Pincha aquí si no puedes visualizar el PDF
Anuncio

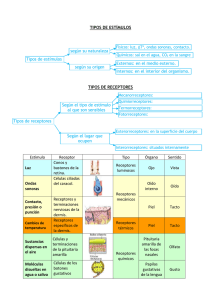
RESUMEN TEMA 11: LOS SISTEMAS SENSORIALES 1.-INTRODUCCCIÓN A LA FISIOLOGIA DE LOS SENTIDOS No cabe duda de que la información es poder. Los seres vivos recibimos información del exterior, y la utilizamos para sobrevivir. Llamamos a toda fuente de energía susceptible de influir en la conducta de un organismo: estímulo sensorial y a los sistemas a través de los cuales el organismo capta la información, sistemas sensoriales o sentidos. La selección natural ha ido perfeccionando el diseño de estos sistemas a fin de que capten diferentes tipos de energía relevantes para la supervivencia. Como sabemos por el capitulo anterior, a semejanza de lo que sucede en un ordenador, en un cierto nivel, el neuronal, toda la información, cualquiera que sea su naturaleza es codificada en forma de señales eléctricas y/o químicas. Para que un sonido, un color un estimulo doloroso o un sabor, puedan influir sobre la conducta de un organismo antes han tenido que ser transformados en cambios eléctricos neuronales, especialmente en impulsos nerviosos. En este capítulo emprenderemos la apasionante tarea de descubrir cómo pueden nuestros sistemas sensoriales “traducir” los distintos tipos de energía: luz, sonido, presión, calor y transformarla en impulsos nerviosos. ¿De qué modo filtramos la información? ¿Cómo la procesamos? ¿De qué manera son adaptativos nuestros sentidos? Iniciamos aquí nuestro viaje al familiar y al mismo tiempo ignoto mundo de nuestros sentidos. 1.1.-TIPOS DE ENERGÍA ESTIMULAR: MODALIDADES SENSORIALES Sucede una cosa muy curiosa con el mundo que llamamos real, y es que para determinar “que hay allí fuera” tenemos que tener claro que dependerá de qué sea relevante para quien esté mirando. El pez perro captura a sus presas gracias a que puede detectar las mínimas distorsiones en el campo eléctrico que ellas producen. Si encierras a un murciélago en una habitación oscura puedes estar seguro que no va a chocar con ningún objeto gracias a su espectacular sentido de la audición. Un cocodrilo es capaz de oler el miedo y nosotros, los primates, tenemos una excelente visión. A la transformación de las diferentes modalidades energéticas y químicas (luz, sonido, tacto, olor, sabor) en actividades eléctricas, que llevan a cabo los receptores sensoriales, se le llama transducccion sensorial. En esta tarea nuestro S.N se enfrenta a un problema, y no pequeño: el hecho de que todos los impulsos nerviosos son iguales. Cuantitativa y cualitativamente un potencial de acción producido en el nervio óptico es igual que uno desencadenado en el nervio auditivo o en el bulbo olfatorio. Entonces, ¿Cómo es posible que los organismos respondamos adecuadamente a los estímulos, siendo como son los impulsos nerviosos todos iguales? La explicación a este enigma viene propuesta por la llamada ley de las líneas marcadas: la cual hace referencia a que las vías sensoriales siguen un trayecto predeterminado y genéticamente programado, desde los receptores a las áreas de proyección sensorial, donde se “interpreta” o descodifica la información recibida. Es decir que la modalidad sensorial depende de la zona del SNC a la que llegan los impulsos nerviosos y no de cuál haya sido la energía estimular. Por ejemplo si estimulamos con calor los receptores cutáneos que informan de la sensación de frio sentiremos frio. Pero no hay por qué preocuparse: en la naturaleza suele ser muy difícil estimular vías sensoriales con estímulos no apropiados. En mayor o menor medida aquello que recibimos por medio de los sentidos tiene que ver con lo que realmente hay “allí fuera” ¡si no fuera así no estaríamos aquí para contarlo! Lo que sucede es que cada receptor está programado para responder a un estrecho margen de valores de un tipo determinado de energía: el estímulo adecuado 1.2.-CUANTIFICACIÓN DE LA ENERGÍA ESTIMULAR: AMPLITUD DEL ESTIMULO Nuestra experiencia nos dice que somos capaces de percibir unas sensaciones como más intensas que otras. Dos son los mecanismos que usan nuestras neuronas para informarnos de esta intensidad diferencial: -El código de frecuencia: Este mecanismo consiste en que a medida que aumenta la intensidad de la estimulación, aumenta correlativamente la frecuencia de potenciales de acción que una neurona sensorial transmite. Ahora bien, hay un valor máximo (debido al periodo refractario del que hablamos en el capítulo anterior).Una neurona no puede generar más de unos 1.000 potenciales de acción por segundo. Cuando una determinada intensidad de estimulación le haga responder con esa frecuencia de disparo, cualquier aumento de esa intensidad pasará desapercibido para esa neurona en concreto. Por suerte, los valores mínimos si son diferentes ya que cada neurona sensorial tiene un determinado umbral de respuesta y mientras unas responden a estimulaciones muy débiles (son muy sensibles) otras sólo lo hacen cuando la estimulación es intensa o muy intensa. - Fraccionamiento según el rango o código poblacional: Este segundo mecanismo utiliza las diferencias en el umbral de disparo: a medida que aumenta la intensidad de la estimulación, las neuronas con un umbral de disparo más alto empiezan a disparar potenciales de acción. El S.N ante este hecho, interpreta que está subiendo la intensidad del estímulo 1.3.-CUANTIFICACIÓN DE LA ENERGÍA ESTIMULAR: DURACIÓN DEL ESTIMULO Un fenómeno bastante habitual es el de la disminución o incluso desaparición de la sensación aun cuando la estimulación sensorial sigue teniendo la misma intensidad. Esta reducción puede explicarse bien por la fisiología de los receptores sensoriales (adaptación sensorial) o bien por qué hay un aprendizaje (no asociativo) u otros procesos más complejos que generan cambios fisiológicos en las sinapsis entre neuronas dentro del SNC (habituación) Si pensamos en un aparato de medida como un termómetro, por ejemplo, vemos que transmite la misma información ante el mismo cambio en la energía estimular: la temperatura sube un grado tanto de 0 a 1 como de 100 a 101. Nuestros receptores sensoriales no funcionan exactamente así, no todos tiene el mismo grado de fidelidad. Por ejemplo los receptores de presión táctil asociados a los corpúsculos de Pacini, que son a diferencia de otros, receptores neuronales, sólo responden con potenciales de acción al principio y al final de la estimulación y dejan de hacerlo el resto del tiempo, es decir que responden únicamente a los cambios en el estímulo (aquí vemos un ejemplo de adaptación sensorial). Por supuesto no todos los receptores se adaptan de la misma forma, algunos lo hacen muy rápidamente y otros no lo hacen en absoluto. Los receptores que se adaptan deprisa reciben el nombre de receptores fásicos y están enfocados a detectar los cambios, (pensemos que rápido nos adaptamos cuando entramos a una habitación oscura con respecto a la luz que había en el exterior). A los receptores que tardan mucho o no se adaptan se les denomina receptores tónicos (si tenemos un fuerte dolor de muelas no nos adaptaremos tan rápidamente sin embargo) 1.4.