BIBLIO Instituto Salesiano de Estudios Superiores Compilación y armado Sergio Pellizza biblioises Apoyatura Académica Bibliotecas digitales para todos Zoología Introducción 1 Resúmenes del Hickman: Principios Integrales de Zoología (13ª Ed.) Capitulo 1: La vida: los principios biológicos y la ciencia zoológica (pg. 21) La zoología es el estudio científico de los animales y forma parte de la biología, el estudio científico de la vida. Los animales y la vida, en general, son entidades constantemente cambiantes que solo se pueden identificar en términos de los atributos que han adquirido a lo largo de su larga historia evolutiva. Las propiedades más importantes de la vida son: la exclusividad química, la complejidad y la organización jerárquica, la reproducción, la posesión de un programa genético, el metabolismo, el desarrollo y la interacción con el medio. Los sistemas biológicos constituyen una jerarquía de niveles integrados (molecular, celular, del organismo, poblacional y de especie), cada uno de los cuales presenta diversas propiedades emergentes especificas. La ciencia se caracteriza por la adquisición de conocimientos mediante la construcción y comprobación de hipótesis a través de la observación de la naturaleza. La ciencia se guía por la ley natural y sus hipótesis son comparables, provisionales y refutables. Las ciencias zoológicas se pueden subdividir en dos categorías, las ciencias experimentales y las evolutivas. Las ciencias experimentales utilizan el método experimental para preguntarse como los animales llevan a cabo sus funciones metabólicas básicas, del desarrollo, de la conducta y reproductoras, lo que incluye investigaciones sobre sus sistemas moleculares, celulares y poblacionales. Las ciencias evolutivas usan el método comparativo para reconstruir la historia de la vida y, posteriormente, valerse de esta historia con el fin de comprender como las diversas especies y sus propiedades moleculares, celulares, del organismo y de las poblaciones que han aparecido a lo largo de la evolución. Las hipótesis que resisten repetidas pruebas y además explican fenómenos muy diversos alcanzan la categoría de teorías. Las teorías poderosas que son guía de investigaciones extensas se denominan “paradigmas”. Los principales paradigmas que gobiernan el estudio de la zoología son la teoría de la evolución de Darwin y la teoría cromosómica de la herencia. Los principios que aparecen en este capítulo ilustran la unidad de las ciencias biológicas. Todos los componentes de los sistemas biológicos están guiados y limitados por leyes naturales. Los organismos vivos proceden exclusivamente de otros organismos vivos, de la misma forma que toda célula proviene de una célula preexistente. Los procesos de reproducción están presentes en todos los niveles de la jerarquía biológica y muestran tanto herencia como variación. La interacción de la herencia y la 1 variación en todos los niveles jerárquicos produce cambios evolutivos y ha generado la enorme diversidad de la vida animal que queda patente a lo largo de este libro. Capítulo 6: Evolución orgánica (pg. 147) La evolución orgánica explica la diversidad de los organismos vivientes como el resultado histórico del cambio gradual a partir de formas preexistentes. La teoría de la evolución está estrechamente ligada a Charles Robert Darwin, quien presento la primera explicación plausible del cambio evolutivo. Darwin extrajo la mayor parte del material que utilizo para construir su teoría de sus experiencias en un viaje de cinco años alrededor del mundo a bordo del HMS Beagle. La teoría evolutiva de Darwin tiene cinco componentes principales. Su proposición más básica es el cambio perpetuo, la teoría de que el mundo no es ni constante ni se halla en un ciclo perpetuo, sino que sufre continuamente cambios irreversibles. El registro fósil demuestra claramente el cambio perpetuo en las fluctuaciones continuas de la forma y la diversidad animales que siguieron a la explosión cámbrica de hace 600 millones de años. La teoría de Darwin del origen común establece que todos los organismos descienden de un antecesor común a través de la ramificación de líneas genealógicas. Esta teoría explica las homologías morfológicas entre los organismos como caracteres heredados con modificaciones a partir de los rasgos correspondientes en su antecesor evolutivo común. Los patrones de homologías formados por el origen común con modificaciones nos permiten clasificar los organismos de acuerdo con sus relaciones evolutivas. Un corolario del origen común es la multiplicación de las especies a lo largo del tiempo evolutivo. La especiación alopátrida describe la evolución de barreras reproductoras entre poblaciones separadas geográficamente, lo que origina nuevas especies. En algunos animales, especialmente insectos parásitos especializados en distintas especies hospedadoras, la especiación puede producirse sin aislamiento geográfico, lo que se conoce como especiación simpátrida. La radiación adaptativa es la proliferación de varias especies diversamente adaptadas a partir de una especie ancestral. Los archipiélagos oceánicos, como las islas Galápagos, son particularmente idóneos para la radiación adaptativa de organismos terrestres. La teoría darwiniana del gradualismo establece que las grandes diferencias fenotípicas entre las especies se producen por la acumulación de muchos pequeños cambios individuales a lo largo del tiempo evolutivo. El gradualismo es todavía objeto de controversia. Las mutaciones con grandes efectos sobre el fenotipo se han utilizado ampliamente en la cría de animales, lo que ha llevado a algunos a discutir el criterio de Darwin de que tales mutaciones no son importantes en la evolución. Desde una perspectiva macroevolutiva, el equilibrio puntuado establece que la mayor parte de los cambios evolutivos tienen lugar en procesos relativamente cortos de especiación ramificada, separados por largos intervalos en los que se van acumulando pequeños cambios fenotípicos. El quinto principio fundamental de Darwin es que la selección natural es el impulso que dirige la evolución. Este principio se basa en el hecho observado de que todas las especies presentan superproducción de sus individuos, lo que produce una competición por los recursos limitados que mantienen la vida. Como no hay dos organismos exactamente iguales, y ya que los rasgos variables son parcialmente heredables, aquellos cuyo bagaje hereditario mejore su utilización 2 de los recursos para sobrevivir y reproducirse, contribuirán desproporcionadamente a la siguiente generación. A lo largo de muchas generaciones, la selección de estas variaciones produce nuevas especies y nuevas adaptaciones. Las mutaciones son la fuente última de toda nueva variación sobre la que actúa la selección natural. La teoría de Darwin subraya que la variación se produce al azar con respecto de las necesidades del organismo, y que la supervivencia y la reproducción diferenciales proporcionan la dirección del cambio evolutivo. La teoría de Darwin de la selección natural fue modificada en este siglo mediante la corrección de sus errores genéticos. Esta teoría modificada se conoce como Neodarwinismo. Los genetistas de poblaciones descubrieron los principios por los cuales las propiedades genéticas de las poblaciones cambian a lo largo del tiempo. Un descubrimiento particularmente importante, conocido como el equilibrio de Hardy-Weinberg, demostró que el proceso hereditario no cambia por si mismo la composición genética de las poblaciones. Las fuentes más importantes de cambio evolutivo son la mutación, la deriva genética, los cruzamientos al azar, la migración, la selección natural y sus interacciones respectivas. El Neodarwinismo, tal y como lo formula la genética de poblaciones, constituyo la base de la Síntesis Evolutiva de los años 30 y 40. La genética, la historia natural, la paleobiología y la sistemática se reunieron bajo el objetivo común de expandir el conocimiento de la evolución darwiniana. La microevolución comprende el estudio del cambio genético en las poblaciones contemporáneas. Estos estudios muestran que la mayoría de las poblaciones naturales contienen enormes cantidades de variación. La macroevolución es el estudio del cambio evolutivo en una escala geológica de tiempo. Los estudios macroevolutivos miden tasas de especiación, extinción y cambios en la diversidad a través del tiempo. Estos estudios han extendido la teoría evolutiva darwinista hasta procesos de nivel superior, que regulan tasas de especiación, extinción y cambios en la diversidad a través del tiempo. Estos estudios han extendido la teoría evolutiva darwinista hasta procesos de nivel superior, que regulan tasas de especiación y extinción entre estirpes, lo que incluye la selección de especies, macroevolución y selección catastrófica de especies. Capitulo 9: El patrón arquitectónico de los animales (pg. 221) Desde los organismos relativamente simples que constituyen los comienzos de la vida en la Tierra, la evolución animal ha progresado a lo largo de una secuencia histórica de formas cada vez más complicadas. Los orgánulos se integraron en células, las células en tejidos, los tejidos en órganos y los órganos en sistemas. Mientras que un protozoo lleva a cabo todas las funciones vitales dentro de los límites de una única célula, un animal multicelular evolucionado es un complejo de unidades subordinadas organizadas en niveles sucesivos. Cada organismo ha heredado un modelo de organización descrito en términos de simetría corporal, numero de hojas embrionarias, grado de organización y numero de cavidades corporales. La mayoría de los animales presenta simetría bilateral, aunque en algunos grupos aparece simetría esférica o radial. La mayor parte de los animales son triblásticos y se desarrollan a partir de tres hojas embrionarias, pero los cnidarios y algunas otras formas son diblásticos. Las esponjas carecen de hojas embrionarias y poseen un grado de organización celular, mientras que el resto de los animales tienen un grado de organización tisular. Todos los animales, salvo las esponjas, tienen cavidad digestiva. En la mayor parte de ellos 3 existe otra cavidad que rodea a la anterior. Esta segunda cavidad puede ser un pseudocele o un celoma. Existen dos sistemas de formación del celoma, la enterocelia y la esquizocelila. Los animales triblásticos se dividen en protostomos y deuterostomos de acuerdo con su particular secuencia del desarrollo. A su vez, los protostomos se dividen en lofotrocozoos y ecdisozoos de acuerdo con caracteres del desarrollo más detallados. El cuerpo de los metazoos está formado por células, la mayoría de las cuales está especializada funcionalmente; fluidos corporales, divididos en intracelulares y extracelulares; y elementos estructurales extracelulares, fibrosos o amorfos, que desempeñan diversas funciones