colonias aisladas en un rango entre 10 – 20 colonias, el número de colonias obtenido fue multiplicado por el factor de corrección (50) para completar el recuento al mililitro y finalmente, por el factor de dilución empleado. El recuento se reportó en UFC/ mL (Collins, 1989). 5.1.2.2. Tolerancia a pH • Producción del inóculo En este procedimiento se utilizó un inóculo preparado de la misma manera, que el descrito en el numeral 5.1.2.1 Al caldo YPG de pH 5.0 se le adicionó HCl sin diluir para disminuir el pH a valores de 2.0, 2.5, 3.0, 3.5, 4.0, 4.5 y 5.0. Posteriormente se esterilizó a 121 ºC durante 15 minutos. 1.0 mL de la suspensión de las levaduras previamente obtenidas fue inoculado en 9 mL de caldo YPG con pH ajustado. Posteriormente, fueron incubados a 30 ºC durante 24 h. Finalizado el periodo de incubación, se realizaron recuentos en placa de la siguiente manera: de cada caldo YPG con pH ajustado, se realizaron diluciones seriadas en solución salina al 0.85% (p/v) y se sembraron por el método de microgota descrito anteriormente. 5.1.2.3. Determinación de resistencia a Jugos Gástricos • Producción del inóculo En este procedimiento se utilizó un inóculo preparado de la misma manera, que el descrito en el numeral 5.1.2.1 Para la elaboración del jugo gástrico se tomó NaCl (0.2g) y pepsina (0.32g), ajustando a pH final de 2-2.3 ± 0.2 con HCl sin diluir, se completó a 100 mL con agua destilada estéril. Como control, se ajustó jugo gástrico artificial a un pH de 6.5-7.0 ±0.2 con NaOH 5N. 46 Posteriormente, se esterilizó por filtración con filtros de membrana de 0.22µm (Spencer, 2001 Se inoculó 1 mL de la suspensión con levaduras previamente preparado, en 9 mL del jugo gástrico artificial pH 2 - 2.3 y pH 6.5 - 7.0 (control). Se incubaron a 30 ºC, tomando muestras a las hora 0, 1, 2, 3, 4 y después de las 24 horas para observar la viabilidad celular (Spencer, 2001). De cada una de las muestras tomadas en cada hora se realizaron diluciones y recuentos en placa por el método de microgota descrito anteriormente. 5.1.2.4. Reducción del colesterol en presencia de sales biliares • Elaboración curva patrón La curva patrón se realizó por duplicado. Se tomaron valores conocidos de colesterol, (60, 70, 80, 90, 100, 110, 120 y 130 μg/mL), y se siguió el protocolo de extracción descrito a continuación para las muestras. Con los datos obtenidos se graficaron los valores de Absorbancia (550 nm) obtenidos contra las concentraciones conocidas y se halló la ecuación de la recta para determinar de los valores de absorbancia obtenidos posteriormente la concentración de colesterol en el medio para la cepa A y B. • Producción del inóculo En este procedimiento se utilizó un inóculo, preparado de la misma manera que el descrito en el numeral 5.1.2.1 Medio para la determinación de reducción de colesterol Medio para la reducción de colesterol (Caldo YPGCHO): Se adicionaron 0.3 g de Bile Oxgall (Difco ®) a 100 mL de caldo YPG y se esterilizó a 121 ºC durante 20 minutos(Spencer, 2001). Posteriormente se adicionaron 224μg/mL de Lipids Colesterol Rich (Sigma ®) ). La solución se mantuvo almacenada a 4 ºC (Psomas et al, 2003) 47 • Determinación de la reducción de colesterol Se inocularon 20 mL del medio anterior con 0.2 mL (1% (v/v)) de un cultivo de levaduras tras 12 horas de incubación, la mezcla se llevó a incubar a 30ºC durante 12 horas. Como control se uso caldo YPGCHO sin inocular. Posteriormente se realizó una centrifugación a 8000g por 15 min. El sobrenadante (muestra) fue colocado en tubo al cual se le adicionaron 3 mL de etanol al 95% (v/v), seguido por 2 mL de hidróxido de potasio al 50% (v/v), agitando en vórtex después de la adición de cada componente. Posteriormente se calentó el tubo en baño maría a 60ºC durante 10 minutos, luego se enfrió hasta los 20ºC, a continuación se adicionaron 5 mL de hexano agitando en vórtex durante 20 segundos. Finalmente,se adicionaron 3 mL de agua destilada y se repitió el mezclado con vórtex. La muestra fue durante 15 minutos o más mantenida a temperatura ambiente hasta que se evidenció la fase de separación. De la fase acuosa (capa de hexano) se transfirieron 2.5 mL a un tubo limpio y se evaporó en un horno a 60ºC. El residuo formado se resuspendió en 4 mL de reactivo 0-phthalaldehido, esta muestra se mantuvo a temperatura ambiente por 10 minutos, luego de los cuales se le adicionaron 2 mL de ácido sulfúrico concentrado agitando vigorosamente. Luego de 10 minutos de reposo se procedió a medir la absorbancia a 550 nm contra el reactivo blanco expresando los resultados en μg de colesterol/mL. Con estos resultados se determinó la cantidad de colesterol residual y los porcentajes de asimilación para cada cepa de acuerdo a la fórmula: Figura Nº 8. Fórmula porcentaje de asimilación de colesterol en muestras de medio YPGCHO. Fuente (Autores, 2007) Donde: %AC = Porcentaje de asimilación de colesterol Ci = Concentración inicial de colesterol del medio YPGCHO Cf = Concentración final de colesterol en la muestra/12 horas de incubación 48 • Sonicación Se inocularon 10 mL de medio YPGCHO con 0.1 mL (1% (v/v)) de un cultivo de levaduras tras 12 horas de incubación, el control usado fue caldo YPG sin colesterol, la mezcla se llevó a incubar a 30ºC durante 12 horas. Posteriormente, se tomaron 2 mL que fueron centrifugados y resuspendidos en PBS. Las muestras fueron sometidas a sonicación en tiempos de 0, 1, 2 y 3 minutos, se realizaron diluciones de 10-1 hasta 10-2, se realizó coloración supravital con azul de lactofenol y recuento en Cámara de Newbauer (Psomas et al, 2003) donde se expresó el resultado como Levaduras/mL, de acuerdo a la siguiente fórmula: Figura Nº 9. Fórmula Recuento (Levaduras/mL) en cámara de Newbauer. Fuente (Madigan et al, 1998) 5.1.2.5. Prueba de adherencia La línea celular Caco-2 se incubo 37ºC a una atmósfera de 5% CO2 - 95% aire, utilizando un medio mínimo esencial hasta observar la monocapa. Posteriormente, las células fueron lavadas tres veces con PBS. La cepa de la levadura se incubó en medio YPG a 30 oC durante 12 horas a 150 rpm; las levaduras se centrifugaron y se lavaron dos veces con medio basal, 0.8 mL de estos fueron inoculados a las células Caco-2. A continuación este montaje se dejo a 37oC por 90 minutos a una atmósfera de 5% CO2 - 95% aire, para ser posteriormente lavada con PBS cuatro veces y realizando posteriormente una coloración con 1 mL de colorante de Wrigth (Anexo 10) mas metanol por 5 minutos, y lavando nuevamente con agua destilada hasta retirar el exceso de colorante. Las células fueron observadas en el microscopio invertido contando el número de levaduras adheridas a las células Caco-2 en 20 campos microscópicos aleatorios. La capacidad de adhesión se expresó como el número de levaduras adheridas a 100 células Caco- 2 (Kumura et al, 2004). 49 6. RESULTADOS Y DISCUSIÓN DE RESULTADOS 6.1. Control de calidad bancos cepa A (estudio) y cepa B (control). De los inóculos y de las colonias crecidas en YPG, que presentaron la morfología macroscópica de levaduras, es decir, colonias grandes de aproximadamente 1 a 2 mm de diámetro, convexas, lisas, de bordes definidos, brillantes (Anexo 1.1), se realizó coloración de Gram donde se observaron las levaduras en gemación, como se evidencia en la Figura Nº 10 y se reporta en el Anexo 1.1. Figura Nº 10. Aspecto microscópico de la cepa A (estudio) En la Figura Nº 11, se grafican los resultados de los recuentos obtenidos para los controles de calidad de los bancos de la Cepa A y B en función del tiempo. 10,00 Log (UFC/mL) 9,00 8,00 Cepa A 7,00 Cepa B 6,00 5,00 4,00 0 1 2 3 4 5 6 Controles Figura Nº 11. Curva de viabilidad (UFC/mL) en función del tiempo para cepa A (estudio) y cepa B (control). 50 Al observar los resultados presentados en la Figura anterior y el Anexo 2.2, se aprecia que la viabilidad del banco de la cepa A decayó drásticamente a partir del tercer mes de haberse realizado. En la cepa B también se observó disminución en los recuentos, sin embargo se mantuvo en el mismo ciclo logarítmico (108 UFC/mL). La disminución en la viabilidad pudo deberse a varios factores, por ejemplo al realizarse el congelamiento a una velocidad mas alta que la óptima se incrementa el daño celular al producirse la formación de cristales de hielo intracelularmente; también se puede presentar injuria cuando en el proceso de congelación se da la precipitación de las proteínas solubles de la membrana plasmática o cuando se presenta una denaturación en su estructura terciaria, debilitando las uniones hidrofóbicas, la cual puede llegar a ser irreversible si existe la formación de hielo extracelular que genera la deshidratación celular (Marín, 2003). Otra posible razón por la cual pudo haberse presentado la disminución de la viabilidad, es el cambio que se presentó en la temperatura de congelamiento en la que se mantuvo este banco, ya que inicialmente se encontraba a -70oC y posteriormente los viales fueron transferidos a un congelador a -20oC, temperatura no aconsejable, ya que debido a la gran concentración de solutos que existen en la suspensión celular, el punto de congelación baja, por lo que se requiere de una menor temperatura para llegar al punto de congelación lo que puede aumentar la posibilidad de la pérdida en los niveles de viabilidad (Marín, 2003). Otra de las posibles causas de este efecto en las células, es el descongelamiento de manera lenta lo que pudo llegar a ocasionar daños a nivel celular (Marín, 2003), lo que coincide con el estudio realizado por Dumont et al, 2003 donde al someter a la levadura S. cerevisiae a cuatro rangos de congelación a diferentes tiempos se evidenció que la congelación lenta provocaba una alta mortalidad en la cepa de estudio al comprobar su recuperación, mientras que a tiempos de congelación rápida se preservaba la viabilidad celular. Sin embargo se recomienda una velocidad de congelación intermedia la cual beneficia la viabilidad de las levaduras, ya que contribuye a que la salida de agua de la célula ocurra de manera lenta previniendo la congelación al interior de la levadura ocasionando la formación de cristales de hielo al interior de la misma (Dumont et al, 2004). En lo referente a la pureza del banco, para cada cepa se realizaron coloraciones de Gram después del tiempo de incubación, en donde se evidenció la morfología microscópica característica de este microorganismo. Al realizar la coloración de las colonias típicas crecidas en agar YPG, se obtuvieron los mismos resultados. Fue posible evidenciar que la 51 obtención del microorganismo conservado es buena, ya que inoculado el vial a un medio que le brinda condiciones favorables de crecimiento a la levadura, como el caldo YPG y la posterior siembra en caja, se evidenció el crecimiento de este microorganismo, es decir, que sigue conservando su funciones metabólicas sin sufrir alteración. Se puede atribuir la conservación de las levaduras, a que en bajas temperaturas, S. cerevisiae, aumenta la expresión del gen OLE1 que codifica una desaturasa (Ole1p), que convierte los ácidos grasos palmítico y esteárico en los correspondientes ácidos grasos monoinsaturados (ácido palmitoleico y ácido oleico), lo que aumenta la instauración de la membrana, alterando la fluidez de la membrana en respuesta a diferentes tipos de estrés, incluido la disminución de temperatura (Sturkie et al, 1989). El crioprotectante usado en la preservación del banco (glicerol al 30% (v/v)), también preserva las células. El glicerol ha demostrado ser una de las mejores sustancias por cuanto sus características moleculares le permiten simular una vitrificación alrededor del microorganismo, lo cual impide que la formación de cristales de hielo lesione las membranas citoplasmáticas (Dumont et al, 2004, Sánchez et al, 2005). Adicionalmente, al no ser un preservante iónico evita la excesiva formación de hielo, reduciendo el riesgo de daño celular. Se puede concluir que, bajo condiciones controladas de congelamiento el glicerol es un buen preservante ya que la disminución en la viabilidad se presentó por problemas en la temperatura de almacenamiento y adicional a lo anterior, se puede inferir que la concentración usada 30% (v/v), es óptima ya que al almacenar los viales a temperatura de -70ºC se observan resultados que demuestran que la viabilidad se mantiene, por encima de los valores que se consideran ideales (mínimo 106 UFC/mL), lo que se corrobora con el trabajo realizado por Monroy et al (2004), donde se evaluó durante 6 meses la estabilidad a tiempo real de cuatro cepas criopreservadas en 10% (v/v) de glicerol entre las que se encontraba una cepa de la levadura en estudio S. cerevisiae y en donde se observó que la levadura mantuvo una viabilidad del 95%. 