-LOCALIZACIÓN DE LA ENERGÍA ESTIMULAR: CAMPOS RECEPTIVOS E INHIBICIÓN LATERAL Por su parte la localización del estímulo se basa en la distribución y densidad de los receptores y en el modo en que estos se relacionan con las diferentes zonas centrales de procesamiento. Esto explica que en algunas zonas dos estímulos muy próximos se sientan como uno solo mientras en otras se distinguen perfectamente lo cual se debe a la diferente densidad de receptores (por ejemplo en los dedos de la mano hay cuatro veces más receptores que en la palma). En el caso de los receptores cutáneos, a la corteza de proyección sensorial se la conoce como humunculo sensorial, que contiene un mapa, algo caricaturesco, de toda la superficie corporal que demuestra la correspondencia entre cada zona de la piel y su respectiva zona de recepción cortical, si bien no todas están representadas con la misma intensidad. Lo mismo sucede con otras modalidades sensoriales, cada una de las cuales posee su propio mapa (recordemos la ley de líneas marcadas) La actividad fisiológica de una neurona sensorial resulta modificada por la estimulación de los receptores que contactan con ella, es lo que se conoce como campo receptivo. Hay que tener en cuenta que el campo receptivo no incluye solo a las neuronas sensoriales y a sus receptores vinculados, sino que también se habla de campo receptivo de neuronas de relevo sensorial de segundo y tercer orden. Esto hace que los campos receptivos táctiles o por ejemplo de las células ganglionares de la retina sean muy complejos, ya que además de los receptores sensoriales incluyen neuronas inhibitorias asociadas a receptores próximos. Un ejemplo de la interacción entre campos receptivos próximos es lo que se conoce como inhibición lateral. Determinados animales, como armiños zorros u osos polares poseen un pelaje blanco que hace que se fundan con el paisaje nevado y de ese modo puedan ver sin ser vistos. Observemos esta figura: Si no fijamos con atención podemos ver una estrecha banda más obscura y otra más clara cerca de los bordes de contacto entre las bandas más anchas. Las bandas de Mach son una ilusión óptica que parte de una imagen con dos bandas, una iluminada y la otra oscura, separadas por una estrecha banda central coloreada con un gradiente de iluminado a oscuro. El ojo humano percibe dos estrechas bandas de diferente luminosidad, que no están presentes en la imagen verdadera, a cada lado del gradiente. Esta ilusión se denomina así debido a Ernst Mach. La observación de los omatidios o receptores visuales de los cangrejos en herradura nos permitió comprobar que la estimulación de un campo receptivo no solo excita a la neurona sensorial sino que inhibe a las que tienen campos receptivos próximos, siendo en esa línea de contacto entre bandas donde se manifiesta el efecto de esta inhibición lateral. La codificación de la intensidad de un estímulo es el resultado de la interacción de ese estímulo y demás estímulos que puedan estar incidiendo incluyendo neuronas inhibitorias, de segundo tercer orden etc…No cabe duda de que la selección natural ha favorecido sistemas sensoriales capaces de detectar cambios. En este sentido la inhibición lateral no es más que un sistema de amplificación. 2.-LOS RECEPTORES SENSORIALES Y LAS TRANSDUCCIÓN SENSORIAL Entre los vertebrados los receptores sensoriales pueden ser: NEURONALES -Olfatorios -Cutáneos -Propioceptivos e interoceptivos CÉLULAS ESPECIALIZADAS -Vista -Oído -Equilibrio -Gusto Por otra parte los receptores también pueden clasificarse en exteroreceptores (vista, oído, olfato, gusto y tacto), interoreceptores, que informan de los cambios en las constantes fisiológicas del organismo, glucosa, oxigeno, CO2, sales…o propioceptores que proporcionan información sobre el estado músculo-esquelético y la posición corporal. En cualquiera de los casos el estímulo produce cambios de tipo graduado (similares a los potenciales postsinápticos) en el potencial de membrana del receptor, que se denominan potencial de receptor o potencial generador si el receptor es una neurona. Normalmente este potencial de receptor es una despolarización (exceptuando los receptores visuales donde se producen hiperpolarizaciones). En cualquier caso, de manera directa o indirecta para que el impulso se transmita es preciso que los cambios de potencial en los receptores generadores alcancen el umbral de excitación en las neuronas sensoriales y se produzca un potencial de acción (en el caso de las neuronas ganglionares visuales, que están siempre disparando, lo que se producen son cambios en la frecuencia de disparo). 2.1.-QUIMIORECEPTORES Y TRANSDUCCIÓN 2.1.1. Receptores gustativos Los receptores del gusto se encuentran en la lengua pero también en el paladar, laringe, faringe, esófago y epiglotis. En los vertebrados, estás células receptoras se insertan en el botón gustativo. Estos botones se concentran en las papilas gustativas, que a su vez poseen distinta distribución y morfología Las células gustativas propiamente dichas proyectan vellosidades hasta el poro gustativo en que termina el botón gustativo y en el otro extremo hacen sinapsis con las neuronas sensoriales (figura 11.7). A las cuatro cualidades gustativas conocidas: dulce, salado, ácido y amargo, cabría añadir el glutamato monosódico (el unami de los orientales). Está claro que la selección natural ha potenciado el desarrollo del sentido del gusto porque para un organismo conocer que sustancias le nutren y cuales le hacen daño es un asunto de primerísima necesidad. Que sintamos predilección por alimentos como el chocolate o las patatas fritas, no es fruto únicamente del gusto personal, sino que alguna vez ha resultado adaptativo tener una reserva de energía. Pero, ¿qué pasa en la boca cuando comemos?, ¿cómo se produce la transducción?, el mecanismo de los quimioreceptores gustativos no es muy diferente del de los quimioreceptores bacterianos: al interactuar diferentes tipos de moléculas orgánicas con las microvellosidades, se desencadenan cambios en la permeabilidad de la membrana a determinados iones. Los receptores gustativos funcionan de manera similar a las neuronas en el sentido de que las sustancias estimulantes actúan sobre los receptores de membrana, bien abriendo canales iónicos bien mediante la intermediación de segundos mensajeros (diferentes según el tipo de sabor). En cualquiera de los casos se produce una despolarización debida a la apertura o cierre de los canales de Na+, K+ o Ca2, que es de tipo graduado, generando la liberación de un neurotransmisor químico en la sinapsis entre el receptor y la neurona sensorial (figura 11.9) 2.1.2. Receptores olfatorios En las especies que carecen de lenguaje hablado, la comunicación con fines sexuales suele iniciarse por vía olfativa. En el caso de la especie humana la función del olfato es algo más imprecisa y sutil (aparte de que no somos una especie que nos caractericemos por un especial desarrollo