en el espacio extracelular. Las células de los metazoos se organizan en diversos tejidos, cuyos tipos básicos son el epitelial, el conjuntivo, el muscular y el nervioso. Los tejidos se organizan a su vez en unidades funcionales mayores, denominadas órganos, y, por último, los órganos se asocian para formar sistemas. Una consecuencia del incremento en la complejidad anatómica es un aumento del tamaño corporal, lo que ofrece ciertas ventajas, como una depredación efectiva, un menor coste energético en la locomoción y una mejora de la homeostasis. Capitulo 10: Clasificación y filogenia de los animales (pg. 239) La sistemática animal tiene tres objetivos fundamentales: (1) identificar todas las especies animales, (2) estudiar las relaciones evolutivas entre las especies, y (3) agrupar las especies animales en una jerarquía de grupos taxonómicos (taxones) que responda a las relaciones filogenéticas y las refleje. Los taxones están dotados de rango para incluir unos dentro de otros: especie, genero, familia, orden, clase, filo y reino. Todos estos rasgos pueden subdividirse para indicar taxones intermedios. Los nombres de las especies son binominales, con una primera denominación que designa el género al que pertenece la especie (con inicial mayúscula) seguida del epíteto especifico (en minúsculas), ambos escritos en letra cursiva. Los taxones del resto de los rangos tienen nombres simples, no escritos en cursiva. El concepto biológico de especie ha guiado el reconocimiento de la mayoría de las especies animales. La especie biológica se define como una comunidad reproductora de poblaciones (aislada de otras desde el punto de vista de la reproducción) que ocupa un nicho especifico en la naturaleza. No es inmutable a través del tiempo, sino que cambia con el curso de la evolución. Debido a que el concepto biológico de especie puede resultar de difícil aplicación en dimensiones espaciales y temporales, y al excluir además a las formas con reproducción asexual, se han propuesto otros conceptos alternativos: el concept o evolutivo de especie y el concepto filogenético de especie. Ninguno de los tres es seguido de forma unánime por los zoólogos. Actualmente hay dos escuelas taxonómicas principales. La taxonomía evolutiva tradicional agrupa las especies en taxones superiores de acuerdo con los criterios de ascendencia común y evolución adaptativa; tales taxones tienen un único origen evolutivo y ocupan una zona adaptativa propia. Un segundo punto de vista, conocido como sistemática filogenética o cladismo atiende exclusivamente a la ascendencia común para agrupar a las especies en taxones superiores. Solamente los taxones monofiléticos (aquellos que tienen un único origen evolutivo y contienen a todos los descendientes del antecesor común mas reciente del grupo) se utilizan en cladistica. Además de los taxones monofiléticos, la taxonomía evolutiva reconoce 4 ciertos taxones parafiléticos (con un único origen evolutivo, pero que excluyen a alguno de los descendientes del antecesor común mas reciente del grupo). Los taxones polifiléticos (aquellos que tienen más de un origen evolutivo) son excluidos por ambas escuelas taxonómicas. Tanto la taxonomía evolutiva como el cladismo requieren la comprobación de los patrones de ascendencia común entre las especies antes de erigir taxones de rango superior. Para ello se utiliza la morfología comparada (incluyendo el desarrollo embrionario), la citología y la bioquímica con el fin de reconstruir las relaciones jerárquicas entre los taxones que refleje la ramificación de los linajes evolutivos a lo largo del tiempo. El registro fósil proporciona estimaciones de la edad de los linajes evolutivos. Conjuntamente, el registro fósil y los estudios comparados nos permiten reconstruir un árbol filogenético que representa la historia evolutiva del reino animal. Tradicionalmente, todas las formas de vida se distribuyeron en dos reinos (animal y vegetal), pero recientemente se ha seguido un sistema de cinco reinos (animales, plantas, hongos, protistas y moneras). Ninguno de estos sistemas se adapta a los principios ni de la taxonomía evolutiva ni de la cladista, ya que sitúan a los organismos unicelulares en grupos parafiléticos o polifiléticos. Según el conocimiento actual del árbol filogenético de la vida, los “protozoos” no constituyen un grupo monofilético y no pertenecen al mundo animal. Las relaciones filogenéticas entre los filos animales se han aclarado con los estudios de filogenia molecular, aunque muchas de estas agrupaciones de alto nivel siguen siendo provisionales. Es particularmente controvertida la agrupación de los animales de simetría bilateral en los clados Deuterostomos, Protostomos, Ecdisozoos y Lofotrocozoos. Capitulo 11: Grupos de protozoos (pg. 273) El conjunto de organismos unicelulares y con rasgos “animales” se incluía antes ene l filo Protozoos. Actualmente se considera que el “filo” estaba compuesto de varios filos con diversas relaciones filogenéticas. El término “protozoo” se sigue manteniendo por conveniencia para referirse a todos estos organismos, tan diversos. En ellos se pone de manifiesto el gran potencial adaptativo de este modelo estructural básico de célula eucarióntica única. Ocupan un vasto conjunto de nichos y hábitat, y muchas especies tienen orgánulos complejos y especializados. Todos los protozoos tienen uno o varios núcleos, y estos a menudo se presentan al microscopio lumínico con aspecto vesicular. Los macronúcleos de los ciliados son compactos. A veces existen en los núcleos endosomas. Muchos protozoos tienen orgánulos semejantes a los que se encuentran en las células de los metazoos. El movimiento por pseudópodos o ameboide, que es un modo de locomoción y captura de alimento de los protozoos, tiene un papel vital y constituye en los metazoos un mecanismo de defensa. Este movimiento tiene lugar mediante el ensamblaje de subunidades de actina en microfilamentos y requiere un gasto energético procedente del ATP. El movimiento ciliar es igualmente importante, tanto en los protozoos como en los metazoos. Actualmente, el mecanismo mas comúnmente aceptado para interpretar el movimiento ciliar es la hipótesis de los microtúbulos deslizantes. Los protozoos pueden ser holofiticos (autótrofos), holozoicos (Heterotrofos) o saprozoicos. El exceso de agua que entra en el cuerpo de los protozoos es expelido por medio de vacuolas pulsátiles (vesículas de expulsión de agua.) La respiración y la eliminación de desechos tienen 5 lugar a través de toda la superficie del cuerpo. Los protozoos se reproducen asexualmente por bipartición (fisión binaria), división múltiple (pluripartición) o gemación; los mecanismos de reproducción sexual son también comunes. La formación de quistes, bajo el estimulo de condiciones ambientales adversas, es un importante mecanismo adaptativo de muchos protozoos. A la evolución de la célula eucarionte le siguió la diversificación en muchos clados, alguno de los cuales contienen formas unicelulares y pluricelulares. Los clados se han identificado en parte de acuerdo con los rasgos moleculares y pueden contener conjuntos de animales de filos tradicionales. Los miembros de algunos filos pueden ser especies fotoautótrofas, como en las Clorofíceas, los Euglenozoos y los dinoflagelados. Algunos miembros de estos grupos son organismos planctónicos muy importantes. Los Euglenozoos incluyen muchas especies no fotosintéticas, algunas de las cuales producen graves enfermedades en la especie humana, como la enfermedad del sueño de África y la enfermedad de Changas. Todos los apicomplejos son parásitos, como Plasmodium, causante de la malaria. Los cilióforos se mueven por medio de cilios o de orgánulos ciliares. Son un grupo grande y diverso, y muchos de ellos tienen estructuras complicadas. Las amebas se mueven mediante pseudópodos y actualmente están repartidas en varios filos. Capitulo 12: Mesozoos y Parazoos Los miembros del filo Mesozoos son animales de organización muy simple que parasitan los sacos renales de los moluscos cefalópodos (clase Rombozoos) y otros grupos de invertebrados (clase Ortonéctidos). Tienen solamente dos estratos de células, pero estas no son homologas de las capas germinales de los metazoos superiores. Tienen un ciclo vital complicado que aun no se conoce por completo. Su organización sencilla podría haber derivado de un antecesor más complejo semejante a un platelmitno. El filo Placozoos tiene solamente un único representante, un pequeño organismo marino laminar. También tienen solo dos capas de células, pero algunos autores consideran que estas capas son homologas del ectodermo y endodermo de los metazoos superiores. Los animales mas próximos a los Placozoos parecen ser los cnidarios. Las esponjas (filo Poríferos) son un grupo marino numeroso con algunos representantes de agua dulce. Tienen varias células especializadas, pero no están organizadas en tejidos u órganos. Dependen del batido flagelar de sus coanocitos para la circulación del agua a través de sus cuerpos, que es fundamental para conseguir el alimento y para el intercambio de gases respiratorios. Están sostenidas por esqueletos secretados de colágeno fibrilar, colágeno en forma de grandes fibras o filamentos (espongina), espículas silíceas o calcáreas, o una combinación de espículas y espongina en la mayoría de las especies. Las esponjas se reproducen asexualmente por gemación, fragmentación y gémulas (yemas internas). La mayor parte de las esponjas son monoicas aunque no producen simultáneamente espermatozoides y ovocitos. La embriogenia es inusual, con una migración de células flageladas desde la superficie hacia el interior (parenquímula) o la producción de una anfiblástula con inversión y crecimiento de los macromeros por encima de los micrómeros. Las esponjas tienen una gran capacidad de regeneración. Las esponjas son un antiguo grupo, en apariencia un pariente filogenéticamente remoto de 6 otros metazoos, pero la prueba molecular sugiere que son un grupo hermano de los eumetazoos. Su radiación adaptativa esta basada en la elaboración de un sistema de circulación del agua y de alimentación por filtración, excepto para una familia de esponjas, en las que la alimentación por filtración ha sido sustituida por la carnívora y donde la simbiosis bacteriana supone un aporte nutritivo extra. Capítulo 13: Los animales radiados (pg. 322) Los filos Cnidarios y Ctenóforos tienen una simetría radial primaria; la simetría radial es una ventaja para los organismos sésiles o que flotan libres, ya que los estímulos ambientales les llegan de todas las direcciones por igual. Los cnidarios son, sorprendentemente, eficaces depredadores ya que poseen orgánulos urticantes denominados nematocistos. Ambos filos son esencialmente diblásticos (algunos triblásticos, dependiendo de la definición de mesodermo), con una pared del cuerpo