6.2. Tolerancia a sales biliares • Recuento inicial de inóculo: El inóculo inicial se mantuvo en 106 UFC/mL durante 12 horas a 30 ºC, 150 rpm para cada cepa. Se realizó coloración de Gram para verificar pureza como se observa en la Tabla 1, en donde se evidenció la morfología característica de esta levadura sin contaminación. 52 Tabla Nº 1. Coloración de Gram y valores de Absorbancia620 nm de inóculos de cepa A (estudio) y cepa B (control) Absorbancia Cepa Microscopía observada a partir de la coloración de Gram del inóculo 620nm A Levaduras sin contaminación microbiana 0.881 B Levaduras sin contaminación microbiana 0.892 Después del período de incubación, en el agar YPG, se evidenciaron colonias grandes de aproximadamente 1 a 2 mm de diámetro, convexas, lisas, de bordes definidos y brillantes. Los recuentos obtenidos se observan en la Tabla 2. Tabla Nº 2. Siembra en superficie en agar YGC de inóculos de cepa A (estudio) y cepa B (control) CEPA UFC/mL A 1.70 E+07 B 2.20 E+07 • Recuentos de Prueba de Tolerancia en Sales Biliares: Los resultados obtenidos se observan en la Figura 12 y en el Anexo Nº 3, en donde se reportan los recuentos (UFC/mL) obtenidos para la cepa estudio (A) y la cepa control (B). 7,50 Log (UFC/mL) 7,00 6,50 Cepa A 6,00 Cepa B 5,50 5,00 4,50 0,05% 0,10% 0,15% 0,20% 0,25% 0,30% Concentracion de sales biliares Figura Nº 12. Recuentos (UFC/mL) en sales biliares para cepa A (estudio) y cepa B (control) 53 Después del período de incubación (24 horas), en las condiciones previamente establecidas, los resultados para las dos cepas, muestran que el género Saccharomyces es capaz de crecer en diferentes rangos de concentraciones de sales biliares similares presentes en el sistema digestivo sin que se altere o afecte su metabolismo. No existió diferencia para concentraciones de 0.05% a 0.3 % (p/v) en los recuentos obtenidos. Para la cepa A, se evidenció que el crecimiento fue constante y estable, manteniéndose en 106 UFC/mL, resultados similares a los presentados para la cepa B (control), ya que no se observó una diferencia estadísticamente significativa (p > 0.05). Para ambos microorganismos se evidencia una disminución del crecimiento para la concentración de 0.3% (p/v) de sales biliares, sin embargo se encuentra en el mismo ciclo logarítmico, de lo que se puede deducir que la reducción en la viabilidad, incluso en la mayor concentración no es drástica debido a que, aunque la levadura está activando los mecanismos de resistencia a estos compuestos, como acumulación de polioles y glicerol, el valor es muy alto y tiende a retardar el metabolismo de la cepa, al disminuir la tasa de producción de metabolitos para su funcionamiento (Rose, 1987). Fietto et al (2004) reportó que se presentó una mejor tolerancia por parte de S. cerevisiae a las sales biliares, soportando un máximo de 0.15% (p/v) comparado con S. cerevisiae var. boulardii, que mostró ser muy sensible incluso a bajas concentraciones, por lo que deduce que ninguna de las cepas en estudio tienen tolerancia a estos compuestos, ya que según Gilliland et al (2004), la concentración que debe resistir un microorganismo probiótico es de 0.3% (p/v) y la mayoría de las especies de levadura crecen incluso en presencia de 0.75% de sales biliares. Sin embargo, para el presente estudio se encontró que hasta en la mayor concentración se presentó crecimiento de las levaduras en 24 horas de incubación, por lo que la cepa A (estudio), cumple con los requerimientos de un microorganismo probiótico. Según Kumura et al (2004), al exponer diversas levaduras a presencia de bilis, estos microorganismos fueron capaces de crecer, siempre y cuando se incubaran a 27ºC, sin embargo las cepas de Saccharomyces sp que fueron estudiadas podían crecer en presencia de sales biliares a 37ºC, temperatura en la que otras levaduras ya se inhibían. Esta es una de las ventajas que se observa de la levadura frente a otros microorganismos probióticos como es la capacidad de crecer a 37 ºC, temperatura relativamente alta para este género, sin que se vea afectado su metabolismo. 54 Según el estudio de Kühle et al (2005), las cepas de S. cerevisiae y S. cerevisiae var. boulardii mostraron resistencia a concentraciones de 0.3 % (p/v), demostrando la sobrevivencia al ser expuestas a concentraciones de sales biliares. En el estudio se observó que el 65% de las cepas de S. cerevisiae y todas las cepas de S. cerevisiae var. boulardii crecían incluso después de las 4 horas de incubación, lo que se pudo verificar en este proyecto ya que el tiempo de incubación fue de 24 horas, obteniéndose recuentos que mostraron que no disminuyó drásticamente la viabilidad de las cepas. El inóculo del que se partió, para ambas cepas se encontraba en 107 UFC/mL, y se evidenció la disminución en una unidad logarítmica después del tiempo de incubación. Los microorganismos probióticos que hacen parte de un biopreparado comercial deben encontrarse en 106 – 108 UFC/mL, al llegar a las células blanco, para asegurar que puedan actuar sobre el huésped (Cavazzoni, 1998), lo que sugiere que es necesario para una evaluación in vivo, partir de un inóculo con una concentración mayor de biomasa, para asegurar que las levaduras al ser enfrentadas a sales biliares, que tienen actividad antimicrobiana, disminuyan en un porcentaje la viabilidad celular, pero se conserven la concentración microbiana necesaria que puedan afectar benéficamente al huésped (Salminen et al, 1999). Los ácidos grasos insaturados juegan un rol esencial en las características de la membrana celular y determinan la función de las proteínas unidas a membrana, que es esencial en la resistencia a las sales biliares (Begley, 2006). Se ha encontrado la presencia de vesículas similares a las encontradas en mamíferos en las levaduras, que pueden internalizar las sales para su posterior degradación (St. Pierre et al, 1994). Una de las reacciones ya caracterizadas en microorganismos como Lactobacillus sp, Listeria sp y Bifidobacterium, es la presencia de enzimas hidrolasas, que actúan sobre el enlace amida liberando glicina y taurina de la base esteroide; este proceso es conocido como deconjugación, dando lugar a los ácidos biliares deconjugados (Begley, 2006). La actividad de transporte dependiente de ATP es esencial para la translocación de muchos componentes de la bilis. Existe un tipo de proteínas unidas a ATP (proteínas ABC), que hacen parte de una familia de proteínas integrales de membrana, responsables de la translocación a través de la misma, de una enorme variedad de substratos que se encuentran presentes desde las bacterias hasta el hombre. La expresión de las proteínas ABC, denominadas MDR1, MDR2 y MRP1, han sido estudiadas en S. cerevisiae, ya que se encuentran implicadas en el transporte de ácidos biliares y sus conjugados. La translocación 55 de glutionato (ácido deconjugado) es mediado por una proteína dependiente de ATP denominada Ycf1p, que se encuentra en una vacuola de la levadura; este organelo es un compartimiento acídico, rico en enzimas catabólicas que juega un rol importante en la acumulación y homeostasis de calcio, fosfato y aminoácidos (Ortiz, 1997), lo que genera una acumulación intravesicular de ácidos biliares (St. Pierre et al, 1994). Estas proteínas ABC, pueden transportar eficientemente ácidos biliares conjugados que deconjugados (Ortiz, 1997). Otros de los mecanismos por el cual la levadura es resistente a altas concentraciones de sales biliares, se centra en la acumulación de polioles y glicerol para regular la presión osmótica de la célula con el medio externo. El glicerol acumulado es expulsado al medio, una vez las sales han sido excluidas del citoplasma (Rose, 1978). Se puede deducir, que la cepa nativa de S. cerevisiae es un microorganismo que es capaz de tolerar concentraciones de sales biliares desde 0.05% (p/v) hasta 0.3% (p/v), pudiendo desarrollar actividades metabólicas, sin verse inhibida o alterada, característica necesaria para la formulación de un biopreparado probiótico dirigido a animales monogástricos, especialmente pollos. 6.3. Tolerancia a rangos de pH Recuento inicial de inóculo: El inóculo inicial se mantuvo en 106 UFC/mL en las condiciones establecidas inicialmente para cada cepa. Se realizó coloración de Gram para verificar pureza como se observa en la Tabla 3, en donde se evidenció la morfología característica de esta levadura sin ningún tipo de contaminación. Tabla Nº 3. Coloración de Gram y valores de Absorbancia620 nm de inóculos de cepa A (estudio) y cepa B (control) Absorbancia Cepa Microscopía observada a partir de la coloración de Gram del inóculo 620nm A Levaduras sin contaminación microbiana 0.821 B Levaduras sin contaminación microbiana 0.854 56 Después del período de incubación, en el agar YPG, se evidenciaron colonias grandes de aproximadamente 1 a 2 mm de diámetro, convexas, lisas, de bordes definidos y brillantes. Los recuentos obtenidos se observan en la Tabla 4. Tabla Nº 4. Siembra en superficie en agar YGC de inóculos de cepa A (estudio) y cepa B (control) UFC/mL CEPA A 1.40 E +07 B 6.30 E +07 • Recuentos de prueba de tolerancia a pH: de cada una de las cepas se realizaron recuentos después de 24 horas de incubación en los diferentes valores de pH (2.0, 2.5, 3.0, 3.5, 4.0, 4.5 y 5.0). 8,00 Log (UFC/mL) 7,00 6,00 5,00 Cepa A 4,00 Cepa B 3,00 2,00 1,00 2 2,5 3 3,5 4 4,5 5 pH Figura Nº 13. Recuentos (UFC/mL) en valores de pH de 2.0 – 5.0 para cepa A (estudio) y cepa B (control). De acuerdo a los resultados presentados en la Figura 13 y en el Anexo Nº 4, se puede evidenciar que las dos cepas, presentaron estabilidad en cuanto a los recuentos realizados a diferentes pH. Asimismo, no se observó una diferencia estadísticamente significativa (p > 57 0.05) entre ambas cepas. A diferencia de la cepa B, la cepa A mantuvo valores de crecimiento en la escala logarítmica de 106 UFC/mL en todos los pH evaluados. La cepa control, desde el pH 2 hasta el pH 4 reportó valores más bajos (105 UFC/mL), y sólo cuando subió el valor de pH a 4.5 pudo aumentar el crecimiento, incluso en 107 UFC/mL a pH de 5.0 debido a que en rangos de pH ácidos, no activa los mecanismos de tolerancia, lo que influye en su metabolismo generando pérdida de viabilidad, y sólo cuando el pH se acerca al óptimo, puede aumentar su crecimiento. La tolerancia a diferentes pH pudo deberse a los antiportadores de Na+/H+ que poseen la levaduras; estos son proteínas encontradas en la membrana citoplasmática así como en las membranas de los organelos de las células. Estas proteínas catalizan el intercambio de cationes monovalentes (Na+ o K+) y H+ a través de las membranas, de tal modo que regulan las concentraciones de cationes y pH a nivel citoplasmático y de organelos (Mitsui et al, 2005). En S. cerevisiae, se han identificado dos tipos de antiportadores de Na+/H+. Uno de los antiportadores es Nha1p, encontrado en la membrana plasmática y el otro es Nhx1p, que se localiza en el compartimiento endosomal prevacuolar (Ohgaki et al, 2005). El antiportador de Na+/H+ Nha1p ubicado en la membrana plasmática es codificado por el gen NHA1. Este antiportador participa en la regulación de las concentraciones intracelulares de Na+ o de K+ usando el gradiente electroquímico de H+ a través de la membrana plasmática; esto elimina exceso de los cationes en el citosol (Sychrova et al, 1999, Brett et al, 2005, Mitsui et al, 2005). Otro de los posibles mecanismos de la levadura para regular el pH y la concentración de iones es una ATPasa, codificada por el gen PMA1 y localizada igual que el antiportador Nha1p en la membrana citoplasmática, esta puede crear un gradiente electroquímico de protones que conduce al transporte secundario de solutos, que esta implicado en el mantenimiento del pH cercano a la neutralidad y que