de este sentido) sin embargo podemos detectar, identificar y discriminar más de 10.000 olores diferentes. Son las sustancias orgánicas, casi exclusivamente las que pueden producir estimulación olorosa. Ácidos, alcoholes, ésteres, aromas florales y aceites esenciales sirven a plantas y animales en el establecimiento de códigos olfativos. Especialmente importantes son las sustancias que informan de la especie y el estado hormonal de los individuos (feromonas, almizcle…) En el hombre, las neuronas receptoras olfatorias o receptores olfatorios, se encuentran en la mucosa olfatoria, situada en la parte superior de la cavidad nasal. Respirando normalmente, entre un 5% y un 10 % de aire pasa cerca de la mucosa. Los receptores olfatorios son neuronas bipolares con una única proyección dendrítica acabada en cilios (ramificaciones) que se dirigen hacia la superficie de la mucosa y una proyección axónica en el lado opuesto, qué, atravesando el hueso craneal conecta ipsilateralmente con el bulbo olfatorio. A diferencia de la mayor parte de neuronas, los receptores olfatorios se regeneran por diferenciación de células basales cada 60 días. A diferencia del gusto, no ha sido posible determinar los olores básicos, es decir, la estructura molecular que determina las sensaciones olfativas básicas. Se han propuesto 7 aromas primarios de los que por combinación se derivarían los demás: etéreo, frutal, floral, canforáceo, menta, pútrido y acre. Los receptores olfatorios, al interactuar con los estímulos apropiados, generan potenciales de acción, que se difunden pasivamente por el soma celular hasta alcanzar el cono axónico donde se genera un potencial de acción cada vez que se alcanza el umbral de disparo. Algunos de estos disparos parecen ser consecuencia de la apertura de canales de Na+ mediada por el segundo mensajero AMPc, en otros casos se produce la apertura de canales de Ca2. La transducción olfatoria se inicia en los cilios de las neuronas receptoras. La frecuencia de disparo de potenciales de acción depende de la concentración de la sustancia olorosa. Sin embargo, ante una exposición prolongada, se produce adaptación sensorial, idéntica a la que ocurre en los receptores gustativos. 2.1.3. Receptores nocioceptivos Ahora nos centraremos en el proceso de transducción de la información nociceotiva (es decir en como determinados procesos químicos afectan a los receptores sensoriales que informan sobre el dolor) dejando de lado, por el momento todo lo relativo al dolor cuya transducción se lleva a cabo por otros mecanismos (mecanoreceptores, termoreceptores….) Los nociceptores son terminaciones nerviosas libres de neuronas sensitivas primarias, cuyos cuerpos neuronales se encuentran en los ganglios raquídeos. Esto quiere decir que los nociceptores no están rodeados de estructuras especiales, como es el caso de otros receptores sensoriales de la piel, como los corpúsculos de Pacini que detectan las vibraciones, o los discos de Merkel, que detectan la presión. Hay tres grandes clases de nociceptores: térmicos, mecánicos y polimodales. Todos ellos tienen en común la existencia de umbrales de excitación elevados, en comparación con otros receptores como los del tacto y la temperatura. Esto implica que normalmente no se activan en ausencia de estimulaciones nocivas. Dado que se trata de terminaciones nerviosas sin estructuras especiales, el término nociceptor se refiere tanto a la fibra nerviosa aferente como a su receptor. Los nociceptores se encuentran en muchos tejidos corporales como la piel, vísceras, vasos sanguíneos, músculo, tejido conectivo, periostio y meninges. Son muchísimas las sustancias, tanto exógenas como endógenas, que afectan a las terminaciones sensoriales nocioceptivas. Algunas de estas sustancias actúan directamente sobre las neuronas sensoriales, otras actúan como moduladoras modificando el umbral de respuesta de los terminales libres. Entre las sustancias endógenas se encuentran las prostaglandinas, la serotonina, la histamina, la acetilcolina, aminoácidos, óxido nítrico, opiodes etc…Muchas de estas sustancias son liberadas como parte del proceso de lesión/inflamación en respuesta a un estímulo doloroso y o bien producen o bien modulan el proceso de transducción generando potenciales de acción en estas neuronas sensoriales. Sabiendo esto podemos comprender como funcionan algunos fármacos, como los antihistamínicos. Como ejemplo curioso está la capsaicina, que da el sabor picante a la guindilla, de modo que al comer una guindilla no nos pica ¡nos duele! 2.2.-MECANORECEPTORES Y TRANSDUCCIÓN 2.2.1. Receptores cutáneos Un mecanorreceptor es un receptor sensorial que reacciona ante la presión mecánica o las distorsiones. Existen cuatro tipos principales en la piel humana: los corpúsculos de Pacini, los corpúsculos de Meissner, las terminaciones nerviosas de Merkel y los Corpúsculos de Ruffini. A modo de ejemplo, y porque son de gran tamaño, por lo que mucha información de las propiedades de los mecanoreceptores se ha obtenido a partir de ellos, vamos a describir la fisiología de la transducción de los corpúsculos de pacini Los Corpúsculos de Pacini poseen una cápsula compuesta por numerosas capas de células de tejido conectivo (a semejanza de una cebolla) que rodean la fibra nerviosa. Responden muy deprisa a cambios en la presión y tienden a reunirse cerca de las articulaciones, en algunos tejidos profundos, así como en las glándulas genitales y mamarias. Informan al cerebro qué algo está ejerciendo presión. Sin embargo aunque responden de forma intensa unos segundos al inicio de la presión, en seguida se produce adaptación sensorial y no responden de nuevo hasta que cesa la presión, se dice por tanto que responden a toques transitorios, en lugar de presión sostenida. Esto es porque un corpúsculo de Pacini es capaz de adaptarse rápidamente de manera que informa de cuando se produce el estímulo y cuando desaparece. A este tipo de receptores se les denomina receptores fásicos. En las zonas de la piel más sensibles al tacto, como las yemas de los dedos o la lengua, el número de corpúsculos de Pacini es mayor que en áreas menos sensibles, sus campos receptivos son pequeños y existe una elevada densidad de inervación. Es decir hay más neuronas en el S.N.C. que procesan la información procedente de las yemas de los dedos en comparación con la procedente de otras zonas. La tranducción de la energía mecánica en potenciales generadores por parte de los corpúsculos de Pacini sigue los siguientes pasos: 1-El estimulo que incide sobre la piel es modificado y transmitido por las células adyacentes 2-Posteriormente incide sobre la membrana de la neurona sensorial para producir un cambio en su potencial de membrana. Si la membrana es deformada por esta estimulación mecánica los canales mecanosensibles al Na+ y al K+ se abren (debido a que están unidos al citoesqueleto de la célula) provocando la despolarización del canal axónico (ver figura 11.16) 3- Si la despolarización es suficiente para alcanzar el umbral, se genera un potencial de acción. 