formada por epidermis y gastrodermis separadas por una mesoglea. La cavidad digestivo-respiratoria (gastrovascular) tiene boca y carece de ano. Los cnidarios poseen un nivel de organización tisular. Tienen dos tipos corporales básicos (polipoide y medusoide), y en muchos hidrozoos y escifozoos el ciclo incluye al polio, con reproducción asexual, y a la medusa, con reproducción sexual. Su orgánulo exclusivo, el cnido, se forma a partir de un cnidoblasto (del cual deriva el cnidocito) y se encuentra enrollado dentro de una capsula. Cuando se descargan, algunos tipos de cnidos llamados nematocistos penetran en la presa e inyectan veneno. La descarga se efectúa al incrementarse la presión hidrostática interna por la alta presión osmótica del interior de la capsula. La mayoría de los hidrozoos son coloniales y marinos, pero otros son dulciacuícolas, como las hidras de agua dulce que se muestran habitualmente en las clases prácticas. Las hidras tienen una forma polipoide típica pero no son coloniales y carecen de estado medusa. La mayor parte de los hidrozoos marinos son colonias ramificadas formadas por muchos pólipos (hidrantes). Las medudas de los hidrozoos pueden nadar libremente o permanecer unidas a la colonia. Los escifozoos son las medusas típicas, en los que la forma predominante es la medusoide y muchos tienen un estado polipoide poco aparente. Los cubozoos son predominantemente medusoides, dentro de los cuales se incluyen las peligrosas avispas de mar. Los antozoos son todos marinos y polipoides; no tienen estado medusoide. Las subclases más importantes son: Hexacorales (con simetría hexámera o polímera) y Octocorales (con simetría octómera). Los órdenes más numerosos de los hexacorales contienen las anemonas de mar, solitarias y sin esqueleto, y los corales pétreos, que son principalmente coloniales y secretan un exoesqueleto calcáreo. Los corales pétreos son los componentes básicos de los arrecifes coralinos, que constituyen un hábitat de gran belleza, productividad e importante valor ecológico y económico. Los octocorales agrupan a los corales blandos y córneos, muchos de los cuales son importantes y bellos componentes de los arrecifes. Los Ctenóforos son birradiales y nadan mediante ocho filas de peines. Los coloblastos, con los que capturan las presas, son característicos de este filo. Los cnidarios y los ctenóforos posiblemente han derivado de un antecesor parecido a una larva plánula de los cnidarios. A pesar de su nivel de organización relativamente simple, los cnidarios constituyen un filo importante económico, ambiental y biológicamente. 7 Capítulo 14: Los animales bilaterales acelomados (pg. 347) Los Platelmintos, los Nemertinos y los Gnatostomúlidos son los filos más simples con simetría bilateral, una condición de valor adaptativo para los animales activos reptantes o nadadores. Carecen de celoma o pseudoceloma (con la posible excepción de los Nemertinos) y, por tanto, son acelomados. Son triblásticos y con un nivel de organización de órganos y sistemas. La superficie externa de los turbelarios está cubierta por un epitelio celular, ciliado, al menos en parte, que contiene numerosas células mucosas y rabditos en forma de varilla, que actúan juntos en la locomoción. Los miembros de las demás clases de platelmintos presentan un tegumento sincitial no ciliado, con un citoplasma distal vesicular y cuerpos celulares debajo de las capas musculares superficiales. En la mayoría la digestión es extracelular e intracelular; los cestodos deben absorber nutrientes predigeridos a través del tegumento, ya que carecen de tracto digestivo. La osmorregulación se realiza mediante las células flamígeras de los protonefridios, y la expulsión de los desechos metabólicos y la respiración se efectúa por difusión a través de la pared del cuerpo. Excepto los Acelos, los platelmintos tienen un sistema nervioso de tipo escalera, con neuronas motoras, sensoriales y de asociación. La mayor parte de los platelmintos son hermafroditas y muchos también se reproducen asexualmente. La clase Turbelarios es un grupo parafilético, con miembros en su mayoría carnívoros y de vida libre. Los trematodos digeneos tienen un molusco como hospedador intermediario y casi siempre un vertebrado como hospedador definitivo. La enorme tasa de reproducción asexual que tiene lugar en el hospedador intermediario aumenta la probabilidad de que alguno de los descendientes alcance al hospedador definitivo. Aparte del tegumento, los digeneos comparten muchos rasgos estructurales básicos con los turbelarios, Los digeneos incluyen muchos parásitos importantes del hombre y los animales domésticos. Contrastan con los monogeneos, que son importantes parásitos de peces y tienen un ciclo vital directo (sin hospedador intermediario). Los cestodos, o tenias, poseen generalmente un escólex en su extremo anterior, seguido de una larga cadena de proglótides cada uno de los cuales tiene un juego completo de órganos reproductores de ambos sexos. Como adultos, viven en el tubo digestivo de vertebrados. La superficie de absorción de su tegumento esta aumentada debido a la presencia de microtricos. Las larvas, con cascara, pasan a las heces, y los juveniles se desarrollan en un hospedador intermediario vertebrado o invertebrado. Los miembros de los Nemertinos tienen un tubo digestivo completo con ano y un verdadero aparato circulatorio. Son animales de vida libre, principalmente marinos, y capturan sus presas enredándolas con su larga probóscide extensible. Los Gnatostomúlidos constituyen un curioso filo de pequeños gusanos marinos que viven entre los granos de los sedimentos del fondo. No tienen ano, y comparten ciertas características con una amplia diversidad de grupos como turbelarios, rotíferos, esponjas y cnidarios. El resultado es que sus relaciones con los otros filos permanecen muy inciertas. Los platelmintos y los cnidarios proceden, probablemente, de un antecesor común (planuloide), algunos de cuyos descendientes se volvieron sésiles o flotadores de vida libre y radial (cnidarios), y otros reptantes y bilaterales (platelmintos). El análisis secuencial del rDNA, así como algunos criterios morfológicos y del desarrollo, sugieren que los Acelos, considerados hasta ahora como un orden de los turbelarios, 8 divergieron de un antecesor compartido con otros Bilaterales y constituyen el grupo hermano de todos los otros filos bilaterales. Capitulo 15: Los animales pseudocelomados (pg. 373) Los filos tratados en este capitulo poseen una cavidad corporal denominada pseudocele, que deriva del blastocele embrionario en vez de una cavidad secundaria de mesodermo (celoma). Varios grupos presentan eutelia, esto es, un número constante de células o núcleos en el individuo adulto de una especie dada. El análisis de semejanza de nucleótidos en el gen para la subunidad menor del 18S del rDNA proporciona pruebas de que algunos filos de pseudocelomados pertenecen al superfilo Ecdisozoos (Nematodos, Nematromorfos, Kinorrincos y Priapúlidos) y otros al superfilo Lofotrocozoos (Rotíferos, Acantocéfalos, Gastrotricos y Endoproctos). De acuerdo con el análisis cladista, los Loricíferos probablemente sean ecdisozoos. Los Nematodos son con mucho el grupo mas grande y representativo de este filo, y aunque hoy solamente hay descritas 25000 especies, se estima que podría contar con más de 500000 especies. Son más o menos cilíndricos, con los extremos terminados en punta y cubiertos por una cutícula resistente y diferenciada. La musculatura de la pared del cuerpo es exclusivamente longitudinal y encierra un volumen de líquido en el pseudocele a alta presión hidrostática que interviene en la locomoción. La presión hidrostática tiene un efecto fundamental en diversas funciones fisiológicas ce los nematodos, como la ingestión de alimento, la eliminación de las heces, la excreción y la copula. Muchos nematodos son dioicos, con cuatro estados juveniles cada uno de ellos separado por la muda de la cutícula. Casi todos los animales y muchas plantas tienen nematodos parásitos, pero muchos otros nematodos viven libres en el suelo y en las plantas acuáticas. Algunos nematodos parásitos tienen parte de su ciclo vital de vida libre, otros sufren una migración por los tejidos del hospedador y algunos tienen hospedadores intermediarios en su ciclo vital. Algunos de los nematodos parásitos causan daños graves a otros animales y a las personas. Los Nematomorfos, parecidos superficialmente a los nematodos, tienen estados juveniles parásitos de artrópodos y un estado adulto de vida libre, acuático. Kinorrincos y Loricíferos son pequeños filos de animales acuáticos pseudocelomados. Los kinorrincos se fijan e impulsan mediante las espinas de su cabeza. Los Loricíferos pueden retraerse en su loriga. Los Priapúlidos son gusanos marinos excavadores de tamaño medio. El filo Rotíferos está constituido por organismos fundamentalmente dulciacuícolas y pequeños, con una corona ciliada que produce una corriente de agua que arrastra alimento planctónico hacia la boca; ésta desemboca en una faringe muscular, o mástax, que está provista de mandíbulas. Los Bdeloideos son partenogenéticos obligados y parece que en este grupo no hay machos. En estado adulto todos los Acantocéfalos son parásitos del intestino de los vertebrados y sus estados juveniles se desarrollan en los artrópodos. Tienen una probóscide anterior invaginable armada con espinas, que fijan a la pared intestinal del hospedador. No tienen tubo digestivo, y deben absorber todos los nutrientes a través del tegumento. Las pruebas moleculares sugieren una afinidad filogenética de acantocéfalos y rotíferos. Los Gastrotricos también son diminutos pseudocelomados acuáticos. Se mueven por cilios o glándulas adhesivas. Los Endoproctos son animales acuáticos, pequeños, sésiles, con una 9 corona de tentáculos ciliados alrededor de la boca y del ano. De todos estos filos, los Nematodos han alcanzado un enorme éxito evolutivo y una gran radiación adaptativa. Capítulo 16: Los Moluscos (pg. 407) Los moluscos son el filo más grande de los lofotrocozoos y uno de los filos mayores y más diversos; sus miembros tienen un tamaño que varía desde muy pequeños a los mayores invertebrados. Las partes básicas del cuerpo son la masa visceral, comúnmente cubierta por la concha, la cabeza y el pie. La mayoría son marinos, pero algunos son de agua dulce y unos pocos terrestres. Ocupan nichos muy diversos. Algunos son importantes desde el punto de vista económico y otros, como huéspedes de parásitos, desde el punto de vista sanitario. Los moluscos son celomados, aunque su celoma está limitado al área circundante al corazón y gónadas y ocasionalmente parte del intestino. El desarrollo evolutivo de un celoma fue importante debido a que permitió una mejor disposición de los órganos viscerales y, en muchos de los animales que lo tienen, un eficaz