además de ser un componente crítico para la adaptación de la levadura a ácidos, es útil para muchas funciones fisiológicas en la levadura como la toma de nutrientes y la regulación intracelular de la concentración de diferentes cationes y aniones (Viegas et al, 1998, Sychrova et al, 1999, Thomas et al, 2002). Ejemplo de lo anterior, es cuando en condiciones de fermentación el pH externo es de 3.0, S. cerevisiae puede mantener el pH 5.5 y 5.75; igualmente, cuando el pH externo varía entre 6.0 y 10.0, la levadura mantiene el pH entre 5.9 y 6.75 ± 0.2 (Thomas et al, 2002). Para mantener los valores adecuados dentro de un rango óptimo de pH, las células expulsan 58 protones a expensas de ATP, lo que genera la disminución del rendimiento de consumo con respecto a la glucosa que genera un aumento en el pH intracelular, permitiéndole a la levadura resistir rangos amplios (2.4 a 8.6) de pH ácidos (Rose, 1987). En el estudio realizado por Narendranath et al, 2001, se encontró que S. cerevisiae era más resistente a la adición de ciertos ácidos al medio de cultivo, debido posiblemente a que los componentes presentes en el medio de cultivo utilizado (YPG) como extracto de levadura, peptona y glucosa pueden conferirle cierta protección a la levadura bajo condiciones de estrés, lo que pudo también haber contribuido para que los recuentos realizados a los diferentes pH en el presente estudio se mantuvieran estables, a pesar de las condiciones de acidez en el medio. Aunque ambas levaduras toleraron los pH ácidos y presentaron mecanismos que les permiten regular el pH interno, la cepa en estudio demostró una mayor resistencia a esta condición, que la cepa control, resultados que se pueden observar y comparar en el estudio llevado a cabo por Kuhlé et al (2005), en el que el crecimiento a pH de 2.5 fue observado en el 44% de las cepas de S. cerevisiae aisladas a partir de alimentos, a diferencia de S. cerevisiae var. boulardii que no pudo replicarse muy bien en estas condiciones. Sin embargo, todas las levaduras demostraron que podían sobrevivir incluso después de 4 horas de incubación en un pH de 2.5. Los resultados obtenidos para este proyecto, muestran que la cepa en estudio y control pueden tolerar, incluso pH de 2.0, por períodos de incubación de 24 horas, sin que la pérdida de viabilidad sea drástica. Los resultados obtenidos son adecuados, ya que por referencia bibliográfica, un microorganismo probiótico debe encontrarse en 108 UFC/mL al ser suministrado y al llegar a los enterocitos, debe encontrarse viable, con una pérdida máxima de viabilidad en dos unidades logarítmicas (Cavazzoni, 1998). 6.4. Resistencia a jugos gástricos • Recuento inicial de inóculo: El inóculo inicial se mantuvo en 106 UFC/mL en las condiciones establecidas inicialmente para cada cepa. Se realizó coloración de Gram para verificar pureza como se observa en la Tabla 5, en donde se evidenció la morfología característica de esta levadura sin contaminación. 59 Tabla Nº 5. Coloración de Gram y valores de absorbancia620 nm de inóculos de cepa A (estudio) y cepa B (control) Absorbancia Cepa Microscopía observada a partir de la coloración de Gram del inóculo 620nm A Levaduras sin contaminación microbiana 0.866 B Levaduras sin contaminación microbiana 0.874 Después del período de incubación, en el agar YPG, se evidenciaron colonias grandes de aproximadamente 1 a 2 mm de diámetro, convexas, lisas, de bordes definidos y brillantes. Los recuentos obtenidos se observan en la Tabla 6. Tabla Nº 6. Siembra en superficie en agar YGC de inóculos de Cepa A (estudio) y Cepa B (control) CEPA UFC/mL A 2,20E+07 B 1,50E+07 • Recuentos de prueba de tolerancia a jugos gástricos: de cada una de las cepas se realizaron recuentos durante 24 horas, tomando muestras a la hora 0, 1, 2, 4 y 24; del jugo gástrico de pH 2.0 – 2.3 y del control (jugo gástrico pH 7.0 -7.2) 6,50 Log (UFC/mL) 6,00 5,50 Cepa A Cepa B 5,00 4,50 4,00 0 2 4 6 8 10 12 14 16 18 20 22 24 Tiempo de muestreo (h) Figura Nº 14. Recuentos (UFC/mL) en Jugos Gástricos de pH 2.0 – 2.3 para Cepa A (estudio) y Cepa B (control) 60 Para el caso de estudio, la cepa control (B) y la cepa estudio (A) presentaron una buena viabilidad (105 UFC/mL) durante el tiempo de muestreo (hora 0 hasta hora 24) para el jugo gástrico artificial de pH 2.0-2.3, resultados que se observan en la Figura 14 y Anexo Nº 5. Los recuentos obtenidos en las 4 primeras horas de muestreo, permanecieron similares para ambas cepas (siendo ligeramente mayor para la cepa B), ya que no se observó una diferencia estadísticamente significativa entre ambas cepas (p >0.05) sin embargo, la cepa en estudio tuvo recuentos superiores a partir de la cuarta hora de mantenimiento en condiciones simuladas para jugos gástricos y para la hora 24 este valor siguió aumentando como se observa en la Figura 12, resultados que al ser comparados con el estudio realizado por Fietto et al (2004) en el que al exponer las levaduras (S. cerevisiae y S. cerevisiae var boulardii) a jugos gástricos por un tiempo de 60 minutos, no se observaron diferencias en la viabilidad de las cepas, sin embargo, después de 15 minutos, S. cerevisiae var boulardii presentó mayor resistencia, incluso manteniendo la viabilidad en un 75%, mientras que S. cerevisiae presentó una disminución cercana al 30% después de 60 minutos; demuestran que la cepa en estudio presenta una muy buena resistencia a jugos gástricos, porque incluso a las 24 horas de incubación, pudo mantener su metabolismo y la pérdida de viabilidad no fue drástica (105 UFC/mL). Este autor notó que las levaduras (S.cerevisiae var. boulardii) expresan proteínas específicas en respuesta a condiciones de estrés, incluida la resistencia a jugos gástricos, lo que sugiere la relación con la adaptación celular y la sobrevivencia en este ambiente (Fietto et al, 2004). Estos resultados, tienen correlación con los datos observados en la Figura 10 para la prueba de tolerancia a rangos de pH, ya que S. cerevisiae var. boulardii (control), presentó un menor crecimiento hasta pH de 4.5, comparada con la cepa estudio, deduciendo que la levadura nativa, presenta mejores mecanismos de resistencia a condiciones adversas como valores ácidos de pH, a pesar de que la levadura administrada en una sola dosis, se aloja y permanece en el tracto gastrointestinal, pero en concentraciones menores (5 x 107 células/g heces) a las que se albergan en los microorganismos naturales de la flora intestinal dominante, lo que demuestra la capacidad de esta levadura para adaptarse a las condiciones fisicoquímicas del medio (Bunts, 2005). La primera barrera que un microorganismo tiene que atravesar después de ser ingerido es un pH extremadamente ácido, producto de la digestión gastrointestinal. El jugo gástrico que es secretado diariamente posee un pH de aproximadamente 2.0 y la resistencia a un pH bajo como se explicó anteriormente pudo deberse a la presencia de antiportadores que ayudan al 61 intercambio de cationes monovalentes a través de las membranas, regulando el pH a nivel citoplasmático y de organelos, así como de una ATPasa la cual cumple con la misma función (Thomas, 2002). Adicionalmente, el jugo gástrico presenta una concentración de sales de aproximadamente 0,5% (p/v), que generan la activación de mecanismos de resistencia como los mencionados en el numeral 6.2, debido a proteínas de membrana que internalizan estas sales y a vacuolas que participan en su posterior degradación. Esta secreción presenta a su vez una barrera enzimática compuesta principalmente por enzimas proteolíticas; todos estos factores forman una barrera que puede llegar a ser letal para la mayoría de los microorganismos (Martins et al, 2005). Debido a lo anterior, si un organismo sobrevive a estas condiciones extremas se puede considerar que puede llegar a tener potencial para ser usado como probiótico. 6,50 Log (UFC/mL) 6,00 5,50 Cepa A Cepa B 5,00 4,50 4,00 0 2 4 6 8 10 12 14 16 18 20 22 24 Tiempo de muestreo (h) Figura Nº 15. Recuentos (UFC/mL) en Jugos Gástricos de pH 7.0 – 7.2 para Cepa A (estudio) y Cepa B (control) A diferencia de los resultados obtenidos anteriormente, para el jugo gástrico control de pH 7.0-7.2, se observó un mayor crecimiento de la cepa B, comparada con la A, como se observa en la Figura 15 y en el Anexo Nº 4. Sin embargo, no se observó una diferencia estadísticamente significativa (p > 0.05) entre ambas cepas. En las primeras horas los recuentos se mantuvieron similares para cada cepa, sin embargo, desde la hora 4 la cepa control incrementó su viabilidad, mientras que la de la cepa estudio se mantuvo estable, lo que confirmó que la cepa control (S. cerevisiae var. boulardii), crece mejor en pH cercanos a 62 la neutralidad, ya que en condiciones ácidas se disminuye su viabilidad, comparado con la cepa en estudio que tanto en el jugo gástrico de pH 2.0 – 2.3 como en el control de pH 7.0 – 7.2, mantuvo su viabilidad en valores similares, sin observarse diferencias en los recuentos (UFC/mL) obtenidos, como se reporta en el Anexo Nº 5. 6.5. Reducción de colesterol en presencia de sales biliares. • Elaboración curva patrón de colesterol: Para la determinación de la curva patrón, se midió la absorbancia550mn de cada solución con concentración de colesterol conocida (60 – 130 μg/mL), de acuerdo al principio en el cual las sustancias tienen propiedad para absorber o emitir una longitud de onda (Chang, 1999). Esta determinación se realizó a 550 nm, que es el valor máximo de absorción. Ya que los datos se agruparon alrededor de la línea y para obtener la ecuación de la recta de acuerdo a la ecuación de la línea recta: y=mx+b, se aplicó el método matemático denominado regresión lineal, que correlaciona los datos. La gráfica relaciona los valores de absorbancia (eje Y) y la concentración de colesterol (eje X) y se hace evidente la Ley de Lambert - Beer, según la cual existe una relación directa entre ambas funciones, al ser directamente proporcionales, ya que ningún valor excedió los límites de lectura (0.1 – 0.9 absorbancia). A bsorbancia (550 nm ) 0,5 0,4 0,3 y = 0,0045x - 0,1564 2 0,2 R = 0,9937 0,1 0 50 70 90 110 130 150 Concentración colesterol (μg/ml) Figura Nº 16. Curva patrón de colesterol (60μg/mL – 130μg/mL) 63 Esta ley sólo es aplicable a soluciones diluidas de las especies absorbentes, que no sobrepasen dicho límite, puesto que la presencia de muestras con concentraciones muy altas, origina la desviación lineal entre la absorbancia y la concentración del analito (Skoog, 1988). De acuerdo a esto, la ecuación de la recta que se obtuvo al relacionar las dos funciones de los ejes X y Y, fue y= 0.0045x + (- 0.1564) y R2 =0.9937, resultados observados en la Figura 16 y en el Anexo Nº 6.2, en donde se observan los resultados obtenidos en las réplicas y el promedio de los valores de absorbancia. Lo anterior se confirma con el R2 obtenido de la regresión lineal, igual a 0.9937, que es el valor de correlación de los datos y muestra que tan dispersos o no son los valores alrededor de la línea. Este debe ser igual a 1 o cercano a este número, para que la ecuación obtenida pueda ser usada, De acuerdo al valor obtenido para la curva patrón elaborada, se deduce que si es posible su uso para la determinación de la reducción del colesterol en presencia de las cepas. 6.5.1. Determinación de reducción Colesterol en presencia de sales biliares • Recuento inicial de inóculo: El inóculo inicial se mantuvo en 106 UFC/mL en las condiciones establecidas inicialmente para cada cepa. Se realizó coloración de Gram para verificar pureza como se observa en la Tabla 7, en donde se evidenció la morfología característica de esta levadura sin contaminación Tabla Nº 7. Coloración de Gram y valores de absorbancia 620 nm de inóculos y de cepa A (estudio) y cepa B (control) Cepa Microscopía observada a partir de la coloración de Gram Absorbancia del inóculo 620nm A Levaduras sin contaminación microbiana 0.866 B Levaduras sin contaminación microbiana 0.874 Después del período de incubación, en el agar YPG, se evidenciaron colonias grandes de aproximadamente 1 a 2 mm de diámetro, convexas, lisas, de bordes definidos y brillantes. Los recuentos obtenidos se observan en la Tabla 8. 