2.2.2. Receptores auditivos El oído conforma los órganos de equilibrio y audición. También se le denomina órgano vestibulococlear. El oído es un órgano que se encuentra muy desarrollado, principalmente en mamíferos como los félidos y los grandes cetáceos y de manera espectacular en los murciélagos, en los cuales cumple una misión semejante a la de la visión en nuestra especie. En el caso del ser humano esta evolución del sentido auditivo no está tan desarrollada. Originariamente el oído era una estructura especializada en recoger las vibraciones del medio líquido, pero posteriormente algunos órganos como el tímpano y la cadena de huesecillos se adaptaron para amplificar las ondas sonoras aéreas (ya que las vibraciones del aire por si solas no son capaces de afectar al liquido coclear, porque se reflejan en gran medida en los medios líquidos). Quizá nos parezca algo extraño pero el sonido es en realidad un fenómeno mecánico de naturaleza vibratoria. Las moléculas de aire vibrante transmiten ondas mecánicas, que son lo que el cerebro interpreta como sonidos. El proceso de audición podemos dividirlo en dos etapas: 1. la fisiológica, de la que se encarga el sistema auditivo períferico. 2. la psicológica (percepción) de la que se encarga el sistema auditivo central. El Sistema auditivo periférico es el responsable de los procesos fisiológicos de la audición. Estos procesos que permiten captar el sonido y transformarlo en impulsos eléctricos susceptibles de ser enviados al cerebro a través de los nervios auditivos. El sistema auditivo periférico lo constituye el oído. El oído humano se divide en tres partes: 1. el oído externo, que canaliza la energía acústica. 2. el oído medio, que transforma la energía acústica en energía mecánica, transmitiéndola - y amplificándola- hasta el oído interno. 3. el oído interno, donde se realiza la definitiva transformación de la energía mecánica en impulsos eléctricos. Cuando el sonido llega al oído, las ondas sonoras son recogidas por el pabellón auricular (o aurícula). El pabellón auricular, por su forma helicoidal, funciona como una especie de "embudo" que ayuda a dirigir el sonido hacia el interior del oído. Una vez que ha sido recogido el sonido, las vibraciones provocadas por la variación de presión del aire cruzan el canal auditivo externo y llegan a la membrana del tímpano, ya en el oído medio, golpeando esta membrana como un tambor. En el oído medio, se produce la transformación la energía acústica en energía mecánica. En este sentido, el oído medio es un transductor mecánico-acústico. Además de transformar la señal, antes de que ésta llegue al oído interno, el oído medio la habrá amplificado. La presión de las ondas sonoras hace que el tímpano vibre empujando a los osículos (martillo, yunque y estribo) que, a su vez, transmiten el movimiento del tímpano al oído interno. Cada osículo empuja a su adyacente y, finalmente el movimiento se trasmite a la ventana oval. Es un proceso mecánico, el pie del estribo empuja a la ventana oval, ya en el oído interno. Esta fuerza empuja a la ventana oval es unas 20 veces mayor que la que empujaba a la membrana del tímpano, lo que se debe a la diferencia de tamaño entre ambas. Esta presión ejercida sobre la ventana oval, gracias a la helicotrema (pequeña abertura cerca del ápice de la cóclea mediante la que se comunican las rampas vestibular y timpánica ) penetra en el interior de la cóclea (caracol) y pone en movimiento el líquido linfático que ésta contiene. En la coclea hay tres canales: las rampas vestibular y timpánica, rellenas de perilinfa y entre ellas la rampa media o conducto coclear, relleno de endolinfa. El líquido linfático se mueve como una especie de ola y, transmite las vibraciones a las dos membranas que conforman la cóclea (membrana tectorial, la superior, y la membrana basilar, la inferior). Entre ambas membranas se encuentra el órgano de Corti, que es el transductor propiamente dicho. En el órgano de Corti se encuentran las células receptoras llamadas células ciliadas Existen aproximadamente 24 000 de estas fibras pilosas, dispuestas en 4 largas filas que son las que recogen la vibración de la membrana basilar. Como la membrana basilar varía en masa y rigidez a lo largo de su longitud su frecuencia de resonancia no es la misma en todos los puntos: En el extremo más próximo a la ventana oval y al tímpano, la membrana es rígida y ligera, por lo que su frecuencia de resonancia es alta. Por el contrario, en el extremo más distante, la membrana basilar es pesada y suave, con lo que su resonancia es de baja frecuencia. Veamos ahora paso a paso el proceso de transducción: 1.-la membrana basilar se curva como consecuencia de la presión que ejerce sobre ella la perilinfa 2.-Los esterocilios (células ciliadas de la membrana tectorial) se inclinan lo que provoca cambios en la conductancia iónica abriéndose los canales de K+ (todo esto se comprende perfectamente mirando la figura 11.20) 3.-La endolinfa posee mucha mayor concentración de K+ que la perilinfa (entre ellas hay una diferencia de -80mv, el llamado potencial endocloquear) y más que en el interior celular. Cuando se abren los canales el K+ entra movilizado por el flujo a favor del gradiente. La despolarización activa los canales de Ca2+ que entran produciéndose una hiperpolarización. 4.- En los laterales de las células ciliadas se encuentran canales de K+ sensibles al Ca2+. En determinado momento se abren estos canales y se produce una repolarización 5.-La células ciliadas más internas se despolarizan e hiperpolarizan de manera diferente, ya que más al interior hay menos esterocilios y estos no están en contacto con la membrana tectorial. Lo más probable es que sus cilios se vean afectados principalmente pos la endolinfa 6.-Las células ciliadas hacen sinapsis con las neuronas del nervio auditivo. Parece ser que liberan glutamato, responsable de la despolarización de las neuronas auditivas Aunque su función es muy distinta, los receptores sensoriales del sistema vestibular o del equilibrio, son igualmente células ciliadas que se despolarizan o hiperpolarizan por la presión de la endolinfa al desplazarse esta por los canales semicirculares. 3.-FOTORECEPTORES Y TRANSDUCCIÓN VISUAL El ojo es el órgano de la visión en los seres humanos y en los animales. Los ojos de las diferentes especies varían desde las estructuras más simples, capaces de diferenciar sólo entre la luz y la oscuridad, hasta los órganos complejos que presentan los seres humanos y otros mamíferos, que pueden distinguir variaciones muy pequeñas de forma, color, luminosidad y distancia. En realidad, el órgano que efectúa el proceso de la visión es el cerebro; la función del ojo es traducir las vibraciones electromagnéticas de la luz en un determinado tipo de impulsos nerviosos que se transmiten al cerebro Para alcanzar la Retina, un haz de luz debe atravesar los medios refringentes del ojo humano, que son cuatro: 1. La Córnea. 2. El Humor acuoso. 3. La lente o cristalino. 