esqueleto hidrostático. El manto y la cavidad del manto son características importantes de los moluscos. El manto secreta la concha y cubre una parte de la masa visceral para formar una cavidad que aloja a las branquias. En algunos moluscos la cavidad del manto se ha modificado en un pulmón. El pie es normalmente un órgano locomotor ventral, en forma de suela, pero puede estar ampliamente modificado, como en los cefalópodos, en los que se ha transformado en un embudo. La rádula está presente en todos los moluscos, excepto los bivalvos y los solenogastros; es un órgano con dientes, en forma de lengua, extensible, utilizada en la alimentación. Excepto en los cefalópodos, que poseen un sistema circulatorio cerrado, el aparato circulatorio de los moluscos es abierto, con corazón y senos sanguíneos. Por lo general los moluscos tienen un par de nefridios que conectan con el celoma, y un sistema nervioso complejo con diferentes órganos sensoriales. La larva primitiva de los moluscos es la trocófora, y la mayor parte de los moluscos marinos tienen una larva mas evolucionada, la velígera. Las clases Caudofoveados y Solenogastros son pequeños grupos de moluscos vermiformes sin concha. Los Escafópodos constituyen una clase ligeramente mayor, con una concha tubular abierta en sus extremos y el manto rodeando el cuerpo. La clase Monoplacóforos es un pequeño grupo marino, univalvo, que muestra pseudometamería. Los Poliplacóforos, más comunes, son organismos marinos con una concha formada por ocho placas. Son animales sedentarios con una fila de branquias a lo largo de cada lado del pie. Los Gasteropodos constituyen la clase mayor y de más éxito evolutivo. Su interesante historia evolutiva incluye la torsión o giro del extremo posterior del cuerpo hacia delante, de tal manera que el ano y la cabeza están en el mismo extremo, y el arrollamiento, un alargamiento y espiralización de la masa visceral. La torsión llevo consigo el problema de la contaminación por la liberación de los productos de desecho frente a las branquias y sobre la cabeza; esto se ha resuelto de diversas formas en los diferentes gasterópodos. Una de las soluciones a la contaminación es introducir el agua por un lado de la cavidad del manto y sacarla por el otro (muchos gasterópodos), algún grado de detorsión (opistobranquios), y la conversión de la cavidad del manto en un pulmón (pulmonados). Los miembros de la clase Bivalvos son marinos y dulciacuícolas; so concha está dividida en dos valvas unidas por un ligamento dorsal y mantenidas juntas por un musculo aductor. La mayor 10 parte de ellos son filtradores, e inhalan agua a través de sus branquias por acción ciliar. Los representantes de la clase Cefalópodos son los moluscos más evolucionados; son todos depredadores y muchos pueden nadar rápidamente. Sus tentáculos capturan presas mediante secreciones adhesivas y por medio de ventosas. Nadan expeliendo violentamente el agua de su cavidad del manto por el embudo, que ha derivado del pie. Hay tanto pruebas embriológicas como moleculares de que los moluscos comparten un antecesor común con los anélidos mas reciente que con los artrópodos o los filos de los deuteróstomos. Sin embargo, queda un punto importante de discusión sobre si los moluscos surgieron de los Lofotrocozoos y de sus relaciones con los otros filos de protóstomos. Capítulo 37: La biosfera y la distribución de los animales (pg. 931) La biosfera es la delgada capa que rodea la tierra y mantiene todas las formas vivas. La vida sobre la tierra es posible gracias al aporte constante de energía por parte del sol, la presencia de agua, unas variaciones de temperatura idóneas, la correcta proporción de los elementos principales y accesorios, y la retención de la letal radiación ultravioleta por el ozono atmosférico. El ambiente terrestre y los organismos que viven en él evolucionaron juntos, dependiendo enormemente entre sí. La biosfera está compuesta por la litosfera, la cubierta rocosa de la Tierra, la hidrosfera, la masa total de agua, y la atmosfera, la capa gaseosa que rodea la tierra. El medio terrestre está constituido por distintos biomas, cada uno con una flora característica y una fauna asociada a ella. Los bosques caducifolios se distinguen por su alternancia estacional y por la caída de las hojas en el otoño. Más al norte de los bosques caducifolios, están los bosques de coníferas, un bioma dominado por arboles con hojas en forma de aguja, bien adaptados para soportar la dura estación de las nieves, y cuyo límite mas norteño forma lo que se denomina la taiga. Los animales de la taiga están adaptados para soportar unos inviernos largos y con mucha nieve. La selva tropical, el más rico de todos los biomas, se caracteriza por la enorme diversidad de sus especies vegetales y por una estratificación vertical de los hábitats de los animales. La mayor parte de los suelos de las selvas tropicales se degradan rápidamente cuando el bosque es destruido. El bioma mas modificado por el hombre es el de los pastizales o praderas, que han sido transformados en su mayoría con fines agrícolas y ganaderos. La tundra del extremo septentrional y el desierto son ambientes muy desfavorables para la vida animal, pero están ocupados por animales que han desarrollado adaptaciones apropiadas. Los ambientes de aguas dulces incluyen los ríos y riachuelos (hábitat loticos) y los estanques y lagos (hábitat lenticos). Son ambientes geológicamente poco duraderos y que dependen enormemente del aporte de nutrientes. Los océanos ocupan el 71% de la superficie de la Tierra. La zona fótica, o iluminada, es donde se produce la actividad fotosintética del fitoplancton. Las comunidades animales oceánicas se clasifican según sus características topográficas, el sustrato y la distancia de la orilla. Las comunidades bentónicas con sustratos rocosos incluyen la franja intermareal con pozas de 11 marea, los arrecifes de coral, los bosques submareales de kelpos, y las comunidades de los afloramientos hidrotermales profundos. Las comunidades bentónicas de alta mar forman parcelas con suspensívoros que habitan en los sustratos arenosos y detritívoros en los fangosos. Los arrecifes de coral son la comunidad bentónica que presenta una mayor diversidad ecológica. Las comunidades pelágicas incluyen una zona de aguas poco profundas sobre la plataforma continental. EN esta zona es donde se encuentran los grandes bancos pesqueros del mundo, que son especialmente productivos en las áreas de afloramiento donde los nutrientes se renuevan constantemente. Las aguas más profundas de alta mar ocupan la mayor parte del océano pero tienen una baja productividad biológica. La zoogeografía es el estudio de la distribución de los animales sobre la Tierra y de su historia, mediante el empleo de los datos procedentes de las distribuciones actuales de las especies animales, el registro fósil y los análisis de la sistemática filogenética. Los animales se han distribuido sobre la Tierra por dispersión, la separación de poblaciones desde sus centros de origen, y por vicarianza, la separación de poblaciones por la aparición de barreras. La deriva continental, ahora muy apoyada por la teoría de la tectónica de placas, ayuda a explicar la forma en que los grupos de animales han podido separarse y diversificarse evolutivamente. También explica la manera en que ciertos grupos, como los mamíferos marsupiales, han podido aislarse unos de otros. Los puentes temporales terrestres también sirvieron como importantes vías para la dispersión de los animales. Capitulo 38: Ecología animal (pg. 952) La ecología es el estudio de las relaciones entre los organismos y sus ambientes para tratar de explicar la distribución y la abundancia de las especies sobre la Tierra. El espacio físico que contiene el ambiente de un animal, es el hábitat. En el hábitat se dan las condiciones físicas y biológicas adecuadas para la supervivencia y la reproducción y, en conjunto, constituyen el nicho de un animal, una población o una especie. Las poblaciones de los animales están formadas por demás cuyos miembros se reproducen entre si y comparten un fondo genético. Las cohortes de los animales tienen unos modelos de supervivencia característicos que representan un equilibrio entre los cuidados parentales y la cantidad de descendientes. Las poblaciones de animales están formadas por cohortes superpuestas con una estructura de edades característica e indicativa de si están creciendo, reduciéndose o en equilibrio. En la naturaleza, cada especie posee un ritmo intrínseco de crecimiento que representa su potencial para crecer de manera exponencial. Las poblaciones humanas están creciendo de manera exponencial, a un ritmo aproximado del 1.23% al año, y se espera que pasen de los 6400 millones de personas que viven en la actualidad a 8000 millones en el año 2040. El crecimiento de una población puede estar regulado intrínsecamente, por la capacidad de soporte del ambiente, o extrínsecamente, por la competencia entre especies por un recurso limitante, o por los depredadores y los parásitos. La densidad de una población es independiente de los factores abióticos que pueden limitar, pero no regular verdaderamente, el crecimiento de dicha población. Las comunidades están formadas por poblaciones que interactúan unas con otras por competencia, depredación, parasitismo, comensalismo y mutualismo. Estas relaciones son el resultado de una coevolución entre las poblaciones de las comunidades. Los gremios de 12 especies evitan la exclusión competitiva por la existencia de una diferenciación característica, el reparto de los recursos limitantes mediante la especialización morfológica. Los depredadores clave controlan la estructura de la comunidad y reducen la competencia entre las presas, lo que incrementa la diversidad de especies. Los parásitos y sus hospedadores han desarrollado unas relaciones benignas que aseguran su convivencia. Los ecosistemas están formados por comunidades y los ambientes abióticos de estas. Los animales que ocupan los niveles tróficos de los herbívoros y los carnívoros son los consumidores de los ecosistemas. Todos los organismos tienen un presupuesto energético, resultado de la producción bruta, la producción neta y la respiración. Para los animales, la respiración normalmente representa más del 90% de su presupuesto energético. Por ello la transferencia de energía de un nivel trófico al siguiente se limita solo a un 10%, lo que limita el número de niveles tróficos de un ecosistema. Las pirámides ecológicas de energía representan gráficamente la disminución de la producción en los sucesivos niveles tróficos de las redes alimentarias. La producción de un ecosistema es el resultado del flujo de energía y de los ciclos de los materiales en los ecosistemas. Toda la energía termina perdiéndose en forma de calor, pero los nutrientes y otras sustancias, incluidos