64 Tabla Nº 8. Siembra en superficie en agar YGC de inóculos de cepa A (estudio) y cepa B (control) • CEPA UFC/mL A 5.00E +07 B 4.10E +07 Reducción del colesterol: Se determinó la concentración inicial de colesterol experimental midiendo el valor de absorbancia550nm el cual fue de (0.839), este fue reemplazado en la ecuación de la recta y se obtuvo un dato correspondiente a 221,2 μg/mL. Posteriormente, se procesaron las muestras de la cepa A (estudio) y la cepa B (control), encontrándose para la primera un valor promedio de absorbancia de 0.303 y de 0.265 para la segunda y no se observó diferencia estadísticamente significativa (p > 0.05) entre ambas cepas. Estos valores fueron despejados en la ecuación de la recta obtenida de la curva patrón y se obtuvieron datos correspondientes al colesterol residual en la muestra, siendo de 102 μg/mL (cepa A) y de 93.64μg/mL (cepa B). Con estos valores se determinó el colesterol degradado restando de la concentración inicial experimental (221.2 μg/mL). Posteriormente, se procesaron las muestras de la cepa A (estudio) y la cepa B (control), encontrándose para la primera un valor promedio de absorbancia de 0.303 y de 0.265 respectivamente, valores que se reportan en el Anexo Nº 6.3. Estos valores fueron despejados en la ecuación de la recta obtenida de la curva patrón (y =0.0045x – 0.1564, R2= 0.9937) y se obtuvieron datos correspondientes al colesterol residual en la muestra, siendo de 102 μg/mL (cepa A) y de 93.64μg/mL (cepa B). Con estos valores se determinó el colesterol degradado, como se observa en la Tabla 9. Tabla Nº 9. Valores de colesterol degradado en 12 horas de incubación por la cepa A (estudio) y la cepa B (control) Colesterol degradado / 12 horas de incubación Cepa A 119.2μg/mL Cepa B 127.56 μg/mL 65 • Porcentajes de asimilación de colesterol: después de determinar la cantidad de colesterol degradado, se determinó el porcentaje de asimilación de cada cepa, de acuerdo a la fórmula del numeral 5.1.2.4. Los resultados obtenidos se observan en la Tabla Nº 10. Tabla Nº 10. Porcentajes de asimilación de colesterol/12 horas de incubación por la Cepa A (estudio) y la Cepa B (control) Cepa Porcentaje de asimilación/12 horas A 54% B 58% De acuerdo a lo que se presenta en los resultados se puede observar que las dos cepas de levaduras en evaluación fueron capaces de asimilar más del 50% del colesterol presente en el medio en 12 horas de incubación. Esto se corrobora con estudios como el realizado por Psomas et al (2003) en donde varias cepas de S. cerevisiae luego de 48 horas de incubación a 37oC mostraron un porcentaje de asimilación del 100 %. Los resultados obtenidos muestran que la cepa en estudio es capaz de reducir el colesterol, característica que es deseable, ya que para humanos la hipercolesteremia o elevados niveles de colesterol en la sangre, es considerada como el mayor riesgo del desarrollo de enfermedades al corazón y en animales, la menor presencia de colesterol genera mejores carnes, de elevada calidad y de mayor demanda, al encontrarse libres de grasas. La administración de probióticos ha demostrado que puede reducir notablemente los niveles de colesterol (De Smet et al, 1998, Du Tuit et al, 1998, Taranto et al, 1998). Es importante la adición de las sales biliares al medio de cultivo adicionado de colesterol, ya que cuando estuvieron ausentes, no fue posible realizar la extracción de las muestras para la curva patrón. Lo anterior debido a que las sales biliares se encuentran presentes en el organismo en actividades de emulsión, solubilización y absorción de lípidos en el intestino (Begley, 2006). Los métodos de determinación del colesterol incluyen el analizar las muestras por cromatografía de gas, al ser un método confiable, rápido y eficaz (Fleutoris et al, 1998). Sin embargo, cuando no se cuenta con esta opción existe un protocolo basado en la reacción de Liebermann – Burchard, en la que posterior a la adición del hexano, se adiciona ácido glacial 66 acético el cual reacciona con el grupo C3 hidroxilo del colesterol en presencia de ácidos fuertes (ácido sulfúrico concentrado) y forma el complejo azul-verdoso y en su defecto rojo/café que se evidenció durante el montaje como se observa en la Figura Nº 17 (Skoog, 1998). De lo anterior, se concluye que este protocolo de extracción es adecuado, ya que permitió obtener valores de colesterol en relación a los valores de Absorbancia (550 nm), ya que al realizar la curva patrón se obtuvo un R2 igual a 0.9937, lo que evidenció que los datos se relacionaban entre sí, además los resultados obtenidos en la determinación de la reducción del colesterol pudieron ser correlacionados con el reportados por Psomas et al (2003). Figura Nº 17. Reacción de Liebermann – Burchard para determinación de Colesterol. Fuente (Autores, 2007) • Sonicación: Las levaduras en Medio YPGCHO y caldo YPG después de 12 horas de incubación fueron sometidas a sonicación por períodos de 0, 1, 2 y 3 minutos, posterior al periodo de incubación se realizaron recuentos en cámara de Newbauer. El objetivo de esta prueba fue verificar si efectivamente las levaduras asimilaban el colesterol a su membrana alterando su estructura y confiriéndole mayor resistencia a condiciones de estrés tal como la lisis celular generada por este protocolo, por lo que al exponer el microorganismo a este método de disrupción, se esperaba que las levaduras inoculadas en medio mas colesterol cambiarán la morfología de la membrana presentando recuentos más altos en comparación con aquellos que no fueron inoculados en medio suplementado. 67 Log Levaduras/mL 8 7 6 5 Cepa A con colesterol 4 Cepa A sin colesterol 3 2 1 0 0 1 2 3 Tiempo (minutos) Figura Nº 18. Recuentos (Levaduras/mL) de Cepa A (estudio) expuesta a sonicación En ambas cepas se observaron recuentos mayores (Levaduras/mL) cuando estas habían sido inoculadas en medio YPGCHO en comparación con los recuentos de levaduras que habían sido inoculadas en medio sin suplemento de colesterol. Para la cepa A, como se observa en la Figura 18, se obtuvieron valores en la escala de 106 levaduras/mL para el tiempo 0, lo que confirma que el microorganismo sigue viable tanto en el medio con colesterol o sin ser suplementado, sin embargo al ser expuesta S. cerevisiae a la sonicación por 1 y 2 minutos se observó que la viabilidad de la cepa disminuye en una unidad logarítmica, ya que se reportan valores de 105 levaduras/mL, siendo levemente más altos los valores de la cepa inoculada en Medio YPGCHO; para el tiempo 3 incluso disminuye drásticamente la viabilidad hasta 102 levaduras/mL en el caldo inoculado sin suplemento de colesterol a diferencia de la levadura en medio suplementado, que sigue conservando la viabilidad en 105 levaduras/mL. Resultados similares se observan en la Figura 19, para los recuentos de la cepa B, presentándose disminución en la viabilidad tras la sonicación, siendo siempre más altos los valores de S. cerevisiae var. boulardii (cepa B) inoculada en Medio YPGCHO, que en caldo YPG sin adición de colesterol, para el último tiempo se observa igual la disminución hasta 102 levaduras/mL. 68 Log Levaduras/mL 8 7 6 5 Cepa B con colesterol 4 Cepa B sin colesterol 3 2 1 0 0 1 2 3 Tiempo (minutos) Figura Nº 19. Recuentos (Levaduras/mL) de Cepa B (control) expuesta a sonicación De acuerdo al estudio realizado por Psomas et al (2003) se sabe que posiblemente la reducción del colesterol en un medio con levaduras, se deba a que estos microorganismos asimilan el colesterol para incorporarlo a su estructura, reportándose que las levaduras expuestas a medio de cultivo enriquecido con colesterol eran más difíciles de lisar tras ser sometidas a sonicación que las levaduras que no crecían en el medio enriquecido con colesterol, lo que sugiere un posible cambio morfológico en la pared (Kollár et al, 1997), ya que el microorganismo al incorporar este esterol a su estructura se hace más resistente a la lisis celular, comparado con los que no lo que no lo incorporan. 6.6. Ensayo de adhesión a células Caco – 2 • Recuento inicial de inóculo: El inoculo se mantuvo en 106 UFC/mL en las condiciones establecidas inicialmente. Se realizó coloración de Gram para verificar pureza como se observa en la Tabla 11, en donde se evidenció la morfología característica de las levaduras sin contaminación. 69 Tabla Nº 11. Coloración de Gram y valores de absorbancia 620 nm de inóculos y de cepa A (estudio) y cepa B (control) Cepa Microscopía observada a partir de la Coloración de Gram Absorbancia del inoculo 620nm A Levaduras sin contaminación microbiana 0.890 B Levaduras sin contaminación microbiana 0.861 En el agar YPG se evidenciaron colonias grandes de aproximadamente 1 a 2 mm de diámetro, convexas, lisas, de bordes definidos y brillantes. Los recuentos obtenidos se observan en la Tabla 12. Tabla Nº 12. Siembra en superficie en agar YGC de inóculos de cepa A (estudio) y cepa B (control) CEPA RECUENTO (UFC/mL) A 9.80E +07 B 8.80E +07 • Obtención de la monocapa: fue posible obtener la monocapa de las células Caco – 2 al ser cultivadas en Medio Mínimo esencial de Eagle’s (Sigma ®) adicionado de SFB al 10%, Penicilina y L-Glutamina 4.4 mM. Al ser fijadas y teñidas con el colorante de Wrigth adicionado de metanol (Anexo 10) y observar en microscopio invertido, se evidenció la morfología de las células redondeadas como se detalla en la Figura 20. Figura Nº 20. Células Caco – 2 en monocapa con coloración de Wrigth. 70 Al cultivar células es importante tener en cuenta factores como el tiempo de cultivo y la densidad, ya que esto puede influir en la replicación y diferenciación y deben ser estandarizados para obtener modelos experimentales reproducibles y comparables. El medio de cultivo puede influir en las características físico – químicas y su composición puede modular funciones de las células como proliferación, diferenciación, permeabilidad e incluso su morfología. La adición de Glutamina es un factor importante, ya que esencial para el mantenimiento de la homeostasis del intestino, así como en la estimulación de factores de crecimiento necesarios para la células como proteínas y moléculas de adhesión tales como integrinas y E-cadherinas (Sambuy et al, 2005). Adicional a esto, se observó que la presencia de SFB es importante para el crecimiento de las células, ya que al mantenerlas con medio suplementado con Glutamina y Penicilina pero sin suero, se observó que la formación de la monocapa no se daba y las células se observaban desprendidas; esto debido a que este compuesto es una mezcla de biomoléculas pequeñas que promueven el crecimiento celular al contener glucosa, nitrógeno, albúmina, fibronectina, ácido búrico, creatina y hemoglobina (Fresney, 1990), Se puede concluir que el medio utilizado para el cultivo de esta línea celular es adecuado, ya que permitió un buen crecimiento de las células, permitiendo obtener la monocapa necesaria para poder realizar los ensayos de adhesión. • Ensayo de adhesión: El colorante de Wright permitió observar la monocapa formada por las células Caco – 2 ya esta es una tinción de tipo Romanowsky que consiste en la utilización de azul de metileno y eosina Y o eosina B. La acción combinada de estos colorantes da una coloración púrpura a los núcleos de los leucocitos y a los gránulos neutrofílicos y de color rosado a los eritrocitos. Los componentes de este efecto son el azul B y la eosina Y. Los agrupamientos de ácidos nucleicos, las proteínas de los núcleos celulares y el citoplasma inmaduro reactivo, fijan el azul B, colorante básico. La eosina Y, colorante ácido, se fija a los agrupamientos básicos de las moléculas de hemoglobina y a las proteínas básicas. Se observó entonces la coloración púrpura de acuerdo a la unión de los componentes del colorante a las diferentes estructuras de las células Caco – 2 y de las cepas en las Figuras 21y 22. 71 Figura 21. Análisis de adherencia por microscopio invertido de Saccharomyces cerevisiae a células Caco-2 con colorante de Wright. Figura 22. Análisis de adherencia por microscopio invertido de S. cerevisiae var. boulardii a células Caco-2 con colorante de Wright. De este total, se reportaron, de 100 células Caco -2 cuantas presentaron adhesión por parte del microorganismo. Para la cepa A (estudio), se evidenciaron 73 Levaduras adheridas/100 células Caco – 2, a diferencia de la cepa B (control) donde se observaron 41 Levaduras adheridas/100 células Caco – 2, resultados que se pueden observar y comparar en la Figura 23. 