4. El humor vítreo. La superficie curva de la retina es muy importante, ya que permite compensar el cambio de trayectoria que sufre un haz luminoso, al pasar por medios con índices de difracción tan distintos. Ya dentro del ojo, los índices de refracción del cristalino, humor acuoso y humor vítreo, son tan parecidos, que el haz de luz no sufre mucho cambio, sólo la inversión que mencionamos anteriormente Además de los elementos nombrados anteriormente, el ojo presenta una estructura no refringente muy importante, cuya función es la de regular la entrada de luz. Nos referimos al Iris, parte pigmentada a la cual le debemos el color de nuestros ojos. Esta posee fibras musculares que de forma involuntaria se contraen o expanden, limitando o facilitando la incidencia lumínica al órgano. El Iris regula el área de la pupila que quedará expuesta a la radiación luminosa Para permitir que la luz llegue a las células fotorreceptoras de la retina sin ser dispersada o absorbida las capas celulares de la retina más próximas al cristalino no tienen mielina y son relativamente transparentes Además La retina tiene una pequeña invaginación en forma de surco, situada aproximadamente, en su punto central denominada fóvea en la que los cuerpos neuronales de las células retinianas se disponen desplazados hacia los laterales para dejar paso libre a la luz con el fin de que no haya distorsión y la luz llegue con menos dificultad En la parte central de la fóvea, llamada foveola, prácticamente sólo hay células fotorreceptoras en alta concentración que permite que la agudeza visual sea máxima allí. Ahora veamos cómo se produce la transducción. Existen 2 tipos de células fotorreceptoras llamadas conos y bastones (en los humanos hay unas 20 veces más cantidad de bastones que conos) Estos fotorreceptores al ser estimulados por la luz responden con cambios en el potencial de membrana, (ojo que no producen directamente potenciales de acción). En ambos fotorreceptores existen moléculas sensibles a la luz llamadas pigmentos visuales o fotopigmentos. Los bastones contienen un tipo de pigmento específico, mientras que hay 3 tipos de pigmentos en los conos que determinan 3 tipos de conos según el pigmento que usan El pigmento visual de los bastones es más sensible a la luz que los de los conos. Un único fotón puede producir una señal eléctrica detectable en los bastones, mientras que se necesitan cientos para que un cono pueda generar la misma respuesta. Los bastones están especializados en la visión en condiciones de baja luminosidad. Presentan una elevada sensibilidad a la luz aunque se saturan en condiciones de mucha luz y no detectan los colores. Los conos, por su parte, están especializados en la visión diurna y son son responsables de la visión en colores porque como ya hemos dicho, en los primates existen tres tipos diferentes de conos, cada uno de ellos es sensible de forma selectiva a la luz de una longitud de onda determinada, verde, roja o azul Tanto conos como bastones establecen sinapsis con neuronas bipolares, pero hay muchos bastones que establecen contacto con la misma célula bipolar, reforzando así la señal eléctrica que se genera en ella, pero esta mayor convergencia es una limitación para ganar en resolución espacial porque en la neurona bipolar los efectos de cada bastón se promedian. Por su parte Los conos presentan mejor resolución espacial porque muestran menos convergencia (sólo unos pocos establecen sinapsis con cada célula bipolar) y ello facilita que la imagen pueda transmitirse de forma más fidedigna manteniendo las diferencias espaciales. Además en la fóvea, donde se concentran, la imagen sufre menor distorsión y no hay convergencia de ningún tipo: cada célula bipolar recibe información de un único cono Otra diferencia entre ambos fotoreceptores es que los conos presentan mayor resolución temporal que los bastones. Los bastones disparan lentamente sumándose el efecto de los fotones que absorben cada 100milisegundos lo que les permite detectar bajas iluminaciones, pero no determinadas frecuencias de luz. En cambio la respuesta de los conos es mucho más rápida, lo que les permite detectar mejor los cambios de la iluminación en el tiempo También se diferencian en las conexiones que establecen con otras células de la retina. Por lo tanto, En el ojo de los vertebrados hay dos sistemas funcionales diferentes captadores de luz: el sistema de los conos y el sistema de los bastones. O dicho en términos más técnicos, disponemos de visión fotópica (conos) y de visión escotópica (bastones). Los mecanismos de la transducción son muy similares en ambos fotorreceptores y se llevan a cabo de manera similar a otras formas de transducción basadas en procesos de activación e inactivación de proteínas G y de segundos mensajeros. En la transducción visual el potencial de receptor constituye una excepción a la regla general ya que no se trata de una despolarización sino de una hiperpolarización. 1.-En ausencia de luz, los iones Na+ están entrando masivamente a través de los segmentos externos de los fotorreceptores lo que les mantiene despolarizados, dando como resultado la liberación del neurotransmisor glutamato. 2.-En presencia de luz, los canales de Na+ se cierran y el potencial de membrana se hiperpolariza al no entrar Na+, por lo que deja de liberarse glutamato. Los canales de Na+ se cierran porque cuando los fotones de luz son absorbidos por los fotopigmentos, se produce la descomposición de éstos en diversos compuestos a través de una cadena de reacciones moleculares. Uno de estos compuestos activa una proteína G que a su vez activa la degradación del segundo mensajero GMPC. Este segundo mensajero mantenía abiertos los canales de Na+, lo que mantenía despolarizado al fotorreceptor y al ser degradado el mensajero los canales de Na+ no pueden mantenerse abiertos y los fotorreceptores sufren una hiperpolarización Es decir, que los fotoreceptores están normalmente despolarizados, al hiperpolarizarse reducen proporcionalmente la cantidad de neurotransmisor liberado. Esto produce cambios en los potenciales de membrana de las neuronas bipolares con las que estos receptores sinaptan. Finalmente las neuronas ganglionares responderán modificando la frecuencia de potenciales de acción, que serán conducidos hacia diversas regiones cerebrales para el procesamiento superior de la información visual. 3.