los pesticidas, se reciclan. Ningún ecosistema está totalmente cerrado, ya que todos ellos dependen de los intercambios de energía y de materiales desde fuentes externas. La biodiversidad existe porque las tasas de especiación, en promedio, han sido ligeramente superiores a las tasas de extinción a lo largo de la historia evolutiva de la tierra. Aproximadamente, el 99% de todas las especies que han existido ahora están extintas. Las tasas de extinción en el pasado geológico son muy episódicas, y han tenido unos calores que, en diferentes intervalos de 1 millón de años, han oscilado desde próximos a cero hasta cercanos al 96%. Las especies que tienen distribuciones geográficas grandes presentan unos índices medios de extinción menores que los de las especies con distribuciones más reducidas, y esta misma relación se mantiene para los taxones de mayor rango. Los estudios paleontológicos de la extinción proporcionan una perspectiva importante para evaluar las posibles consecuencias evolutivas de la extinción de especies debida a las actividades humanas. Por Álvaro Vallvé Fritschi Resúmenes del Hickman: Principios Integrales de Zoología (13ª Ed.) Segundo Cuatrimestre Capítulo 17: Los Gusanos Segmentados (pg. 428) El filo Anélidos es un gran grupo cosmopolita, que incluye los poliquetos marinos, las lombrices de tierra, los oligoquetos de agua dulce y las sanguijuelas. Sin duda, la innovación estructural mas importante, fundamental en la diversificación de este grupo, es el metamerismo: la división del cuerpo en una serie de segmentos similares, cada uno de los cuales contiene 13 diversos órganos y sistemas en una disposición repetida. En los anélidos, el celoma está muy desarrollado, y esto, junto con la disposición de compartimientos llenos de líquido y un eficaz esqueleto hidrostático para una excavación precisa y movimientos de natación. Los artrópodos presentan especializaciones segmentarias adicionales, que se trataran en los Capítulos 18, 19 y 20. Los poliquetos constituyen la mayor clase de los anélidos y son casi todos marinos. Cada segmento tiene un par de parapodios que llevan muchas sedas. Los parapodios muestran una gran variedad de adaptaciones entre los poliquetos, entre las que se incluyen especializaciones para la natación, respiración, culebreo, mantenimiento de la posición en la galería, bombeo de agua a través de la galería y como órganos accesorios para la alimentación. Algunos son principalmente depredadores y tienen una faringe protráctil con mandíbulas. Otros poliquetos rara vez abandonan las galerías o tubos en los que viven. Entre los miembros de este grupo aparecen varios tipos de alimentación por filtración o detritívora. Los poliquetos son dioicos, tienen un sistema reproductor primitivo, carecen de clitelo, la fecundación es externa y presentan larva trocófora. La clase Oligoquetos incluye las lombrices de tierra y muchas formas de agua dulce; tienen un pequeño número de sedas por segmento (comparado con los poliquetos) y carecen de parapodios. Tienen un sistema circulatorio cerrado y el vaso dorsal es el principal órgano bombeador. En casi todos los metámeros hay un par de nefridios. Las lombrices de tierra presentan el sistema nervioso típico anelidiano: un cerebro dorsal conectado a un doble cordón nervioso ventral, que recorre el cuerpo del animal, con ganglios segmentarios. Los oligoquetos son hermafroditas, y practican la fecundación cruzada. El clitelo tiene un importante papel en la reproducción; interviene en la producción del moco que rodea a los gusanos durante la cópula y en la secreción del capullo que recibe los huevos y el esperma, dentro del cual transcurre el desarrollo embrionario. Del capullo sale un pequeño gusano joven. Las sanguijuelas (clase Hirudíneos) son principalmente dulciacuícolas, aunque unas pocas son marinas y otras terrestres. Por lo general se alimentan de líquidos, pero muchas son depredadoras, algunas parasitas temporales y unas pocas parasitas permanentes. Las sanguijuelas son hermafroditas y se reproducen de una manera semejante a los oligoquetos, con fecundación cruzada y formación de un capullo por el clitelo. Las pruebas embriológicas sitúan a los anélidos con los moluscos y los artrópodos en los Protóstomos. Las recientes pruebas moleculares sugieren que los anélidos y los moluscos están más estrechamente relacionados entre sí (en el superfilo Lofotrocozoos) que cada uno de ellos con los artrópodos (en el superfilo Ecdisozoos). Capítulo 18: Los Artrópodos (pg. 444) Los artrópodos constituyen el mayor filo y de más éxito en el mundo. Son animales celomados, protostomos, ecdisozoos y segmentados, con sistemas de órganos bien desarrollados. La mayoría muestran una marcada tagmatización. Presentan una gran diversidad y aparecen en todos los ambientes capaces de soportar vida. Quizás más que ningún otro factor, el éxito de los artrópodos se explica por su exoesqueleto cuticular, que ha hecho posible las distintas adaptaciones. Otros importantes elementos de su éxito son los apéndices articulados, la respiración traqueal, los órganos sensoriales eficaces, el comportamiento complejo y la metamorfosis. 14 Los trilobites son actualmente un subfilo extinguido que fue dominante en el Paleozoico. Los miembros del subfilo Quelicerados no tienen antenas y sus principales apéndices para la alimentación son los quelíceros. Además, tienen pedipalpos (que pueden parecerse a patas marchadoras) y cuatro pares de patas marchadoras. La clase Merostomados incluye a los euriptéridos (extinguidos) y al antiguo grupo, aunque todavía existente, de los cangrejos cacerola. La clase Picnogónidos contiene a las arañas de mar, animales extraños de pequeño tamaño que presentan una larga probóscide suctora y un abdomen vestigial. La gran mayoría de los quelicerados vivientes pertenecen a la clase Arácnidos: arañas (Orden Arañas), escorpiones (Orden Escorpiones), segadores (Orden Opiliones), garrapatas y ácaros (Orden Ácaros) entre otros. Los tagmas de las arañas (cefalotórax y abdomen) no presentan segmentación externa y están unidos por un pedicelo, una especie de cintura. Las arañas son depredadoras, sus quelíceros están provistos de glándulas de veneno para paralizar a sus presas. Respiran por pulmones en libro, tráqueas, o ambos. Las arañas pueden tejer hilos de seda que utilizan para distintos fines, entre ellos atrapar a sus víctimas. Los caracteres distintivos de los escorpiones son sus grandes pedipalpos quelados, su peine y su abdomen claramente segmentado y provisto de un aparato terminal punzante. Los segadores tienen cuerpos ovoides, pequeños, con patas muy largas y delgadas. Su abdomen está ampliamente unido al cefalotórax. El cefalotórax y el abdomen de las garrapatas y los ácaros están completamente fusionados, y las piezas bucales están situadas en un capítulo en la región anterior. Son los más numerosos de los arácnidos, y algunos son importantes transmisores de enfermedades y de graves plagas de plantas. Capítulo 19: Los mandibulados acuáticos (pg. 468) Además de un par de mandíbulas, los Crustáceos y los unirrámeos tienen en común al menos un par de antenas y un par de maxilas. Sus tagmas son: cabeza y tronco, o cabeza, tórax y abdomen. Los Crustáceos son un gran subfilo, principalmente acuáticos. Tienen dos pares de antenas, sus apéndices son ancestralmente birrámeos, y muchos tienen un caparazón. Todos los artrópodos deben mudar su cutícula periódicamente (ecdisis) y aumentar su tamaño antes de que la cutícula recién secretada se endurezca de nuevo. Los períodos de premuda y posmuda están controlados hormonalmente, asi como algunos otros procesos, tales como los cambios de color del cuerpo y la diferenciación de las características sexuales. Los hábitos alimentarios de los crustáceos varían ampliamente; hay muchos depredadores, carroñeros, filtradores y parásitos. La respiración tiene lugar a través de la superficie del cuerpo o mediante branquias. Los órganos excretores toman la forma de glándulas maxilares o antenales. La circulación, como en otros artrópodos, se realiza a través de un sistema abierto de senos (hemocele) y el principal órgano bombeador es un corazón dorsal y tubular. La mayor parte de los crustáceos tienen ojos compuestos formados por unidades denominadas omatidios. Los sexos están normalmente separados. Los crustáceos de la clase Branquiópodos se caracterizan por tener filopodios y contener, entre otros, al orden Cladóceros, que es un grupo ecológicamente importante como zooplancton. Dentro de la clase Maxilópodos, los miembros de la subclase Copépodos carecen de caparazón 15 y de apéndices abdominales. Son muy abundantes y se encuentran entre los más importantes consumidores primarios en muchos ecosistemas marinos y de agua dulce. Muchos son parásitos. La mayor parte de los miembros de la subclase Cirrípedos (percebes) son sésiles en estado adulto, secretan una concha de placas calcáreas, y filtran mediante sus apéndices torácicos. Los Malacostráceos forman la mayor clase de crustáceos, y los órdenes más importantes son los Isópodos, Anfípodos, Eufansiáceos y Decápodos. Todos tienen tanto apéndices torácicos como abdominales. Los Isópodos carecen de caparazón y generalmente son aplanados dorsoventralmente. Los Anfípodos también carecen de caparazón, pero suelen ser comprimidos lateralmente. Los Eufansiáceos constituyen el importante plancton oceánico llamado Krill. Los Decápodos incluyen a los cangrejos, camarones, bogavantes, cangrejos de río y otros; tienen cinco pares de patas marchadoras (incluyendo los quelípedos) en el tórax. Los Mandibulados Terrestres (pg. 503) Los miembros del subfilo Unirrámeos presentan apéndices unirrámeos y en la cabeza tienen un par de antenas, un par de mandíbulas y dos pares de maxilas (un par de maxilas en los milpiés). Los tagmas son: Cabeza y tronco en los miriápodos, y cabeza, tórax y abdomen en los insectos. Los insectos constituyen la clase más amplia del mayor de los filos animales. Los insectos se identifican fácilmente por la combinación de sus tagmas y por tener tres pares de patas torácicas. El éxito evolutivo de los insectos se explica por la posesión de una serie de características que les ha permitido explotar los biotopos terrestres, como las alas (presentes en la mayoría) la impermeabilidad de su cutícula y otros mecanismos, para reducir la perdida de agua y la capacidad de entrar en letargo durante los periodos en que las condiciones son adversas. La mayoría de los insectos tienen dos pares de alas en el tórax, aunque algunos solo tienen un par y otros ninguno. En algunos insectos los movimientos de las alas están controlados por músculos del vuelo directos, que se insertan directamente en la base de las alas. En otros, los músculos del vuelo son indirectos y mueven las alas por cambios en la forma del tórax. Cada contracción de los