72 Levaduras adheridas/100 células Caco-2 80 70 60 50 40 30 20 10 0 A B Cepa Figura Nº 23. Gráfica de barras de Levaduras adheridas/ 100 células Caco -2 de la cepa A (estudio) y Cepa B (control) De acuerdo a los resultados obtenidos se puede observar que ambas cepas presentaron adherencia a la línea celular Caco -2, sin embargo la cepa estudio A presentó mayor adhesión que la cepa B, resultados comparables con los encontrados por Jacobsen et al (1999), estudio en el que se realizaron ensayos de adhesión a células Caco-2 con varias cepas de Lactobacillus, determinando que las cepas que presentaban un recuento de adhesión menor a 40 microorganismos en los 20 campos contados al azar se consideraban como no adherentes, entre 41 y 100 microorganismos como adherentes y mas de 100 microorganismos como fuertemente adherentes. Adicional a este estudio, se pueden comparar los resultados con los encontrados por Kumura et al (2005), en el que las cepas en estudio de Saccharomyces cerevisiae presentaron adhesión a células Caco – 2, aunque en niveles bajos. Kühle et al (2005) reportó que se presentó la capacidad de adhesión más alta para las cepas de S. cerevisiae var. boulardii,en porcentajes del 16.2% y 28.0% y para S. cerevisiae en un 13.6% en células IPEC-J2. Por lo tanto para el presente estudio se puede concluir que ambas cepas, tanto la cepa de estudio como la control se pueden considerar como adherentes a la línea celular Caco-2. Un criterio importante para una cepa probiótica es su habilidad para adherirse a las superficies de la mucosa de tracto gastrointestinal, por lo cual se requiere de estudios in vitro de adherencia los cuales permitan confirmar que una cepa de microorganismos es capaz de cumplir con este criterio. Lo anterior constituye el primer mecanismo de defensa contra la invasión de patógenos. El paso a través del intestino delgado es muy rápido (2.5 horas) y 73 disminuye conforme va llegando al ano. Por lo tanto, para que el microorganismo se pueda multiplicar es necesario que se adhiera a la mucosa y/o epitelio del intestino. Como el epitelio está constantemente regenerándose (cada 3-4 días), las cepas probióticas pueden colonizarlo solo si la tasa de generación es mayor que la tasa de recambio celular (Jonnson et al, 1962). El primer contacto del microorganismo con el huésped en el intestino, es la capa mucosa que recubre al epitelio intestinal y que tiene un grosor variable entre 50 y 450 mm (Lesson et al, 1990). Tiene diferentes funciones, entre las que se encuentran inhibir la adhesión de ciertas bacterias, al mismo tiempo que provee un hábitat para otras (Ouwehand et al, 1996). El principal componente del mucus es agua, mientras que el componente orgánico más importante es la mucina, responsable de la viscosidad del mucus. Las mucinas (MUC) son glicoproteínas de alta masa molecular (0.25-2 megadaltones) constituidas de esqueletos polipeptídicos cuya parte central está altamente glicosilada con O-glicanos principalmente, en grupos de secuencias repetitivas, mientras que los extremos amino y carboxilo presentan una menor glicosilación. Los tipos de mucinas que se encuentran mayormente en el intestino delgado y grueso son MUC2 y MUC3 producidas por las células globosas (Lesson et al, 1990). Mediante diferentes mecanismos los microorganismos se mantienen en el tracto intestinal en contra del flujo del peristaltismo. Uno de los mecanismos usados por Saccharomyces cerevisiae para adherirse a las células epiteliales son las manoproteínas constituyentes de la membrana celular. Estas manoproteinas son un grupo heterogéneo de proteoglicanos con diferentes pesos moleculares y son mejor conocidas como adhesinas (Tuite et al, 1991). Las adhesinas se unen a aminoácidos específicos o residuos de azúcares presentes en la superficie de otras células. (Verstrepen et al, 2006). Las denominadas tipo lectina o sensibles a azúcares, son aquellas que dependen de la unión a un residuo de estos compuestos en la superficie de la célula; en este caso reconocen glicoconjugados específicos en las células epiteliales intestinales, carbohidratos presentes en las mucinas que pueden funcionar como receptores tanto para bacterias patógenas como probióticas, ocurriendo así una competencia por los sitios de adhesión. Esos receptores pueden ser Dgalactosa, N-acetilglucosamina, N-acetilgalactosamina o ácido N-acetilneuramínico asociado a las glicoproteínas intestinales. (Giannasca et al, 1996, Verstrepen et al, 2006). Adicionalmente en S. cerevisiae la adherencia de este tipo se puede dividir en dos subcategorías, una se denomina Flo1 y la otra nuevo Flo, la primera se une a manosa, 74 mientras la segunda se une a azúcares como manosa, glucosa y oligómeros de glucosa como maltosa (Verstrepen et al, 2006). Cuando se habla de adhesión por parte de los microorganismos con capacidad probiótica a las células epiteliales se pone en manifiesto el uso otra ventaja de los probióticos: la exclusión competitiva, esta implica la prevención de la entrada o el establecimiento de una población bacteriana al tracto gastrointestinal debido a la competencia por sustrato y por espacio generado por el grupo de microorganismos que ocupa los potenciales sitios de adhesión. Los microorganismos probioticos compiten directamente o afectan la adhesión de patógenos a los receptores específicos de la mucosa del tracto gastrointestinal (Maija et al, 2006). En microorganismos como Bifidobacterias, la adhesión al intestino ocurre por la asociación de las bacterias con la mucosa o por adherencia al epitelio y se debe a la interacción de fuerzas atractivas y repulsivas entre las superficies participantes. La composición de la pared celular bacteriana, así como la naturaleza de la superficie a la cual se adhiere, afectan este fenómeno (Fontaine et al, 1994). Teniendo en cuenta que en las pruebas de tolerancia a sales biliares, pH, jugos gástricos y en la de reducción del colesterol en presencia de sales biliares no se observaron diferencias estadísticamente significativas (p > 0.05) entre la cepa A (estudio) y la cepa B (control), y que sólo en la prueba de adherencia a la línea celular Caco -2 se observó un mayor porcentaje de adhesión por parte de la Cepa A en comparación con la Cepa B, se puede concluir que la cepa nativa de estudio de Saccharomyces cerevisiae presenta las características necesarias para que pueda llegar a ser empleada en un futuro como suplemento probiótico en la nutrición de animales monogastricos como pollos. 75 7. CONCLUSIONES • Las pruebas de conservación de cepas con glicerol 30% (p/v) demostraron que hacia el tercer mes de haberse realizado el banco la viabilidad disminuyó significativamente. • Se determinó la tolerancia a sales biliares en concentraciones desde 0.5% a 3.0% (p/v) de la cepa estudio (cepa A) y de la cepa control (cepa B), evidenciándose que el género Saccharomyces sp. es capaz de crecer a todas las concentraciones de sales evaluadas. • En la prueba de tolerancia a pH, se encontró que tanto la cepa A (estudio) como la cepa B (control), presentaron estabilidad y viabilidad celular en los rangos de pH evaluados (2.0 a 5.0) • En la prueba de resistencia a jugos gástricos, ambas cepas demostraron estabilidad y viabilidad celular manteniéndose en una concentración de 105 UFC/mL durante los tiempos evaluados. • Se determinó la capacidad de reducción del colesterol para ambas cepas, en donde luego de 12 horas de incubación en un medio suplementado con colesterol se evidencio una reducción del 54% para la cepa A y del 58% para la cepa B. • Se realizó la prueba de sonicación para las dos cepas A y B crecidas en medio con y sin colesterol, en donde se comprobó que las levaduras crecidas en medio con colesterol son más resistentes a la lisis producida por este método de disrupción celular. •En la prueba de adherencia realizada con la línea celular Caco-2, fue posible determinar la habilidad de adherencia por parte de la cepa de estudio A y la cepa control B, siendo ambas cepas consideradas como adherentes. • Se observó que no existió diferencia estadísticamente significativa entre la cepa A y B en las pruebas de tolerancia a sales biliares, pH, jugos gástricos y reducción del colesterol en presencia de sales biliares. 76 • De acuerdo a los resultados obtenidos en las pruebas realizadas para el presente estudio se puede inferir que la cepa nativa de Saccharomyces cerevisiae en estudio, cumple con los requisitos para ser usada como suplemento en probiótico en nutrición animal. 77 8. RECOMENDACIONES • Se recomienda la realización de estudios in vivo a partir de los cuales se pueda determinar los verdaderos efectos de un futuro suplemento probiótico con Saccharomyces cerevisiae en modelos animales específicos. • Al observarse una reducción de colesterol mayor al 50% en 12 horas de incubación por parte de las dos cepas usadas en este estudio, se recomienda la evaluación a tiempos superiores a las 12 horas para determinar si se logra alcanzar una reducción del 100%. • Se recomienda la evaluación de la actividad de la enzima hidrolasa que actúa sobre las sales biliares en Saccharomyces cerevisiae, la cual ha sido encontrada en otros microorganismos con capacidad probiótica como mecanismo de tolerancia. • Se recomienda el estudio de la formulación de un suplemento nutricional usando Saccharomyces cerevisiae como microorganismo probiótico, determinando la concentración y forma más apropiada para su administración en animales monogastricos. • Se recomienda que para el mantenimiento de la viabilidad de las cepas de estudio y la estabilidad del banco de conservación de las mismas la temperatura de almacenamiento se mantenga estable siendo esta preferiblemente de -70oC. 78 9. BIBLIOGRAFIA ABDUDRAHIM, S, M., HADDADIN, M. S. Y., ODETALLAH, N. H. M., ROBINSON, R.K., 1999, Effect of Lactobacillus acidophilus and zinc bacitracin ad dietary additives for broiler chickens., British Poultry Science, Vol 40, P. 91 - 94 ACUÑA, G.A., 1995, Efecto del tratamiento químico del agua de consumo en pollo de engorde sobre un probiótico., Universidad Nacional de Colombia, Facultad de Medicina Veterinaria y Zootecnia, Tesis de Pregrado. P. 22 – 28. AGARWAL, N., KAMRA, D. M., CHAUDHARY, L. C., SAHOO, A. AND PATHAK, N. 2000 Selection of Saccharomyces cerevisiae strains for use as a microbial feed additive. Letters in Applied Microbiology, Vol 31, P. 270 - 273. AGUIRRE, A. POUTOU, R. 2003. Obtención de un biopreparado a partir de cepas nativas de levaduras para ser empleado como probiótico. Pontificia Universidad Javeriana. Facultad de Ciencias. Maestría en Microbiología. Tesis de Postgrado. P. 45 – 49, 72 – 79. ALANDER, M., KORPELA, R., SAXELIN, M., VILPPONEN – SALEMA, T., METTILA – SHANDHOLM, A., von WRIGHT. 1997. Recovery of Lactobacillus rhamnosus GG from human colonic biopsies. Letters of Applied Microbiology..Vol 24. P. 361 – 364. ALANDER, M., SATOKARI, R., KORPELA, R., SAXELIN, M., VILPPONEN – SALEMA, T., METTILA – SHANDHOLM, A., von WRIGHT. 1997. Persistence of Colonization of Human Colonic Mucosa by a Probiotic Strain, Lactobacillus rhamnosus GG, after Oral Consumption Applied and Environmental Microbiology, Vol 65, P. 351 – 354. ALVAREZ, R. J., VALDIVIA, M., 1980. Energía metabolizable y retención de nitrógeno en dietas con levadura torula para pollos de engorde. Revista cubana de ciencia agrícola. Vol 14. P 55. ARORA, D. 1991, Handbook of Applied Mycology the Yeast, Marcel Dekker, New York. P. 42 – 48. ATTC. 1992. Catalogue of cell lines and hibridomas. 7ª edition. American type Culture Collection. 79 BARNETT, J. A., PAYNE, R. W., YARROW, D., 1990. Yeast, characteristics and identification. 