-LA TRANSMISIÓN DE LA INFORMACIÓN SENSORIAL AL SISTEMA NERVIOSO CENTRAL Una de las características fundamentales del procesamiento de la información sensorial en el SNC es su organización jerárquica que supone la intervención de diversos núcleos y regiones encefálicos que funcionan de forma integrada. Cada región está especializada en determinadas funciones, de modo que las señales sensoriales fluyen desde niveles inferiores a niveles superiores, estableciéndose una jerarquía en el procesamiento sensorial. Además de esta organización jerárquica existe también una organización espacial. La información procedente de los receptores sensoriales llega principalmente a la médula espinal y a regiones situadas en el tronco del encéfalo y en el diencéfalo para alcanzar finalmente la corteza cerebral (excepto la información olfativa que llega en primer lugar a los hemisferios cerebrales). Las áreas corticales se sitúan en el nivel superior (jerárquico y espacial) y son las responsables del procesamiento más complejo de la información sensorial, así como de su integración e interpretación. La información relevante sobre los estímulos sensoriales es recogida por receptores especializados situados en los órganos de los sentidos. Esta información es recibida por el SNC a través de vías aferentes que están constituidas por los axones de neuronas sensoriales que generalmente se encuentran a grupadas en los ganglios del sistema somático, que como sabemos forman parte del S.N.P, y pueden ser ganglios espinales o ganglios craneales. Las neuronas que constituyen estos ganglios son las neuronas sensoriales de primer orden o primarias. Los axones de estas neuronas sensoriales primarias (raíces dorsales de la médula y fibras aferentes de los nervios craneales) establecen sinapsis con neuronas situadas en diferentes divisiones del SNC, que constituyen las neuronas sensoriales de segundo orden. Finalmente los axones de estas neuronas secundarias alcanzan el tálamo, donde establecen sinapsis con neuronas situadas en los diversos núcleos (neuronas sensoriales de tercer orden) especializados como sabemos en el procesamiento de señales procedentes de las diferentes modalidades sensoriales. Desde estos núcleos especializados del tálamo se envían las señales de cada modalidad hacia áreas específicas de la corteza cerebral. En algunos casos, como la recepción de la información olfativa, visual o algunos tractos nocioceptivos, no se producen estos relevos sucesivos. Por ejemplo, la información visual, a través del quiasma óptico, llega directamente al tálamo y la olfatoria al bulbo olfatorio cortical. La organización jerárquica de los circuitos sensoriales implica un procesamiento en serie, que no consiste solo en la simple transmisión de información ya que en cada relevo tiene lugar una integración de las señales sensoriales que se va haciendo cada vez más compleja a medida que se alcanzan niveles superiores de procesamiento. Por otro lado, nuestro S.N. desarrolla también un procesamiento en paralelo de la información sensorial: las señales se transmiten a través de diferentes vías para ser analizadas en circuitos paralelos que desempeñan distintas funciones. Veamos algún ejemplo: el sistema somatosensorial está constituido por 2 circuitos diferentes: 1.- El sistema lemniscal: es el responsable de la transmisión de la información táctil más compleja y precisa, y de las señales propioceptivas. 2.- el sistema anterolateral: que transmite fundamentalmente las señales nociceptivas y la información sobre la temperatura. Aunque desempeñan funciones diferentes, en ocasiones hay un cierto grado de solapamiento y ambos sistemas comparten información, por ejemplo, el sistema lemniscal participa en la localización del estímulo doloroso. También en el sistema visual parecen estar implicados circuitos en paralelo en lo que se refiere a la forma de un objeto por un lado y su localización espacial por otra. Lo interesante de estas vías paralelas es que en ocasiones pueden ofrecer cierta posibilidad procesamiento alternativo en caso de lesión Otra de las características de la organización de estas vía sensoriales es la decusación o cruce contralateral que tiene lugar a diferentes niveles según el tipo de señal sensorial. Por ejemplo, en referencia a los sistemas somatosensoriales descritos más arriba: las fibras aferentes primarias del sistema lemniscal forman las columnas dorsales, axones mielinizados de gran tamaño que se disponen ordenadamente para representar con precisión el mapa de la superficie corporal y que ascienden hacia el encéfalo estableciendo sinapsis con neuronas situadas en los núcleos de las columnas dorsales del bulbo raquídeo (neuronas secundarias) de ahí que al sistema lemniscal también se le denomine sistema de las columnas dorsales. Como puede verse en la figura de más abajo, los axones de estas neuronas cruzan al lado contrario del cuerpo a nivel bulbar y forman un tracto ascendente denominado lemnisco medial que alcanza en su mayor parte el tálamo y desde donde envían proyecciones a la corteza somatosensorial. Por su parte las fibras aferentes del sistema anterolateral están constituidas por axones más finos que los del sistema lemniscal, transportan la información más lentamente con menor precisión en cuanto a la localización estimular. Estas fibras establecen sinapsis en el asta dorsal de la médula espinal con las neuronas secundarias cuyos axones cruzan al lado opuesto y ascienden a lo largo de la médula espinal alcanzando el tálamo y diversas regiones del tronco del encéfalo. Desde el tálamo la información es enviada, al igual que ocurría con el sistema lemniscal, hasta la corteza somatosensorial (también aquí la corteza cerebral de cada hemisferio recibe la información procedente del lado contralateral del cuerpo) Vemos claramente aquí el patrón general de organización jerárquica en el que intervienen los 3 tipos de neuronas (primarias, secundarias, terciarias). Otras vías sensoriales, como las auditivas o las vestibulares, decusan a nivel troncoencefálico. También en el sistema visual encontramos una decusación de las fibras que transmiten la información sobre los estímulos luminosos. Los axones de las células ganglionares de la retina, que constituyen el nervio óptico, se dividen en 2 grupos en el quiasma óptico: los que provienen de la mitad de la retina más cercana a la nariz (hemirretina nasal) cruzan hacia el lado contralateral, mientras que la otra mitad de la retina (hemirretina temporal) proyecta sus axones ipsilateralmente. En consecuencia la información proveniente de la parte derecha del campo visual se proyecta en la parte izquierda de cada retina, mientras que la información proveniente de la parte izquierda del campo visual se recibe en la mitad derecha de cada retina. A partir del quiasma óptico los axones constituyen el tracto óptico que transmite la información visual en su mayor parte al núcleo geniculado lateral del tálamo, del que parten distintas vías que constituyen la radiación óptica que alcanzan la corteza visual primaria. Los sistemas sensoriales presentan una disposición ordenada de sus aferencias que se mantiene en los diferentes niveles de relevo de la información sensorial, desde los inferiores a los superiores. 