músculos de vuelo sincrónicos requiere un impulso nervioso, mientras que los músculos de vuelo asincrónico se contraen mucho tiempo por cada impulso nervioso. Los hábitos alimentarios varían mucho dentro de los insectos, y hay una enorme diversidad de especializaciones de aparatos bucales que reflejan los particulares hábitos alimentarios de un determinado insecto. Respiran mediante un sistema traqueal, un conjunto de tubos que se abren al exterior por espiráculos situados en el tórax y en abdomen. Los órganos excretores son tubos de Malpigio que flotan libres en el hemocele. Los sexos son separados y la fecundación es generalmente interna. Casi todos los insectos sufren una metamorfosis durante su desarrollo. En la metamorfosis de los hemimetábolos (incompleta), los estados juveniles se denominan inmaduros o ninfas, y el adulto emerge tras la última muda ninfal. En la metamorfosis de los holometábolos (completa), la última muda produce un estado inactivo (pupa). De la pupa emerge, finalmente, un adulto con alas. Ambos tipos de metamorfosis son controlados hormonalmente. Los insectos son importantes para el bienestar humano, ya que polinizan los cultivos de plantas, controlan las poblaciones de otros insectos perjudiciales mediante la depredación y el 16 parasitismo, y sirven de alimento a otros animales. Muchos insectos son perjudiciales para los intereses humanos porque infligen grandes daños en cosechas, alimento, bosques, ropa y haciendas, y muchos son vectores de importantes enfermedades humanas y de animales domésticos. Aunque algunos especialistas consideran que los miriápodos forman un clado con los quelicerados, la mayor parte de las pruebas apoyan un clado mandibulado, formado por miriápodos, insectos y crustáceos. Las patas desaparecieron del abdomen de los insectos y están, actualmente, confinadas en cada uno de los tres segmentos torácicos. Los insectos endognatos y ectognatos son grupos hermanos. Las alas de los insectos alados ancestrales pudieron derivar de branquias externas de ninfas acuáticas o de adultos, y pudo ser un estado con expansiones tergales (o proalas), articuladas o no, el que sirvió para realizar una termorregulación eficaz hasta que alcanzaron un tamaño para formar verdaderas alas aerodinámicas funcionales. La diversidad adaptativa y el número de especies y de individuos de los Insectos es enorme. Capítulo 21: Los protóstomos menores (pg. 523) Los diez filos “menores” estudiados en este capítulo se han reunido juntos aquí por conveniencia. Los experimentos de secuenciación molecular siguieren incluir a algunos en el superfilo Lofotrocozoos (Equiúridos, Sipuncúlidos, Pogonóforos, Braquiópodos, Ectoproctos y Foronídeos) y a otros en el superfilo Ecdisozoos (Pentastómidos, Onicóforos, Tardígrados y Quetognatos). Muchos autores siguen debatiendo sobre la naturaleza protóstoma de estos filos debido a sus rasgos de tipo deuteróstomo. Queda mucho por investigar antes de poder resolver las relaciones de estos grupos de forma definitiva. Los sipuncúlidos son gusanos marinos pequeños, minadores, con un introverto extensible en el extremo anterior. La probóscide lleva unos tentáculos con los cuales se alimentan del sustrato. Los sipuncúlidos no están segmentados. Los equiúridos son más diversos que los sipuncúlidos, pero hay un menor número de especies. También son excavadores vermiformes, y la mayoría son detritívoros, con una probóscide por delante de la boca. Tampoco son segmentados, pero la validez de este grupo como filo es objeto de debate. Los pogonóforos viven en tubos en los fondos oceánicos profundos, y son metamericos. No tienen ni boca ni tubo digestivo, pero absorben los nutrientes mediante la corona de tentáculos de su extremo anterior. Obtienen la mayor parte de su energía de bacterias quimioautótrofas alojadas en el trofosoma. Muchos expertos consideran actualmente que este filo no es válido, y en su lugar clasifican juntos a pogonóforos y vestimentíferos como miembros de una familia de Anélidos, los Siboglínidos. Los Foronídeos, los Ectoproctos y los Braquiópodos presentan un lofóforo, o corona de tentáculos ciliados alrededor de la boca pero no del ano, que contiene una prolongación del mesocele. Son sésiles en estado adulto, tienen un tubo digestivo en forma de U y una larva nadadora. Las funciones del lofóforo son respiratorias y alimenticias, creando con sus cilios una corriente de agua cuyas partículas filtra. Los foronídeos son los lofoforados menos comunes; viven en tubos en aguas costeras poco profundas. El lofóforo se extiende fuera del tubo para alimentarse. 17 Los ectoproctos son abundantes en los hábitat marinos, donde viven sobre diversos sustratos sumergidos; un cierto número de especies son comunes en aguas dulces. Los ectoproctos son coloniales, y aunque cada individuo es bastante pequeño, las colonias tienen generalmente varios centímetros de diámetro. Cada individuo vive en una cámara (zoecio), constituida por un exoesqueleto secretado por la epidermis, de naturaleza calcárea, quitinosa o gelatinosa. Los braquiópodos fueron my abundantes en el Paleozoico. Pero han venido declinando desde el principio del Mesozoico. Sus cuerpos y lofóforos están cubiertos por un manto, que segrega una valva ventral y otra dorsal (concha). Generalmente se encuentran unidos al sustrato directamente o mediante un pedúnculo. Los pentastómidos son parásitos vermiformes de los pulmones y los conductos nasales de los vertebrados carnívoros. Están relacionados con los artrópodos y muchos autores creen que deberían abandonar el estatus de filo y pasar a clasificarse con los crustáceos. Los onicóforos son animales parecidos a orugas que se encuentran en ambientes húmedos, fundamentalmente tropicales. Son metamericos y se mueven mediante una serie de apéndices no articulados pero provistos de garras. Los tardígrados son animales pequeños, mayoritariamente terrestres, que viven en la película de agua que cubre musgos y líquenes. Tienen cuatro pares de patas no articuladas y una cutícula no quitinosa. La cavidad principal del cuerpo es un hemocele, como en los artrópodos. Pueden sufrir criptobiosis, soportando condiciones adversas durante largos periodos. Los gusanos flecha o sagitas (filo Quetognatos) son un grupo pequeño, pero tienen importancia como componentes del plancton marino. Tienen un celoma bien desarrollado. Y son depredadores eficaces, que capturan otros organismos planctónicos con los dientes y las cerdas quitinoideas que rodean su boca. Las relaciones de estos taxones entre si y con otros filos de protostomos y deuterostomos sigue siendo objeto de debate y de interés científico considerables. En algunos casos ( como en los pogonóforos), los rasgos moleculares, morfológicos y embriológicos cuestionan la validez del estatus de filo para estos razones. En otros (como los equiúridos) los datos moleculares y los morfoembriológicos no están de acuerdo sobre la validez taxonómica y las relaciones entre determinados grupos. Queda mucha investigación por hacer antes de que la taxonomía y las relaciones de estos grupos, entre si y con otros filos de protostomos y deuterostomos, puedan resolverse inequívocamente. Capítulo 22: Los equinodermos y los hemicordados (pg. 552) El filo Equinodermos muestra las características del grupo de los Deuteróstomos del reino animal. Son un grupo marino importante claramente discernible de cualquier otro grupo de animales. Presentan simetría pentarradial, pero derivan de antecesores bilaterales. Las estrellas de mar (clase Asteroideos) pueden utilizarse como ejemplo de los equinodermos. Generalmente tienen cinco brazos, que emergen gradualmente de un disco central. Como el resto de los equinodermos, carecen de cabeza y poseen muy pocos órganos sensoriales. La boca está dirigida hacia el sustrato. Tienen osículos dérmicos, papulas respiratorias y surcos ambulacrales abiertos. Muchas estrellas de mar poseen pedicelarios. Su aparato vascular acuífero es un complejo sistema hidráulico, derivado embriológicamente de uno de los compartimentos celomaticos. A lo largo de las aéreas ambulacrales, las ramificaciones (pies) del sistema ambulacral son importantes para la locomoción, la captación del alimento, la respiración y la excreción. Muchas estrellas de mar son depredadoras, mientras que otras se 18 alimentan de pequeñas partículas. Los sexos son separados, y los sistemas reproductores muy simples. Las larvas, de vida libre y simetría bilateral, se fijan para transformarse en un individuo juvenil radial que posteriormente se libera y se convierte en una estrella adulta y libre. Los brazos de las ofiuras (clase Ofiuroideos) son delgados, y tienen un origen muy marcado en el disco central. No tienen pedicelarios ni ampollas, y sus surcos ambulacrales están cerrados. Sus pies ambulacrales carecen de ventosas, y su madreporito se encuentra en la cara oral. Reptan mediante el movimiento de sus brazos, mientras que utilizan sus pies ambulacrales para capturar alimento. Los osículos dérmicos de los erizos de mar (clase Equinoideos) son placas íntimamente unidas, que configuran un cuerpo compacto y sin brazos. Las aéreas ambulacrales están cerradas y se extienden hacia el polo aboral. Los erizos se mueven por medio de pies ambulacrales o mediante sus espinas. Algunos erizos (los dólares de arena y erizos acorazonados) están evolucionando hacia el regreso a la simetría bilateral. Los osículos dérmicos de los cohombros de mar (clase Holoturoideos) son muy pequeños, lo que se traduce en una pared del cuerpo blanda. Las holoturias están alargadas según el eje oral-aboral y descansan sobre un costado. Como ciertas aéreas ambulacrales son las que están siempre en contacto con el sustrato, las holoturias también están evolutivamente volviendo a la simetría bilateral. Los pies ambulacrales alrededor de la boca se han transformado en tentáculos, con los que se alimentan. Poseen un árbol respiratorio interno, y el madreporito cuelga libre en el celoma. Los lirios de mar (clase Crionoideos) son un enigmático grupo de afinidades inciertas con el resto de las clases de equinodermos. Tienen forma circular, pies ambulacrales marginales y dos canales anulares concéntricos en su sistema vascular acuífero. El antecesor de los equinodermos tenia simetría bilateral, y probablemente evoluciono a través de un estado sésil que desarrollo simetría radial y posteriormente dio lugar a las formas de vida libre. Los hemicordados son gusanos marinos que fueron considerados afines a los cordados porque se pensaba que su divertículo bucal era homologo de una notocorda. En común con los cordados, algunos poseen hendiduras branquiales y un cordón nervioso dorsal hueco. Las porciones en que se divide el cuerpo (probóscide, collar, tronco) poseen las típicas cavidades celomaticas (protocele, mesocele, metacele) de los deuterostomos. Los hemicordados de la clase Enteropneustos incluyen especies fosoras que se alimentan de partículas que filtran del agua por medio de las hendiduras branquiales. Los pterobranquios son tubícolas y se alimentan por filtración realizada con sus tentáculos. Los hemicordados, filogenéticamente, son importantes porque muestran afinidades con los cordados y los equinodermos. Junto con estos últimos, parecen ser el grupo hermano de los cordados. Capitulo 23: Los cordados (pg. 573) El filo Cordados recibe su nombre de la notocorda en forma de varilla que forma un eje corporal rígido en alguna etapa del ciclo vital de cada cordado. Todos los cordados comparten cinco características distintivas que los separan de cualquier otro filo: notocorda, cordón nervioso tubular dorsal, hendiduras branquiales faríngeas, endostilo y cola post-anal. Dos de los tres subfilos de cordados son invertebrados y carecen de cabeza bien desarrollada. Son los Urocordados (Tunicados), la mayoría de los cuales son sésiles cuando son adultos, pero que poseen todos un estado larvario nadador, y los Cefalocordados (lancetas), animales pisciformes que incluyen al famoso Anfioxo. 