2ª edición. Cambridge University Press. New York. USA. P. 72 – 73. BARROW, P., 1992. Probiotics for Chickens. Probióticos. The Scientific Basis. R. Fuller ed. Chapman and Hall editorial. Gran Bretaña. P. 226 – 250. BEGLEY, M., HILL, C., CORMAC, G., GAHAN, M., 2006. Bile Salt Hydrolase Activity in Probiotics, Applied and environmental microbiology, Vol. 72, No. 3, P. 1729 - 1738. BEGLEY, M., HILL, C., CORMAC, G., GAHAN, M., 2006. The interaction between bacteria and bile. FEMS Microbiology, Vol 29, P. 625 - 651. BORRIELLO, S., HAMMES, W., HOLZAPFEL, W., MARTEAU, P., SCHREZENMEIR, J., VAARA. M., AND VALTONEN, V. 2003. Safety of probiotics that contain lactobacilli or bifidobacteria. Clinical Infection Disease, Vol 36, P. 775 - 780. BOUIX, M., LEVEAU, J, Y., 2000. Microbiología industrial: los microorganismos de interés industrial. Editorial Acribia. Zaragoza, España. P. 3 – 89 BRETT, C. TUKAYE, D. MUKHERJEE, S. RAO. R. 2005. The Yeast Endosomal Na+ (K+)/H+ Exchanger Nhx1 Regulates Cellular pH to Control Vesicle Trafficking. Molecular Biology of the Cell. Vol 16, P. 1396 - 1405. BRUGIER, S. AND PATTE, F. 1975. Antagonisme in vitro entre l'U ltraLevure et diffearents germes bacteariens. Meadecin de Paris. 45, P. 61 – 66. BUTS, J.P., BERNASCONI, P., VAN CRAYNEST, M.P., MADALGUE, P. AND DE MEYER, R. 1986. Response of human and rat small intestinal mucosa to oral administration of Saccharomyces boulardii. Pediatric Research Vol 20, P. 192 - 196. BUTS, J. 2005. Ejemplo de un medicamento probiótico: Saccharomyces boulardii liofilizada. Gastroenterology, Vol 25, P 176 - 188. 80 CAMEOTRA, S., S., 2007. Preservation of micro-organisms as deposits for patent application, Biochemical and Biophysical Research Communications, Vol 353, N 4, P. 849 - 850. CASAL, M., CARDOS, H., and LEÃO, C., 1996. Mechanism regulating transport of acetic acid in Saccharomyces cerevisiae. Microbiology, Vol 142, P. 1385 - 1390. CAVAZZONI, V., ADAMI, A., CASTROVILLI, G., 1998, Performance of broiler chickens supplemented with Bacillus coagulans as probiotic, British Poultry Science., 39: 526 – 529 CHANG, L.T. and ELANDER, R. P., 1986. Long-term preservation of industrially important microorganisms. In: DEMAIN, A.L.; SOLOMON, N.A., (Ed.), Manual of industrial microbiology and biotechnology, Washington: CRC Press, cap.5, p.49-55. CHANG, R., 1999, Química, Sexta edición, Mc graw Hill, México D.F., México, P. 244 COLLINS, C, 1989. Métodos microbiológicos, Acribia, Zaragoza, España. P. 155 COUCH, J. R. 1978. Poultry researchers outline benefits of bacteria, fungistatic compounds, other feed addtivies. Feedstuffs, Vol 50, N 14, P. 6. CRAWFORD, J. S. 1979. Probiotics in animal nutrition. Arkansas Nutririon Reference. Proceedings of Natural Academic Science, Vol 1, P. 45-55. DANN, H. M., DRACKLEY, J.K., McCOY, G. C., HUTJENS, M. F., GARRET, J. E.,2000. Effects of yeast culture (Saccharomyces cerevisiae) on prepartum intake and postpartum intake and milk production of Jersey cows, Journal of Dairy Science, Vol 83, N 1, P. 123 127. DAWSON, K., A. 1987. Mode of action of yeast culture, tea Sacc, In: Rumen: a natural fermentation modifier. Biotechnology in food industry, LYONS, T. P., (Ed). Alltech technical. Nicolasville , EUA, P. 119 – 125. DE SMET, I., DE BOEVER, P. and VERSTRAETE, W., 1998. Cholesterol lowering in pigs through enhanced bacterial bile salt hydrolase activity, British Journal of Nutrition, Vol 79, P. 185 - 194. 81 DILWORTH, B. C., DAY, E. J. 1978. Lactobacillus cultures in broiler diets. Poultry. Science, Vol 57, P.1101. DUCLUZEAU, R. AND BENSAADA, M. 1982. Effect compareä de l'administration unique ou en continue de Saccharomyces boulardii sur l'establissement de diverses souches de Candida dans le tractus digestif de souris gnotoxe aniques. Annales de Microbiologie (Institut Pasteur) Vol 133B, P. 491 - 501 DUMONT, F. MARECHAL, P. GERVAIS, P. 2004. Cell Size and Water Permeability as Determining Factors for Cell Viability after Freezing at Different Cooling Rates. Applied and Environmental Microbiology. Vol 70, P. 268 - 272. DUMONT, F. MARECHAL, P. GERVAIS, P. 2003. Influence of cooling rate on Saccharomyces cerevisiae destruction during freezing: unexpected viability at ultra-rapid cooling rates. Cryobiology. Vol 46, P.33 – 42. ENDO, T., NAKANO, M., SHIMIZU, S., FUKUSHIMA, M., 1999, Effects of a probiotic on a lipids metabolism of cocks fed on a cholesterol – enriched diet, Bioscience, Biotechnology and Biochemical, Vol 63, N 9, P. 1569 - 1572. FENNEMA, O.R., Química de los alimentos. 2.ed. Zaragoza: Acribia, 2000. P. 56 -58. FENTON, M. 1992. Chromatographic separation of cholesterol in foods. Journal of Chromatogrophy, Vol 624, P. 369 - 388. FERREIRA, C., FÁVARO, C., 2004, Stability of free and immobilized lactobacillus acidophilus and Bifidobacterium lactis in acidified milk and of immobilized B. lactis in Yoghurt, Brazilian Journal of Microbiology, Vol 35, P. 151 – 156. FIETTO, J., ARAUJO, R., VALADAO, N., FIETTO, G., BRANDAO, R., NEVES, M., GOMES, F., NICOLI, J. and CASTRO, L., 2004. Molecular and physiological comparisons between Saccharomyces cerevisiae and Saccharomyces Boulardii. Canadian Journal of Microbiology, Vol 50, P. 615 – 621. 82 FLETOURIS, D. J.,., BOTSOGLOU, N. A., PSOMAS, I. E., AND.MANTIS, A. I. 1998. Rapid determination of cholesterol in milk and milk products by direct saponification and capillary gas chromatography. Journal of Dairy Science. Vol 81, P. 2833 - 2840. FRESNEY, R., 1990. Cell lines al culture. 3ª edición, EUA, P. Chapter 1, P. 10 – 15. FONTAINE, J.F., E.A. AISSI, AND S.J. BOUQUELET 1994. In vitro binding of Bifidobacterium bifidum DSM 20082 to mucosal glycoproteins and hemagglutinating activity. Current Microbiology, Vol 28, P. 325 - 330. FULLER, R. 1989. Probiotics in man and animals. Journal of Applied Bacteriology, Vol 66, N 5, P. 365 - 378. FULLER, Roy. Probiotics: 1992 The scientific basis. Primera edición. Editorial Chapman – Hall. Londres. Pág. 10 – 20, 214 - 222 GARCIA. R. H., 1995, Estudio del efecto de dos probióticos comerciales sobre la población intestinal de Escherichia coli en un lote de pollos de engorde., Universidad Nacional de Colombia, Facultad de Medicina Veterinaria y Zootecnia, Tesis de Pregrado. P. 8-24. GEDEK, B. R., 1999. Adherence of Escherichia coli serogroup 0 157 and the Salmonella typhimurium mutant DT 104 to the surface of Saccharomyces boulardii, Mycoses, Vol 42, P. 261-264. GHASEMI, H.A., TAHMASBI, A.M., MOGHADDAM, GH., MEHRI, M., ALIJANI, S., KASHEFI, E., FASIHI, A., 2006, The Effect of Phytase and Saccharomyces cerevisiae (Sc47) Supplementation on Performance, Serum Parameters, Phosphorous and Calcium Retention of Broiler Chickens., International Journal of Poultry Science, Vol 5, N 2, P. 162 - 168. GHERNA, R.L., 1995. Culture preservation, En: manual of methods for general bacteriology. American Society of Microbiology, P. 278 - 290. GIANNASCA, K. GIANNASCA, P. NEUTRA, M. 1996. Adherence of Salmonella typhimurium to Caco-2 Cells: Identification of a Glycoconjugate Receptor. Infection and Immunity. Vol 16, P. 135 – 145. 83 GILLILAND, S. E., STALEY, T. E., BUSH, L. J., 1984. Importance of bile tolerance of Lactobacillus acidophilus used as dietary adjunct. Journal of Dairy Science, Vol 67, P. 3045 3051. GILLILAND, S. E., and WALKER, D. K. 1990. Factors to consider when selecting a culture of Lactobacillus acidophilus as a dietary adjunct to produce a hypocholesterolemic effect in humans. Journal of Dairy Science. Vol 73, P. 905 - 911. GOLDIN, B. R., GORBACH, S.L., SAVELIN, M., BARAKAT, S., GUALTIERI, L. and SALMINEN, S., 1992. Survival of Lactobacillus species (strain GG) in human gastrointestinal tract. Digestive Diseases and Sciences, Vol 37, P. 121 - 128. GONZALEZ, A., VALENZUELA, L., 2000, Saccharomyces cerevisiae, En: Departamento de Genética Molecular, Instituto de Fisiología Celular, Universidad Nacional Autónoma de México. P. 1 – 5. GUSLANDI, M., GIOLLO, P., AND TESTONI, P., 2003. A pilot trial of Saccharomyces boulardii in ulcerative colitis. European Journal of Gastroenterology and Hepatology, Vol 15, P. 697 – 698. HAVENAAR, R., AND J. H. J. HUIS IN’T VELD, J. H. J., 1992. Probiotics: a general view. The lactic acid bacteria. En: Wood, B. J. B. (Ed). The lactic acid bacteria in health and disease. Elsevier Applied Science Vol 1, P. 151 - 171. HOLZAPFEL, W.H., HABERER, P., SNEL, J., SCHILLINGER, U., AND HUIS IN’T VELD, J.H. 1998. Overview of gut flora and probiotics. International Journal of Food Microbiology, Vol 41, P. 85 – 101. JACOBSEN, C. ROSENFELDT, V. HAYFORD, A. MOLLER, P. MICHAELSEN, K. PAERREGAARD, A SANDSTRO, B. TVEDE, M. JAKOBSEN, M. 1999. Screening of Probiotic Activities of Forty-Seven Strains of Lactobacillus spp. by In Vitro Techniques and Evaluation of the Colonization Ability of Five Selected Strains in Humans. Applied And Environmental Microbiology. Vol 16, P. 4949–4956 JONSSON, E. and CONWAY, P., 1992. Probiotics for pigs. Probióticos. The Scientific Basis. R. Fuller ed. Chapman and Hall editorial. Gran Bretaña. P. 268 . 84 JORDAN, F. T. W., PATTISON, M., 1998. Enfermedades de las aves. Tercera Edición, Editorial El Manual Moderno, México, P. 19. KHALIL, A.H.; MANSOUR, E.H. 1998. Alginate encapsulation Bifidobacteria survival in mayonnaise. Journal of Food Science, Vol 63, P. 702 – 705. KLIBS, N. G., SANTOS, J. E. P., COSCIONI, A., VILLASEÑOR, M., SISCHO, W. M., BERGE, A. C. B., 2005, Effect of feeding live yeast products to calves with failure of passive transfer on performance and patterns of antibiotic resistance in fecal Escherichia coli, Reprodution and Nutrition Development, Vol 45, P. 427 - 440. KOLLAR, R., REINHOLD, B. B., PETRAKOVA, E., YEH, H. J. C., ASHWELL, G., DRGONOVA, J., KAPTEYN, J. C., KLIS, F. M. AND CABIB, E. (1997). Architecture of the yeast cell wall: β (1,6)-glucan interconnects mannoproteins, β (1,3)-glucan and chitin. Journal of Biology and Chemistry. V 272, P. 17762 - 17775. KÚHLE, A.., SKOVGAARDB, K., JESPERSEN, L., 2005. In vitro screening of probiotic properties of Saccharomyces cerevisiae var. boulardii and food-borne Saccharomyces cerevisiae strains, International Journal of Food Microbiology Vol 101, P. 29 – 39. KUMPRECHTOVA, D., ZOBAC, P., KUMPRECHT, I., 2000, The effect of Saccharomyces cerevisiae SC47 on chicken broiler performance and nitrogen output. Czech Journal of Animal Science, Vol 45, P. 169 -177. KUMURA, H, TANOUE, Y, TSUKARA, M, TANAKA, T AND SHIMAZAKI, K, 2004. Screening of dairy yeast strains for probiotic applications, Journal of Dairy Science, Vol 87, P. 4050 – 4056. KUNG. L., KRECK, E., TUNG, R., HESSION, A., SHEPERD, A., COHEN, M., SWAIN, H. and LEEDLE, J. 1997. Effects of a live yeast culture and enzymes on in vitro ruminal fermentation and milk production of dairy cows, Journal of Dairy Science, Vol 80, N 9, P. 2045 - 2051. LESSON, T., LESSON, C., PAPARO, A., 1990. Texto/Atlas de Histología. Editorial Interamericana. México D.F., México, P. 422, 446. 85 LILLY, D., STILLWELL, R., 1965. Probioticis Growth promoting factors produced by microorganisms, Science, Vol 47, P.747 – 748. MACHADO-CAETANO, J.A., PARAMENS, M.T., BABO, M.J. ET AL. 1986. Immunopharmacological effects of Saccharomyces boulardii in healthy human volunteers. International Journal of Immunopharmacology, Vol 8, P.245 - 259. MADIGAN, M. T., MARTINKO, J. M., PARKER, J., 1998. Brock Biología de los microorganismos Octava Edición, Prentice Hall, España, P. 155 – 156. MAIJA, M. POKROTNIEKS, J. 2006. Probiotics as functional food: microbiological and medical aspects. Acta Universitatis Latviensis. Vol 710, P. 117–129 MANTILLA, J. SUAREZ, M. 2000. Presencia de Salmonella serovariedad enteritidis en productos de origen avícola y su repercusión en salud pública. Latreia 13: 237-245. MARIN, J., 2003. Evaluación de métodos de conservación para cepas de hongos filamentosos con actividad enzimática amilolítica. Tesis de Pregrado. Microbiología Industrial. Facultad de Ciencias. Pontificia Universidad Javeriana. P. 46-50 MARTINS, F. FERREIRA, F. PENNA, F. ROSA, C. DRUMMOND, R. NEVES, M. NICOL, J. 2005. Estudo do potencial probiótico de linhagens de saccharomyces cerevisiae através de testes in Vitro. Revista de Biologia e Ciências da Terra. Vol 5, P. 1 – 13. MASSOT, J., DESCONCLOIS, M. AND ASTOIN, J. 1982. Protection par Saccharomyces boulardii de la diarrheä ä Escherichia coli du souriceau. Annales Pharmaceutiques Francëaises, Vol 40, P. 445 - 449. MATHEWS, C., KENSAL, E., AHER, K. G., 2003. Bioquímica. 