1. Organización Somatotópica: Algunas estructuras del SNC como el sistema somatosensoral, tienen una organización topográfica de sus partes (organización somatotópica); esto significa que porciones determinadas de estas estructuras se asocian a determinadas áreas topográficas del cuerpo. La organización somatotópica da lugar a un mapa de la superficie corporal que no guarda una relación directa con el tamaño de cada zona, sino que depende de la importancia funcional de esa región. Esta organización topográfica es posible gracias a los dermatomas: áreas específicas de la superficie cutánea inervadas por las fibras de cada raíz dorsal. La organización segmentaria de las raíces dorsales de la médula espinal se mantiene en los diferentes sistemas ascendentes: las fibras se organizan en una disposición ordenada que mantiene las relaciones de la superficie corporal según su origen somático. Por su parte en el sistema visual existe un mapa retinotópico que permite localizar la situación de los objetos en el campo visual. Cada punto del campo visual se proyecta en un punto determinado de la retina de forma invertida. La información procedente de cada punto del campo visual es codificada y transmitida por los axones de las células ganglionares de la retina, que se disponen de forma ordenada para representar el campo visual completo. Esta disposición ordenada de los axones se mantiene también en los niveles superiores del procesamiento visual a nivel talámico y cortical. Al igual que sucedía con el sistema somatosensorial, la representación es mayor cuanto mayor es la importancia de la región, (como es el caso de la fóvea) En el sistema auditivo también se da una disposición ordenada de los axones aferentes primarios, que nos permite distinguir las distintas frecuencias de los sonidos, como explicamos al hablar de la transducción auditiva. 3.1 UN EJEMPLO DEL PROCESAMIENTO NIVELES: EL PROCESAMIENTO VISUAL A DISTINTOS 3.1.1. PROCESAMIENTO VISUAL PRIMARIO Cuando llega la luz a los receptores visuales estos se hiperpolarizan y liberan menos glutamato y estos cambios afectan al potencial de membrana de las células bipolares y a través de estas a las ganglionares. Al conjunto de fotoreceptores que envían información de un área concreta a estas células se le llama campo receptivo .Pues bien, sucede algo muy interesante: Los campos receptivos del sistema visual son de forma más o menos circular y antagónicos. Esto último se debe a que en las primeras etapas del procesamiento de la información visual, los campos tienen una organización centro-periferia concéntrica. Los estímulos que caen sobre la periferia del campo producen una respuesta contraria a la respuesta del centro al ser estimulado. Las primeras neuronas que poseen esta conformación son las células bipolares. Según su respuesta funcional existen dos tipos: las células bipolares de centro "on" o "depolarizantes" sé despolarizan como respuesta un estímulo luminoso en el centro del campo. Por otra parte, las células bipolares de centro "off" o "hiperpolarizantes" responden contrariamente ante una estimulación de su centro. Además, se dividen en células bipolares para bastones y bipolares para conos Los terminales de los fotorreceptores pueden establecer contactos sinápticos simultáneamente con células bipolares de Centro ON y de Centro OFF de modo que existen dos modos de análisis de la información visual que se mantendrán en niveles superiores de procesamiento Los cambios en el potencial de membrana de las células bipolares producen una mayor o menor liberación de glutamato en las sinapsis que éstas establecen con las correspondientes células ganglionares. Las células ganglionares son análogas a las bipolares: también existen dos tipos: Las células ganglionares de Centro ON señalan los aumentos de la iluminación que incide en el centro de su campo receptivo, mientras que las de Centro OFF codifican los decrementos en la iluminación (al responder despolarizándose en la periferia). Por ello se denominan células sensibles al contraste Como vemos en la imagen las células ganglionares responden de forma débil cuando la iluminación es difusa y afecta tanto al centro como a la periferia y de forma marcada si se ilumina el centro (centro on) o la periferia (centro off) por eso parece que su función sería informar de los contrastes de iluminación e informarnos de los contornos de los objetos (pues los “bordes” implican diferencias de contraste de iluminación). Es decir que lo significativo y lo que recogen estas células es el contraste entre la iluminación de la figura y el fondo. Hemos hablado de los contrastes de iluminación pero también existen células ganglionares sensibles a la dirección del movimiento (es decir que responder a un punto de luz que se mueve en una dirección concreta, independientemente de que se localice en el centro o en la periferia) respondiendo a cambios en la iluminación en función del tiempo en lugar de a cambios espaciales. Otras células ganglionares presentan también campos antagónicos, pero esta vez referidos al color, se las llama células oponentes al color 3.1.2. PROCESAMIENTO VISUAL EN EL TALAMO El tálamo es una especie de antesala a la cual llega la información sensorial procedente de la médula y el tronco del encéfalo (si bien la información visual llega directamente al tálamo y la olfatoria, pasa primero por la corteza y llega al tálamo en segundo lugar) El tálamo está organizado en diferentes grupos nucleares, entre los que se encuentran los núcleos de relevo sensorial, los que se encuentran en el grupo ventral no son simples estaciones de relevo sino que también interviene en el procesamiento de la información: cada núcleo en el de una modalidad sensorial diferente. Estos núcleos envían también de forma específica proyecciones a áreas concretas de la corteza cerebral, recibiendo también una retroalimentación por parte de estas áreas corticales. Aparte de diferir en su función los núcleos talámicos difieren en su citoarquitectura. Algunos tienen poblaciones celulares muy uniformes y otros se organizan en capas. En el caso del sistema visual,las señales llegan al núcleo geniculado lateral, cuyas células se agrupan en función de su tamaño y de las proyecciones que reciben de la retina, en capas parvocelulares (células de pequeño tamaño) y capas magnocelulares (células de gran tamaño) de manera que se forman dos sistemas diferentes, aunque a veces comparten información: el sistema parvocelular y el sistema magnocelular que proyectan la información a distintas zonas de la corteza. Parece ser que las neuronas de este núcleo son similares a las células bipolares y ganglionares y se organizan concéntricamente con células centro on y off Las neuronas magnocelulares poseen campos receptivos más grandes, responden a grandes objetos y son sensibles al movimiento, contribuyendo a la percepción de las características generales del estímulo Las neuronas parvocelulares por su parte, tienen campos receptivos más pequeños y son responsables del análisis de los detalles finos de la imagen e intervienen en la percepción de la forma y del color. Una vez que las señales visuales han sido procesadas en el tálamo, la información es enviada a la corteza visual y a otras regiones encefálicas a través de las radiaciones ópticas 3.1.3. PROCESAMIENTO VISUAL A NIVEL CORTICAL Como hemos visto la información que llega al núcleo geniculado lateral del tálamo, es enviada mediante las radiaciones ópticas a un área concreta de la corteza cerebral situada en el lóbulo occipital, la corteza visual primaria (V1), aunque existen también otras áreas de la corteza visual denominadas V2, V3, V4 y V5 que procesan también diversos aspectos de la información visual, con funciones menos conocidas, por lo que nos centraremos en el área visual primaria Celulas simples y complejas En la corteza visual, al igual que otras áreas de la neocorteza, está constituida por seis capas, cuya organización es compleja y jerarquizada, de modo que mientras algunas de sus neuronas responden a estímulos relativamente simples, otras necesitan estímulos cada vez más complejos. Se distinguen dos tipos de células: 1.