19 Se cree que los Cordados descienden de antecesores semejantes a hemicordados o equinodermos, probablemente en el periodo Precámbrico, pero el verdadero origen de los Cordados no se conoce todavía con certeza. En conjunto, los Cordados tienen una unidad fundamental de sistemas orgánicos y esquema corporal mayor que la que presentan muchos filos de invertebrados. El subfilo Vertebrados está formado por los miembros con huesos del reino animal (los mixines carecen en realidad de vertebras, pero se incluyen en los vertebrados por tradición, ya que comparten numerosas homologías). Se caracterizan como grupo por poseer una cabeza bien desarrollada, por su tamaño comparativamente grande, alto grado de movilidad y patrón corporal particular, lo que implica varias características distintivas que han conducido al enorme éxito del grupo. Las más importantes de estas características son: El endoesqueleto vivo, que permite el crecimiento continuo y proporciona un buen armazón para el anclaje y la acción eficaces de la musculatura; una faringe perforada por hendiduras branquiales (perdidas o muy modificadas en los vertebrados superiores) con un enorme incremento de la eficacia respiratoria; un tubo digestivo muscularizado, un corazón con cavidades, un riñón glomerular para satisfacer altas necesidades metabólicas y un sistema nervioso avanzado con un cerebro desarrollado y órganos sensoriales pares. La evolución de las mandíbulas y las extremidades pares probablemente contribuyo al increíble éxito de un grupo de vertebrados, los gnatostomados. Capitulo 24: Los peces (pg. 607) Los peces son vertebrados acuáticos poiquilotermos, con respiración branquial y con aletas. Son los vertebrados más antiguos, habiéndose originado a partir de un cordado antecesor desconocido en el cámbrico o posiblemente antes. Hay cinco clases de peces. Los mas primitivos son los mixines (Clase Mixines) y las lampreas (clase Cefalospidomorfos), con cuerpo anguiliforme, sin aletas pares; esqueleto cartilaginoso, una notocorda que persiste a lo largo de la vida y una boca en forma de disco adaptada para chupar o morder. El resto de los vertebrados tienen mandíbulas, un paso decisivo en la evolución de los vertebrados. Los miembros de la clase Condrictios (tiburones, rayas, torpedos y quimeras) son un grupo con gran éxito evolutivo, que tienen esqueleto cartilaginoso (característica regresiva), aletas pares, excelente equipamiento sensorial y característicos hábitos depredadores. Los peces óseos (clase Osteictios), pueden subdividirse en dos grupos. Los peces de aletas lobuladas de la clase Sarcopterigios, representados hoy por el celacanto y los peces pulmonados, constituyen un grupo parafiletico si se excluyen de él a los Tetrápodos, como ocurre en las clasificaciones tradicionales. El otro grupo son los peces de aletas con radios espinosos (clase Actinopterigios), un inmenso conjunto moderno que comprende casi todas las especies de peces marinos y de agua dulce más conocidos. Las modificaciones de los sistemas esquelético y muscular han mejorado la locomoción y la eficacia alimentaria. Los peces óseos modernos (peces teleósteos) se han diversificado en casi 23600 especies, que presentan una enorme cantidad de adaptaciones en la forma del cuerpo, el comportamiento y las preferencias ecológicas. Los peces nadan por contracciones ondulatorias de la musculatura, que generan un empuje (fuerza propulsora) y una fuerza lateral de sustentación. Los peces anguiliformes ondulan la totalidad del cuerpo, pero, en los nadadores más rápidos, las ondulaciones están limitadas a la región caudal, o solamente a la aleta caudal. La mayoría de los peces óseos pelágicos consiguen una flotación neutra para mantenerse en el 20 agua utilizando una vejiga natatoria rellena de gas, el mecanismo más eficaz de secreción de gas conocido en el reino animal. La sensibilidad a los sonidos puede mejorarse mediante los osículos de Weber, que transmiten los sonidos desde la vejiga natatoria hasta el oído interno. Las branquias de los peces tienen un flujo contracorriente eficaz entre el agua y la sangre, que les facilita unos elevados niveles de intercambio de oxígeno. Todos los peces, a excepción de los mixines, tienen una regulación osmótica e iónica bien desarrollada, conseguida principalmente por los riñones y las branquias. Con la excepción de los agnatos, sin mandíbulas, todos los peces tienen mandíbulas que están diversamente modificadas según los distintos hábitos de alimentación carnívora, herbívora, planctófaga y detritívora. La mayoría de los peces son, hasta un cierto punto, migradores y, algunos, como las anguilas de agua dulce y el salmón anádromo, hacen migraciones notables de gran amplitud y precisión. Los peces muestran una variedad extraordinaria de estrategias para la reproducción sexual. La mayoría son ovíparos, aunque los ovovivíparos y los vivíparos tampoco son raros. La estrategia reproductora puede ser, o bien producir gran número de gametos (y huevos) con baja supervivencia (muchos peces marinos), o poner pocos huevos con muchos cuidados por parte de los padres para una mayor supervivencia (peces de agua dulce. Capítulo 25: Los primeros tetrápodos y los anfibios modernos (pg. 630) Los anfibios son vertebrados cuadrúpedos primitivos y ectotérmicos, que poseen una piel glandular y respiración pulmonar, branquial o cutánea. Son los supervivientes de una de las dos grandes ramas de la filogenia de los tetrápodos; los representantes actuales de la otra rama son los amniotas. Los anfibios modernos se dividen en tres grupos evolutivos. Las cecilias (orden Gimnofiones) son un pequeño grupo tropical de formas alargadas y ápodas. Las salamandras (orden Urodelos) son anfibios con cola que han mantenido el arquetipo con cuatro patas de sus antecesores Paleozoicos. Las ranas y sapos (orden Anuros) son el mayor grupo de anfibios actuales y están especializados para la locomoción en tierra. La mayoría de los anfibios tienen un ciclo vital bifásico que comienza con una larva acuática; esta larva sufre una metamorfosis para dar lugar a un adulto terrestre que vuelve al agua para poner los huevos. Algunas ranas, salamandras y cecilias han conseguido un desarrollo directo que omite la etapa larvaria acuática, y algunas cecilias incluso presentan viviparismo. Las salamandras son únicas entre los anfibios por presentar varias especies perennibranquiadas, que retienen la morfología larvaria durante toda su vida, eliminando por completo la fase larvaria. La condición perennibranquiada es obligada en varias especies, pero en otras puede inducirse la metamorfosis a un estado terrestre mediante la desecación del hábitat acuático. Aunque los anfibios han desarrollado adaptaciones en la fase acuática de su ciclo, las correspondientes a su existencia terrestre son particularmente notables. El intercambio gaseoso para la respiración se produce a través de la piel en todos los anfibios y se complementa en muchos de ellos mediante pulmones. Sorprendentemente, las salamandras más estrictamente terrestres carecen de pulmones, mientras que otras formas acuáticas utilizan los pulmones como su principal mecanismo respiratorio. La vida en la tierra también requiere reforzar y rediseñar elementos esqueléticos, especialmente las costillas, las cinturas pectoral y pelviana y las extremidades. La percepción sensorial en el medio terrestre está favorecida por transformaciones derivadas de los sistemas auditivo y visual. A pesar de sus adaptaciones a la vida en tierra, los adultos y los huevos de todos los anfibios 21 necesitan ambientes frescos y húmedos, si no charcas, estanques o ríos. Los huevos y la piel de los adultos no tienen una protección eficaz ante condiciones de frio, calor o sequedad, lo que restringe en gran manera la radiación adaptativa de los anfibios a los ambientes con temperaturas moderadas y agua abundante. Capitulo 26: El origen de los amniotas y los grupos de reptiles (pg. 653) Los reptiles se separaron filogenéticamente de los anfibios laberinodontos durante el final del Paleozoico, hace 300 millones de años. Su éxito, como los primeros vertebrados verdaderamente terrestres, es atribuible al huevo amniótico, cuyas tres membranas extraembrionarias proporcionan al embrión la posibilidad de desarrollarse con la protección de una cascara. Por ello los reptiles pudieron poner huevos en tierra. Los reptiles también se diferencian de los anfibios por su piel seca y escamosa que impide la pérdida de agua, por sus mandíbulas más poderosas y por presentar fecundación interna, así como una serie de avances de los sistemas respiratorio, excretor y nervioso. Al igual que los anfibios, son ectotérmicos, pero muchos ejercen un considerable control etológico sobre su propia temperatura corporal. Antes del final del paleozoico, los amniotas sufrieron una radiación que los dividió en tres líneas: los anápsidos, que dieron lugar a las tortugas; los sinápsidos, una línea de reptiles mamiferoides que condujeron a los actuales mamíferos; y la línea de los diápsidos, de la que derivaron el resto de los reptiles y las aves. La gran radiación reptiliana del Mesozoico produjo una fauna cosmopolita de gran diversidad que incluía a los ictiosaurios, los plesiosaurios, los pterosauros y los dinosaurios, todos extintos. Las tortugas (Orden Testudines), con sus caparazones característicos, han cambiado poco su diseño desde el Triásico. Las tortugas son un pequeño grupo de longevas especies terrestres, semiacuáticas, acuáticas y marinas. Carecen de dientes. Son todas ovíparas y entierran sus huevos, incluso las marinas. Los lagartos, serpientes culebrillas ciegas (Orden Escamosos) comprenden el 95% de todos los reptiles vivientes. Los lagartos (Suborden