3º edición. Addison Wesley. Madrid, España, Págs. 100- 102 MATHEW, A., CHATTIN, S., ROBBINS, C., GOLDEN, D., 1998. Effects of a direct – fed yeast culture on enteric microbial populations, fermentation acids, and performance of wealing pigs., Journal of animal science, Vol 76, P. 2138 -2145. MCFARLAND, L., BERNASCONI, P. 1993. Saccharomyces boulardii: a review of an innovative biotherapeutic agent. Microbiology and Ecology Health Disease, Vol 6, P. 57 – 171 86 MCFARLAND, L., SUWAWICZ, C., GREENBERG, R.,1994. A randomized placebocontrolled trial of Saccharomyces boulardii in combination with standard antibiotics for Clostridium difficile disease. Journal of American Medical Association, Vol 271, P. 1913 1918 MEDINA, B, GIRARD, D., JAVOTOT, E., JULLIAND, V., 2002. Effect of a preparation of Saccharomyces cerevisiae on microbial and fermentation patterns in the large intestine of horses fed a high fiber or a high starch diet, Journal of Animal Science, Vol 80, P. 2600 2609. METCHNIKOFF, E. 1907. Lactic acid as inhibiting intestinal putrefaction. En: The Prolongation of Life: Optimistic Studies. HEINEMANN, W. (Ed), London, 1907, P. 161 - 183. MEZA, R. MONROY, A. MERCADO, M. POUTOU, R. RODRÍGUEZ, P. PEDROZA, A. 2004. Study of the stability in real time of cryopreserved strain banks. Universitas Scientiarum. Vol 9, P. 25 - 42. MILLAR, M. R., BACON, C., SMITH, S. I., WALKER, V. and HALL, M.A. 1993. Enteral feeding of premature infants with Lactobacillus GG. Archives of Disease in Childhood, Vol 69, P.483 - 487. MITSUI, K.YASUI, H. NAKAMURA, N. KANAZAWA, H. 2005. Oligomerization of the Saccharomyces cerevisiae Na+/H+ antiporter Nha1p: Implications for its antiporter activity. Biochimica et Biophysica Acta Vol 1720, P. 125 - 136. MOLINA, L, 2002. La avicultura en Colombia, Editado por FENAVI, Bogotá, P. 24- 26. MOSER, S. A., and SAVAGE, D. C., 2001. Bile salt hydrolase activity and resistance to toxicity of conjugated bile salts are unrelated properties in lactobacilli. Applied Environmental Microbiology. Vol 67, P. 3476 - 3480. MURAKAMI, A., V. M. BARBOSA, J. ARIKI, O.M. JUNQUEIRA, Y S. KRONKA. 1993. Levedura de vinhaca (Saccharomyces cereviciae) como fonte proteica na alimentação de frangos de corte. Revista of Society of Brasil. Vol 22, N 5, P. 876 -883. 87 NARENDRANATH, V., THOMAS, K., INGLEDEW, W. 2001. Effects of acetic acid and lactic acid on the growth of Saccharomyces cerevisiae in a minimal medium. Journal of Industrial Microbiology and Biotechnology, Vol 3, P. 171 – 177. NETHERWOOD, T., GILBERT, H., PARKER, S., O’DONNELL, G., 1999, Probiotics shown to change bacterial community structure in the avian gastrointestinal tract, Applied and Environmental Microbiology. P. 5134 - 5138. NEWBOLD, C.J., WALLACE, R.J., CHEN, X.B. AND MCINTOSH, F.M. 1995. Different strains of Saccharomyces cerevisiae differ in their effects on ruminal bacterial numbers in vitro and in sheep. Journal of Animal Science, Vol 73, P. 1811 - 1818. OHGAKI, R. NAKAMURA, N. MITSUI, K. KANAZAWA, H. 2005. Characterization of the ion transport activity of the budding yeast Na+/H+ antiporter, Nha1p, using isolated secretory vesicles. Biochimica et Biophysica Acta. 1712:185 – 196 ORTIZ, D., St.PIERRE, M., ABDULMESSIH, A., ARIAS, I., 1997. A yeast ATP-binding cassette-type protein medianting ATP-dependent bile acid transport. The Journal of Biological Chemistry. Vol 272, N 24, P. 15358 - 15365. OLIVEIRA, G., BERCHIERI, A., BARROW, P., 2000, Prevention of salmonella infection by contact using intestinal flora of adult birds and/or a mixture of organic acids, Brazilian Journal of Microbiology, Vol 31, P. 116 – 120. OTERO, G., FORERO, M., 1997. Evaluación del uso de dos probióticos comerciales en lechones lactantes. .Universidad Nacional de Colombia, Facultad de Medicina Veterinaria y Zootecnia, Tesis de Pregrado, P. 14. OUWEHAND, A.C. AND P.L. CONWAY 1996. Purification and characterization of a component produced by Lactobacillus fermentum that inhibits the adhesion of K88 expressing Escherichia coli to porcine ileal mucus. Journal of. Applied. Bacteriology., Vol 80, P. :311-318. OYETAYO, V.O., and OYETAYO, F.L., 2005. Potential of probiotics as biotherapeutic agents targeting the innate immune system. African Journal of Biotechnology, Vol 4, N 2, P. 123 – 127. 88 PARDIO, V., WALISZESWSKI, K. y ROBLEDO G., 1996. Los probióticos y su futuro. Archivo Latinoamérica de nutrición. Vol 46, N 1, P. 6 - 10. PARK, J.K.; CHANG, H.N. 2000. Microencapsulation of microbiall cells. Biotechnology. Advances, Vol 18, P. 303-319. PARKER, R. B. 1974. Probiotics, the other half of the antibiotic story. Animal Nutrition Health, Vol 29, P. 4 - 8. PELIZON, A. C., KANENO, R., SOARES, A. M., MEIRA, D. A., SARTORI, A.,2003. Downmodulation of lymphoproliferation and interferon-gamma production by beta-glucan derived from Saccharomyces cerevisiae. Memorial Institute Oswaldo Cruz, Vol 98, N 8, P. 1083 1087 PEÑA, M. J., 2000, Efecto de la adición de microorganismos al agua de bebida y en la camada, como fuentes de probiótico de pollos parrilleros. University of Puerto Rico. Tesis Maestría, Puerto Rico. P. 2-3.. PERDOMO, M. C., VARGAS, R. E., CAMPOS, J., 2004, Valor nutritivo de la levadura de cervecería (Saccharomyces cerevisiae) y de sus derivados, extracto y pared celular, en la alimentación aviar., Archivos Latinoamericanos de Producción Animal, Vol 12, N 3, P. 89 – 95. PEREZ, L. S., TALAVERA, M., LAGUNAS, S., CUARON, J., VAZQUEZ, J., 2005, In vitro evaluation of the binding capacity of Saccharomyces cerevisiae Sc47 to adhere to the wall of Salmonella spp. Revista Latinoamericana de Microbiología, Vol. 47, No. 3-4, P. 70 - 75 PSOMAS, E. FLETOURIS, D. LITOPOULOU, E. TZANETAKIS, N. 2003. Assimilation of Cholesterol by Yeast Strains Isolated from Infant Feces and Feta Cheese. Journal of Dairy Science, Vol 86, P. 3416 - 3422. RÉ, M., 1998, Microencapsulation in spray drying, En. Institute for Technology Research – Chemistry division, Sao Paulo, Brazil, Pág 1206. RODRIGUES, A. P., NARDI, R.M., BAMBIRRA, E.A., VIEIRA, E.C. AND NICOLI, J.R. 1996. Effect of Saccharomyces boulardii against experimental oral infection with Salmonella 89 typhimurium and Shigella flexneri in conventional and gnotobiotic mice. Journal of Applied Bacteriology, Vol 81, P. 251 - 256. ROSE, A. H., 1987. Responses to the chemical environment. En: Rose, A. J., The yeast V2, yeast and the environment. Academic Press. Londres. P. 5. ROSE, S. P., 1997 Principios De La Ciencia Avícola, Acribia, Zaragoza, España, P. 113 – 117. RUIZ, C., VICTOR, J.,, LUSSIER, M., MOLINA, M., and NOMBELA, C., 1999. A Large-scale Sonication Assay for Cell Wall Mutant Analysis in Yeast, Yeast, V 15, P. 1001 - 1008 SALMINEN, S., OUWENHAND, A., LEE, Y. 1999. Probiotics: How should they be defined? Trends of Food Science and Technology Vol 10, P.107 - 110. SAMBUY, Y., DE ANGELIS, I., RANALDI, G., SCARINO, M. L., STAMMATI, A., ZUCCO. F., 2005. The Caco-2 cell line as a model of the intestinal barrier: influence of cell and culturerelated factors on Caco- cell functional characteristics. Cell biology and toxicology. Vol 21, P. 1 -26. SÁNCHEZ, L., CORRALES, L. 2005. Evaluación de la congelación para conservación de especies autóctonas bacterianas. Nova. Vol 3, P. 21 - 29. SARMIENTO, A., HERRERA. J., 2002. Obtención y caracterización de un banco de levaduras nativas con potencial aplicación microbiológica. Tesis de Pregrado. Facultad de Ciencias. Pontificia Universidad Javeriana. P. 17 – 20, 29 – 32, 65 -66. SCHREZENMEIER, J., DE VRESE, M. 2001. Probiotics, prebiotics and symbiotics approching a definition. American Journal of Clinical Nutrition, Vol 73, P. 361 - 364. 65 SHEU, T.Y.; MARSHALL, R.T.; HEYMANN, H. 1993. Improving survival of culture bacteria in frozen desserts by microentrapment. Journal of Dairy Science, Vol 76, P. 1902 – 1907. SKOOG, D., 1988. Fundamentos de química analítica, Segunda edición, Editorial Reverté S.A. Barcelona, España, P. 10 – 15, 20 – 24. 90 SMIMOV, A., PEREZ, R., AMIT-ROMACH, E., SKLAN, D., UNI, Z., 2005. Mucin dynamics and microbial populations in chicken small intestine are changed by dietary probiotic and antibiotic growth promoter supplementation, Journal of Nutrition, Vol 135, P. 187 - 192. SPENCER, J.F. RAGOUT, A.L. 2001. Métodos microbiológicos Humana Press Inc. Totowa. New Jersey, P. 173 -181 SPERTI, G. S., 1971. Probiotics, Avi Publishing Co., West Point, Connecticut. St PIERRE, M., RUETZ, S., EPSTEIN, L., GROS, P., ARIAS, I., 1994. ATP-dependent transport of organic anions in secretory vesicles of Saccharomyces cerevisiae. Proceedings of Natural Academic Science, Vol 91, P. 9476 - 9479. STURKIE, J. E., V. M. MCDONOUGH, AND C. E. MARTIN. 1989. Isolation and characterization of OLE1, a gene affecting fatty acid desaturation from Saccharomyces 5 cerevisiae. Journal of Biology and Chemistry, Vol 264, P. 16537 - 16544. SURAWICZ, C.M., MCFARLAND, L.V., ELMER, G., CHINN, J., 1989. Treatment of recurrent Clostridium difficile colitis with vancomycin and Saccharomyces boulardii. The American Journal of Gastroenterology Vol 84, P. 1285– 1287. SYCHROVAÈ, H. RAMIREZ, J. PEÑA, A. 1999. Involvement of Nha1 antiporter in regulation of intracellular pH in Saccharomyces cerevisiae. Microbiology Letters, Vol 171, P.167 - 172. TARANTO, M. P., MEDICI, M., PERDIGON, G., RUIZ HOLGADO, A. P.and VALDEZ, G. F., 1998. Evidence for hypocholesterolemic effect of Lactobacillus reuteri in hypercholesterolemic mice. Journal of Dairy Science, Vol 81, P. 2336 - 2340. TAYLOR, F. R., and PARKS., L.W., 1981. An assessment of the especifity of sterol uptake and sterification in Saccharomyces cerevisiae. Journal of Biology and Chemical, Vol 256, P. 981 – 985. THOMAS, K. C., HYNES, S. H., Y INGLEDEW, M. W., 2002. Influence of Medium Buffering Capacity on Inhibition of Saccharomyces cerevisiae Growth by Acetic and Lactic Acids. Environmental Microbiology, Vol 68, N. 4, P. 1616 - 1623. 91 TISSIER, H., 1906. Traitement des infections intestinales par la méthode de transformation de la flore bactérienne de l'intestin. Comptes Rendus des Séances et Mémoires de la Société de Biologie, Vol 60, P. 359 - 361. TODOKIRI, K., MUKAI, T., SATO, S. AND TOBA, T. 2001 Inhibition of adhesion of foodborne pathogens to Caco-2 cells by Lactobacillus strains, Journal of Applied Microbiology, Vol 91, P. 154 – 159. TOMA, M. M., RAIPULIS, J., KALNINA, I., RUTKI, R., 2005. Does probiotic yeast act as antigenotoxin?. Food Technology and Biotechnology. Vol 43, N 3, P. 301–305. TORTUERO, F. 1973. Influence of the implantation of Lactobacillus acidophilus in chicks on growth, feed conversion, malabsorption of fat syndrome and in-testinal flora. Poultry Science, Vol 52, P. 197-202. TUITÉ, M., 1991. Saccharomyces, En: Biotechnology Handbooks N 4. Ed. Plenum Press. New York P. 10 – 23. VALERIO, F. DE BELLIS, P.LONIGRO, S. MORELLI, L.VISCONTI, A. LAVERMICOCCAIN, P. 2006. In vitro and in vivo survival and transit tolerance of potentially probiotic strains carried by artichokes in the gastrointestinal tract. Applied and Environmental Microbiology. Vol 72, P. 3042 - 3045. VERSTREPEN, K. KLIS, F. 2006. Flocculation, adhesion and biofilm formation in yeasts. Molecular Microbiology. Vol 60, P. 5–15 VIEGAS, C. ALMEIDA, P. CAVACO, M. CORREIA. I. 1998. The H1-ATPase in the plasma membrane of Saccharomyces cerevisiae is activated during growth latency in Octanoic Acidsupplemented medium accompanying the decrease in intracellular pH and cell viability. Applied and Environmental Microbiology. Vol 64, P. 779 - 783. WARLIES, F. AHRENS, and HOLZAPFEL, W. H., 1998. Characterisation and selection of probiotic lactobacilli for a preliminary minipig feeding trial and their effect on serum cholesterol levels, faeces pH and faeces moisture content. International Journal of Food Microbiology. Vol 40, P. 93 - 104. 92 WILLICOCKS, S., 1990, Growth and characterisation of human faecal Astrovirus in a continuos cell line. Archives of Virology, Vol 113, P. 78 - 81. WOERNLE, H.,1996, Enfermedades de las aves, Acribia, Zaragoza, España, P. 62 - 67. ZWEIBAUM, A., LABURTHE, M., GRASSET, E. y LOUVARD, D., 1991. Use of cultured cell lines in studies of intestinal cell differentiation and function. Handbook of Physiology. The Gastroenterology System. Vol 4, P. 223 - 225. Recursos electrónicos BD Worldwide. (2007: USA). BioCoat™ HTS Caco-2 Assay System [en línea]. <http://www.bdbiosciences.com/discovery_labware/Products/inserts/BD_Biocoat_insert_syst ems/caco-2_assay_system/hints.shtml > [Consulta: 14 Jun 2007] University of Bath (2002: Reino Unido) Yeast genetics and genomics [en línea]. Department of Biology and Biochemistry. <http://www.bath.ac.uk/bio-sci/wheals2.htm> [Consulta: 14 Jun 2007] FENAVI (2006: Colombia). Avicultura creció 10% un semestre fuera de lo presupuestado Boletín N 10, Julio, Pág 10- 14 [en línea], <http:www.fenavi.org.co/> [Consulta: 14 de Jun 2007] Revista electrónica de Veterinaria REDVET (2005: Cuba). SOLANO, G., SALCEDO, M. L., RAMIREZ, R., 2005. Dietas para pollos en ceba a base de subproductos de la agroindustria local. REDVET, Vol, 6, N 2, [en línea], <http:www.veterinaria.org/revistas/redvet> [Consulta: 14 de Jun 2007] 93 10. ANEXOS ANEXO Nº 1. ASPECTO MACROSCÓPICO DE S. cerevisiae y S. cerevisiae var. boulardii • Anexo 1.1. Figura Nº 1.1. Aspecto macrocópico de S. cerevisiae var. boulardii en Agar YPG • Anexo 1.2. Figura Nº 1.2. Aspecto macrocópico de S. cerevisiae en Agar YPG 94 • Anexo 1.3. Figura Nº 1.3. Recuento en microgota de S. cerevisiae en Agar YPG ANEXO Nº 2. TABLAS DE RESULTADOS OBTENIDOS PARA CONTROL DE CALIDAD DE BANCOS DE CEPA A Y B • Anexo 2.1. Tabla Nº 2A Coloración de Gram de inóculos y colonias de cepa A (estudio) y cepa B (control) Cepa A Gram Gram inóculos colonias Levaduras Levaduras sin sin contaminación contaminación B Levaduras Levaduras sin sin contaminación contaminación 95 • Anexo 2.2. Tabla Nº 2B Técnica: Siembra en superficie en agar YPG de control de calidad de Cepa A (estudio) y Cepa B (control) Mes Cepa A Cepa B 1 5,6E+08 4,5E+08 2 9,0E+08 9,8E+08 3 6,5E+07 6,0E+08 4 5,0E+05 4,0E+08 5 1,7E+08 1,8E+08 ANEXO Nº 3. TABLA RESULTADOS DE PRUEBA DE TOLERANCIA A SALES BILIARES Tabla Nº 3. Técnica: Siembra en microgota en agar YPG suplementado con sales biliares para cepa A (estudio) y cepa B (control) Concentración Cepa A Cepa B (% p/v) UFC/mL UFC/mL 0.05 % 6,60E+06 7,90E+06 0.10 % 7,30E+06 7,90E+06 0.15 % 6,80E+06 7,50E+06 0.20 % 5,00E+06 8,80E+06 0.25 % 8,20E+06 8,00E+06 0.30 % 5,50E+06 5,00E+06 Sales Biliares 96 ANEXO Nº 4. TABLA RESULTADOS DE PRUEBA DE TOLERANCIA A RANGOS DE pH Tabla Nº 4 Técnica: Siembra en microgota en agar YPG de pH para cepa A (estudio) y cepa B (control) Valor Cepa A Cepa B pH UFC/mL UFC/mL 2.0 3,20E+06 5,80E+05 2,5 2,20E+06 2,60E+05 3.0 2,90E+06 3,30E+05 3,5 3,60E+06 4,80E+05 4.0 6,30E+06 1,50E+05 4,5 3,30E+06 7,00E+06 5.0 8,80E+06 6,50E+07 ANEXO Nº 5. TABLA RESULTADOS DE PRUEBA DE RESISTENCIA A JUGOS GÁSTRICOS Tabla Nº 5. Técnica: Siembra en microgota en agar YPG de jugos gástricos para cepa A (estudio) y cepa B (control) CEPA A CEPA B MUESTREO Jugo Gástrico pH 2.0 – 2.3 Jugo Gástrico Jugo Gástrico pH 7.0 – 7.2 pH 2.0 – 2.3 Jugo Gástrico pH 7.0 – 7.2 Horas UFC/mL UFC/mL UFC/mL UFC/mL 0 6,40E+05 6,00E+05 7,40E+05 8,80E+05 1 5,30E+05 7,00E+05 8,50E+05 7,80E+05 2 6,00E+05 7,30E+05 9,20E+05 9,30E+05 3 6,30E+05 5,30E+05 7,30E+05 6,50E+05 4 7,30E+05 7,40E+05 6,80E+05 8,00E+05 24 9,50E+05 6,90E+05 5,00E+05 9,50E+05 97 ANEXO Nº 6. TABLAS Y FIGURAS DE PRUEBA DE DETERMINACIÓN DE REDUCCION DE COLESTEROL EN PRESENCIA DE SALES BILIARES Anexo 6.1. Protocolo de elaboración de curva patrón de colesterol - Se parte del reactivo Lipids Colesterol Rich (Sigma ®) con una concentración inicial de colesterol de 9 – 11 g/L y se realiza la conversión a microgramos/mililitro, es decir la concentración inicial del reactivo es 9000 a 11000 μg/mL - Se toma como concentración inicial el promedio, es decir 10000 μg/mL. - Se aplica la fórmula V1 * C1 = V2 * C2 Donde: V1= Volumen inicial de Lipids Colesterol Rich (mL) C1= Concentración inicial (10000 μg/ml) V2= 20 mL de caldo YPG C1= Diferentes concentraciones conocidas de colesterol (60μg/mL – 130 μg/mL) Anexo 6.2. Curva patrón Tabla Nº 6.2. Valores de Absorbancia (550 nm) obtenidos en la elaboración de la réplicas de la curva patrón para la determinación de colesterol Absorbancia (550 nm) Absorbancia (550 nm) Absorbancia (550 nm) colesterol (μg/mL) Réplica 1 Réplica 2 Promedio 60 0.123 0.101 0.112 70 0.178 0.145 0.1615 80 0.206 0.204 0.205 90 0.266 0.233 0.2495 100 0.271 0.301 0.286 110 0.366 0.347 0.3565 120 0.387 0.398 0.3925 130 0.416 0.422 0.419 Concentración 98 Anexo 6.3. Tabla Nº 6.3. Valores de Absorbancia (550 nm) obtenidos en la determinación de reducción de colesterol del medio YPGCHO por las cepa A y Cepa B Cepa A Cepa B Réplicas Absorbancia (550 nm) Absorbancia (550 nm) 1 0.319 0.258 2 0.308 0.261 3 0.282 0.277 Promedio 0.303 0.265 Medio YPGCHO Anexo 6.4. Figura Nº 6.4. Medio YPGCHO inoculado con la cepa A (estudio) después de 12 horas de incubación 99 Anexo 6.5. Figura Nº 6.5. Medio YPGCHO inoculado con la cepa B (control) después de 12 Horas de incubación Anexo 6.6. Figura Nº 6.6. Separación de fases orgánica y acuosa en la extracción de colesterol de las muestras (cepa A y B) 100 Anexo 6.7. Figura Nº 6.7. Reaccion de Liebermann - Burchard para la determinación de colesterol en medio YPGCHO (cepa A) Anexo 6.8. Figura Nº 6.8. Reaccion de Liebermann - Burchard para la determinación de colesterol en medio YPGCHO (cepa B) 101 ANEXO 7. FIGURAS DE ENSAYO DE ADHESION A CÉLULAS Caco – 2 Anexo 7.1. Figura 7.1. Análisis de adherencia por microscopio invertido de Saccharomyces cerevisiae a células Caco-2 con colorante de Wright. Anexo 7.2. Figura 7.2. Análisis de adherencia por microscopio invertido de Saccharomyces cerevisiae var. boulardii a células Caco-2 con colorante de Wright. 102 ANEXO 8. COMPOSICION DE MEDIOS DE CULTIVO Y REACTIVOS Anexo 8.1. Agar YPG TABLA Nº 6.1, Composición (g/L) de Medio YPG Componente Gramos/Litro Peptona 20 Glucosa 20 Extracto de levadura 10 Agar 15 Preparación: Se disuelven los componentes en un litro de agua desmineralizada por calentamiento. Tratar en autoclave (15 minutos, 120oC, 15 libras de presión). pH 6.6 2 a 25 oC. Anexo 8.2. Caldo YGC TABLA Nº 6.2, Composición (g/L) de Caldo YGC Componente Gramos/Litro Peptona 20 Glucosa 20 Extracto de levadura 10 Cloramfenicol 0.1 Preparación: Se disuelven los componentes en un litro de agua desmineralizada por calentamiento. Tratar en autoclave (15 minutos, 120oC, 15 libras de presión). pH 6.6 ± 2 a 25 oC. 103 Anexo 8.3. Caldo YPGCHO TABLA Nº 6.3. Composición (g/L) de Caldo YPGCHO Componente Gramos/Litro Peptona 20 Glucosa 20 Extracto de levadura 10 Bile Oxgall (Difco ® ) 0.1 Lipids Colesterol Rich (Sigma ®) Según uso Preparación: Se disuelven la peptona, glucosa, extracto de levadura y las sales biliares (Bile Oxgall) en un litro de agua desmineralizada por calentamiento. Tratar en autoclave (15 minutos, 120oC, 15 libras de presión). En el momento del uso suplementar con el reactivo Lipids Colesterol Rich (Sigma ®), según uso para elaborar curva patrón o determinación de reducción de colesterol por microorganismos probióticos. Anexo 8.4. Reactivo de extracción de colesterol. • Etanol 95 % (v/v): preparado al momento de usar o refrigerado a 4 ºC. • Hidróxido de Potasio 50% (p/v): preparado al momento de usar o con antelación dejando en temperatura ambiente. • Hexano: debe ser puro y dejarse refrigerado a 4 ºC. • O – phthaldehído (Sigma ®): este reactivo debe mantenerse refrigerado y al momento de usar, se pesan 0.5 mg y se disuelven en 1 mL de Ácido Glacial Acético. Anexo 8.5. Medio Mínimo Esencial Eagle El medio debe suplementarse con una solución que contiene 0.6436 gramos de Glutamina (4.4 mM), 0.006 gramos de Penicilina y 100 mg de Estreptomicina llevados a 10 mL de agua destilada y posteriormente debe esterilizarse con filtros de membrana de 0.22 μm. Esta solución se adiciona a 1 Litro de Medio Mínimo Esencial Eagle. Adicionalmente, se debe 104 inactivar el Suero Fetal Bovino (30 minutos a 57ºC en baño termostatado) y filtrar de la manera descrita anteriormente. Se debe adicional 100 mL de SFB por cada Litro de Medio de Cultivo. Anexo 8.6. PBS Se deben pesar 8 gramos de NaCl, 0.2 gramos de KCl, 0.51 gramos de KH2PO4 y 1.6 gramos de Na2HPO4 y llevarlos a 1000 mL con agua destilada, ajustando el pH con HCl 1 N a 7.1 – 7.2 y posteriormente esterilizar en autoclave 121ºC durante 20 minutos y almacenar a 4ºC. ANEXO 9. PROTOCOLO DE TRIPSINIZACION DE CÉLULAS Caco-2. - Se toma una caja con células confluentes crecidas en medio mínimo esencial Eagle con 15% se suero fetal bovino. - Se retira el medio mínimo esencial - Las células son lavadas con 3 mL de solución de PBS la cual es posteriormente retirada. - Se adiciona 0.5 mL de Tripsina a 37oC para humedecer la monocapa y subsecuentemente desprender las células. - Una vez las células se vean individuales, se golpea suavemente el frasco para desprender las células del mismo. - A continuación las células son resuspendidas en 5 mL de medio mínimo esencial con suero fetal bovino al 15%. ANEXO 10. TINCION DE WRIGHT Se pesan 0.5 g de bicarbonato sódico y 1 g de azul de metileno en 100 ml de agua destilada. La mezcla se calienta a 100º C durante 1 hora. Se enfría y se añade una solución de eosina al 1%. El precipitado se recoge en un filtro, se seca y disuelve en alcohol metílico en la proporción de 0.5 g/litro de alcohol. ANEXO 11. ANÁLISIS ESTADÍSTICO Para las pruebas llevadas a cabo en este estudio, se realizó una Prueba t - student. Teniendo en cuenta que se plantean hipótesis donde: Ho: Hipótesis nula Hi: Hipótesis alterna 105 Y se tiene que: Si P < 0.05 Se rechaza Ho Si P > 0.05 Se acepta Ho Anexo 11.1. Prueba Tolerancia a Sales Biliares. Ho: La tolerancia a las concentraciones de sales biliares evaluadas (0.05% - 0.3%) por parte de la cepa A (estudio) es igual a la presentada por la cepa B (control). Hi: La tolerancia a las concentraciones de sales biliares evaluadas (0.05% - 0.3%) por parte de la cepa A (estudio) es no es igual a la presentada por la cepa B (control). P = 0.214 Por lo tanto se acepta la hipótesis nula y se concluye que no existe evidencia estadísticamente significativa para afirmar que hay diferencia entre las medias de los recuentos (UFC/mL) de ambas cepas. Anexo 11.2. Prueba Tolerancia a pH. Ho: La tolerancia a los diferentes valores de pH (2.0 – 5.0) por parte de la cepa A (estudio) es igual a la presentada por la cepa B (control). Hi: La tolerancia a los diferentes valores de pH (2.0 – 5.0) por parte de la cepa A (estudio) no es igual a la presentada por la cepa B (control). P = 0.523 Por lo tanto se acepta la hipótesis nula y se concluye que no existe evidencia estadísticamente significativa para afirmar que hay diferencia entre las medias de los recuentos (UFC/mL) de ambas cepas. Anexo 11.3. Prueba Tolerancia a Jugos gástricos. Jugo Gástrico pH 2.0 – 2.3 Ho: La tolerancia al jugo gástrico artificial (pH 2.0 – 2.3) por parte de la cepa A (estudio) es igual a la presentada por la cepa B (control). 106 Hi: La tolerancia al jugo gástrico artificial (pH 2.0 – 2.3) por parte de la cepa A (estudio) no es igual a la presentada por la cepa B (control). P = 0.517 Por lo tanto se acepta la hipótesis nula y se concluye que no existe evidencia estadísticamente significativa para afirmar que hay diferencia entre las medias de los recuentos (UFC/mL) de ambas cepas. Jugo Gástrico pH 7.0 – 7.2 Ho: La tolerancia al jugo gástrico artificial (pH 7.0 – 7.2) por parte de la cepa A (estudio) es igual a la presentada por la cepa B (control). Hi: La tolerancia al jugo gástrico artificial (pH 7.0 – 7.2) por parte de la cepa A (estudio) no es igual a la presentada por la cepa B (control). P = 0.833 Por lo tanto se acepta la hipótesis nula y se concluye que no existe evidencia estadísticamente significativa para afirmar que hay diferencia entre las medias de los recuentos (UFC/mL) de ambas cepas. Anexo 11.4. Prueba reducción de colesterol en presencia de sales biliares. Ho: La reducción de colesterol en presencia de sales biliares por parte de la cepa A (estudio) es igual a la presentada por la cepa B (control). Hi: La reducción de colesterol en presencia de sales biliares por parte de la cepa A (estudio) no es igual a la presentada por la cepa B (control). P = 0.154 Por lo tanto se acepta la hipótesis nula y se concluye que no existe evidencia estadísticamente significativa para afirmar que hay diferencia entre las medias de los valores de absorbancia 550 nm medidos para ambas cepas. 107