Lacertilia) son un grupo diversificado y abundante adaptado para la marcha, la carrera, la escalada, la natación y la excavación. Se diferencian de las serpientes por presentar típicamente dos pares de patas (algunas especies son ápodas), ambas hemimandíbulas inferiores unidas, parpados móviles, oído externo y carencia de colmillos. Muchos lagartos están muy bien adaptados para sobrevivir en condiciones desérticas, áridas y calurosas. Las culebrillas ciegas (suborden Anfisbenios) son un grupo tropical poco numeroso, de formas ápodas hipogeas, adaptadas a la vida excavadora. Las serpientes (suborden Serpientes), además de ser totalmente ápodas, se caracterizan por sus cuerpos alargados y por un cráneo altamente cinético, que les permite tragar presas enteras, incluso mucho mas grandes que el diámetro de la propia serpiente. La mayor parte de las serpientes depende de sentidos químicos, especialmente os órganos de Jacobson, para cazar, más que de los poco desarrollados sentidos auditivo y visual. Dos grupos de serpientes (las boas y las víboras de foseta) tienen órganos especiales para la detección de infrarrojos que sirven para seguir a presas de sangre caliente. Muchas serpientes son venenosas. Los tuataras de Nueva Zelanda (orden Esfenodontos) es una especie relicta, y el único superviviente de un grupo que ha desaparecido hace 100 millones de años. Presentan varias características primitivas que son totalmente idénticas a las de los diápsidos fósiles del Mesozoico: Los cocodrilos y los caimanes (orden Crocodilios) son los únicos reptiles representantes de la 22 línea de los arcosaurios, que dio lugar a los dinosaurios, ya extintos, y a las aves actuales. Los cocodrilos tienen varias adaptaciones para una vida carnívora y semiacuática, lo que incluye un cráneo masivo con mandíbulas poderosas y un paladar secundario. Son los mayores reptiles vivientes y tienen la conducta social más compleja de todo el grupo. Capítulo 27: Las aves (pg. 684) Las mas de 9900 especies de aves vivientes son vertebrados endotérmicos ovíparos, cubiertos con plumas, que presentan las extremidades anteriores modificadas como alas. Están estrechamente relacionados común grupo de dinosaurios del Mesozoico con varios caracteres aviares, los terópodos. El ave fósil mas antigua que se conoce, Archaeopteryx, del período jurásico, tenía muchos rasgos reptilianos y era casi idéntico a ciertos dinosaurios terópodos excepto por la presencia de plumas. Probablemente debe considerarse como el grupo hermano de las aves actuales. Las adaptaciones de las aves para el vuelo son de dos tipos básicos: las que reducen peso corporal y las que producen mayor potencia para el vuelo. Las plumas, distintivo de las aves, son derivados complejos de las escamas reptilianas, y combinan ligereza con fuerza, impermeabilidad y alta capacidad aislante, El peso corporal se reduce, además por la eliminación de algunos huesos, fusión de otros (para proporcionar rigidez para el vuelo) y por la presencia en muchos huesos de espacios aéreo o cavidades neumáticas. El pico ligero y queratinizado, que reemplaza a las pesadas mandíbulas y dientes de los reptiles, sirve como mano y boca para muchas aves, y se ha adaptado de varias formas para los diferentes tipos de dieta. Las adaptaciones que proporcionan potencia para el vuelo incluyen una tasa metabólica alta y temperatura corporal acoplada a una dieta altamente energética; un sistema respiratorio muy eficaz, que consiste en un sistema de sacos aéreos dispuestos para producir un flujo constante y unidireccional de aire a través de los pulmones; músculos poderosos de vuelo y de las patas, dispuestos cerca del centro de gravedad del ave, y una circulación eficaz y de alta presión. Las aves tienen muy buena vista, buen oído y una extraordinaria coordinación para el vuelo. Los riñones metanéfricos producen acido úrico como principal producto nitrogenado. Las aves vuelan aplicando los mismos principios aerodinámicos que un aeroplano, y utilizando un equipo similar: alas para apoyo y elevación, y cola para giros y aterrizajes; también utilizan hendiduras alares para controlar el vuelo a bajas velocidades. La ausencia de vuelo en las aves es rara, pero ha evolucionado independientemente en varios órdenes, normalmente en islas donde los depredadores terrestres son escasos o faltan; todas ellas proceden de antecesores voladores. La migración de las aves implica movimientos regulares entre los cuarteles de verano y las regiones de invernada. La migración de primavera hacia el Norte, donde existe más comida disponible para los polluelos, permite mayor éxito reproductor. Son utilizados muchos tipos de señales para orientarse durante la migración, incluyendo el sentido innato para la dirección y la capacidad de navegar utilizando el Sol, las estrellas y el campo magnético terrestre. La conducta social altamente desarrollada de las aves se manifiesta en una serie de vívidos cortejos, selección de compañeros de apareamiento, comportamiento territorial, incubación y cuidado de los polluelos. Capítulo 28: Los mamíferos (pg. 714) 23 Los mamíferos son vertebrados homeotérmicos y endotérmicos, con el cuerpo cubierto de pelo, que alimentan a sus crías con leche. Las aproximadamente 4800 especies descienden del linaje sinápsido de reptiles mamiferoides que aparecieron en el período Carbonífero de la Era Paleozoica. Su evolución se puede seguir desde los Pelicosaurios del periodo Pérmico hasta los Terápsidos de los periodos Pérmico y Triásico de la era Mesozoica. Un grupo de terápsidos, los cinodontos, dieron lugar durante el Triásico a los Terios, los verdaderos mamíferos. La evolución de los mamíferos se ha visto acompañada por la aparición de muchos caracteres derivados de gran importancia, como un cerebro mayor, con más integración sensorial, tasas metabólicas altas, endotermia y muchos cambios en el esqueleto que permitieron una vida más activa. Los mamíferos se diversificaron rápidamente durante el periodo Terciario de la era Cenozoica. Los mamíferos reciben su nombre de los órganos glandulares secretores de leche de la hembra (rudimentarios en el macho), una adaptación única que, combinada con un prolongado cuidado de la prole, asegura a los hijos la provisión de alimento y facilita la transición al estado adulto. El pelo, una diferenciación tegumentaria que cubre el cuerpo de muchos mamíferos, sirve de variadas formas como protección mecánica, aislamiento térmico, coloración protectora e impermeabilización. La piel de los mamíferos es rica en glándulas sudoríparas, que funcionan en el enfriamiento por evaporación; las odoríferas, utilizadas en la conducta social, y las sebáceas, que segregan aceites lubricantes para la piel. Todos los mamíferos placentarios presentan dientes deciduos que son reemplazados por dientes permanentes (dentición difiodonta). Los cuatro grupos de dientes, incisivos, caninos, premolares y molares, pueden estar altamente modificados en diferentes mamíferos para las distintas tareas de cortar, triturar y desgarrar alimentos, pudiendo incluso faltar. Los hábitos de alimentación de los mamíferos influyen enormemente en su forma corporal y su fisiología. Los insectívoros tienen dientes puntiagudos para perforar el exoesqueleto de los insectos y otros pequeños invertebrados. Los mamíferos herbívoros tienen dientes especializados para triturar la celulosa y las plantas ricas en sílice, y presentan cámaras especiales para almacenar microflora intestinal, que descompone la celulosa de la dieta vegetal. Los mamíferos carnívoros tienen adaptaciones como los dientes y la musculatura de las mandíbulas, para matar y procesar su alimento, principalmente mamíferos herbívoros. Los omnívoros se alimentan tanto de plantas como de animales, y presentan diversos tipos de dientes. Algunos mamíferos marinos, terrestres y aéreos migran; algunas migraciones, como las de las focas o los renos, son amplias. Las migraciones son generalmente hacia climas favorables y condiciones alimentarias óptimas o para aproximar ambos sexos para el apareamiento. Los mamíferos con vuelo verdadero, los quirópteros, son nocturnos y, por tanto, evitan la competencia directa con las aves. La mayor parte de ellos emplean ecolocalización ultrasónica para volar y alimentarse en total oscuridad. Los mamíferos vivientes más primitivos son los monotremas de la región australiana, que ponen huevos. Tras el nacimiento, las crías se alimentan con leche de la madre. Todos los demás mamíferos son vivíparos. Los marsupiales tienen periodos de gestación breves, por lo que los embriones nacen subdesarrollados y completan su crecimiento en el marsupio de la madre, alimentándose con leche. Los mamíferos restantes son euterios, mamíferos que desarrollan una unión placentaria evolucionada entre la madre y los embriones, a través de la cual estos pueden alimentarse durante largo tiempo. 24 Las poblaciones de los mamíferos fluctúan por razones tanto dependientes como independientes de la densidad, y algunos mamíferos, particularmente los roedores, pueden experimentar ciclos de extrema abundancia en la densidad de sus poblaciones. El éxito indiscutible de los mamíferos como grupo no puede ser atribuido a una perfección de los sistemas orgánicos, sino a su impresionante adaptabilidad de conjunto, la capacidad para engranarse de manera más perfecta en el conjunto organizado de las condiciones ambientales, y, de este modo, explotar prácticamente todos los hábitats de la Tierra. Los principios evolutivos darwinistas nos proporcionan una buena perspectiva de nuestros propios orígenes. El hombre es un primate, un grupo de mamíferos que descienden de un antecesor parecido a una musaraña. El ancestro común de todos los primates actuales era arborícola, y tenía dedos prensiles y ojos frontales capaces de visión binocular. Los primates se diversificaron durante los últimos 80 millones de años, para formar dos grandes líneas de descendencia: los prosimios (lémures, lorises y tarseros) y los simios (monos y homínidos). Los primeros homínidos aparecieron en África hace unos 7 millones de años, y dieron lugar a diversas especies de australopitecinos, que pervivieron durante más de 4 millones de años. Estos dieron lugar a la especie Homo habilis, el primer fabricante de herramientas de piedra, y convivieron con él. Homo erectus apareció hace 1,8 millones de años y fue sustituido eventualmente por el hombre moderno. 25
 0
0
Anuncio
Documentos relacionados



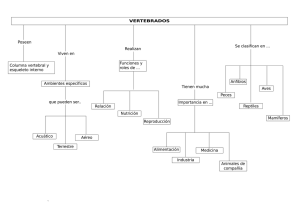
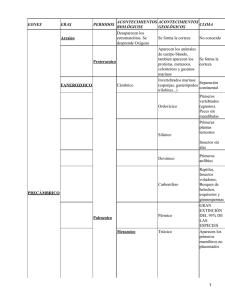





Descargar
Anuncio
Añadir este documento a la recogida (s)
Puede agregar este documento a su colección de estudio (s)
Iniciar sesión Disponible sólo para usuarios autorizadosAñadir a este documento guardado
Puede agregar este documento a su lista guardada
Iniciar sesión Disponible sólo para usuarios autorizados