- Células simples: al igual que las bipolares y ganglionares, presentan campos receptivos con una zona central y una zona periférica, pero los estímulos que son óptimos para la activación de las células simples no son puntos o anillos de luz, sino más bien tienen forma de barras alargadas. Cada célula simple responde a una orientación característica del estímulo en su campo receptivo y, al mismo tiempo, todas las orientaciones posibles del estímulo están presentes en el conjunto global de la población de las células simples. Por otra parte no todas las neuronas presentan zonas excitadoras e inhibidoras del tipo un centro rodeado por una periferia. Algunas tienen una zona excitadora que ocupa la mayor parte de la neurona, con una pequeña zona periférica en un lateral. Otras tienen campos receptivos con una zona excitadora ocupando exactamente la mitad del mismo y una zona inhibidora ocupando la otra mitad 2.- Células complejas: Existen diferentes tipos de células complejas que están organizadas jerárquicamente dependiendo de las características de los estímulos que son capaces de desencadenar su activación. Son las más abundantes de la corteza visual y a diferencia de las células simples que están agrupadas en su mayor parte en el área V1, se localizan también en la V2 y en otras áreas de la corteza visual. Este tipo de neuronas son estimuladas por el movimiento brusco de un hilo o una barra de luz alargada que recorre su campo receptivo, pero con una orientación determinada. Las células complejas tienen unos campos receptivos más grandes que los de las células simples, lo que sugiere que estos campos receptivos son el resultado de aferencias provenientes de diversas células simples, que están organizadas espacialmente de una forma característica. De manera que cada nivel superior de complejidad va integrando información de los niveles inferiores (ver fig.11.43) A esta propiedad que tienen las células complejas de responder ante movimientos del estímulo luminoso en una dirección y no hacerlo cuando el estímulo se presenta en otras direcciones se le llama selectividad de la dirección. Otra propiedad de estas células corticales es la finalización del campo receptivo, por la que la mayoría de estas células responden mejor ante un estímulo cuyo tamaño se ajusta a su campo receptivo, y su respuesta es más débil sí la barra de luz es más corta que el propio campo receptivo El movimiento tiene gran importancia en la percepción visual, se ha comprobado que si las imágenes fueran perfectamente fijas, al cabo de poco tiempo no se percibirían. Que sea un factor tan importante a la hora de percibir nuestro medio ambiente explica que la mayoría de las células complejas del área V2 respondan selectivamente al movimiento y que las células simples (que no detectan movimiento) disminuyan en los centros superiores de análisis de la información visual Columnas y módulos corticales En la corteza visual primaria de los mamíferos se ha comprobado que la mayoría de las neuronas, simples o complejas, reciben información procedente de los 2 ojos, son binoculares. Sin embargo, la mayoría de ellas reciben más aferencias desde un ojo que desde el otro, lo que se conoce como dominancia ocular. Las células con la misma dominancia ocular, están organizadas en columnas de 1mm de ancho llamadas columnas de dominancia ocular. También existen columnas de orientación ya que las neuronas (simples o complejas) que responden a la misma orientación del estímulo se sitúan en la misma columna. Las células de las columnas corticales responden ordenadamente a cambios sucesivos de 10 grados cubriendo así una rotación de 180 grados. Al conjunto de las columnas que cubren todos los posibles ángulos de orientación se les llama hipercolumnas Las agrupaciones neuronales sensibles al color se denominan estacas o manchas (blobs) y se disponen exactamente alineadas en las columnas de dominancia ocular atravesando las capas de la corteza primaria. Muchas de las células de las estacas son oponentes al color (por ejemplo centro Rojo+, periferia Verde-). Pero también hay células corticales del tipo: centro R+V- periferia R-V+, a este tipo de células se les llama doblemente oponentes al color Las columnas de dominancia ocular y las de orientación se disponen formando ángulos entre sí, de forma que pueden hacerse bloques de corteza que contienen todos los elementos neurales necesarios para el procesamiento de la información visual procedente de la retina. A estos bloques de corteza se les denomina módulos corticales. Los módulos serian las entidades mínimas capaces de procesar todos los aspectos de los estímulos que llegan a una zona determinada de la retina: forma, contornos, orientación, color…Miles de estos módulos procesarían en conjunto y en paralelo la imagen que se recoge en la retina Nos queda hablar de las dos tipos más de células las responsables de la percepción de la profundidad (que requieren un alineamiento binocular exacto del estímulo para responder) y las células moduladoras de la disparidad, que detectan las pequeñas diferencias en la percepción posición de la imagen derivadas de que la separación de los ojos. Como hemos ido viendo en este apasionante viaje a través del sentido de la vista, en todos los niveles se mantiene una segregación de funciones: el color, el movimiento, la forma o la profundidad se analizan de forma separada. Por otra parte no hay ningún área privilegiada del cerebro que se encargue de “unificar” esta imagen, son todas las áreas implicadas las que generan la imagen integrada ¿Cómo realizan semejante azaña ? eso es algo que, aun, continua siendo un misterio. 3.2. OTRO EJEMPLO DE PROCESAMIENTO CORTICAL: EL PROCESAMIENTO SOMATOSENSORIAL La corteza somatosensorial está estructurada por zonas, todas dentro del lóbulo parietal. •S1 o corteza somatosesnsorial primaria, compuesta por tres áreas de Brodmann (1, 2, 3a, 3b) •S2 corteza somatosensorial secundaria (situada lateral y debajo del área primaria) •Corteza parietal posterior, compuesta por otras dos áreas de Brodmann (5 y 7). En ella se producen procesos complejos que relacionan distintas sensaciones para la identificación de objetos (por ej. diferenciar una llave de una cuchara) El Mapa somatosensorial: El córtex tiene el equivalente a un mapa de todo el cuerpo, reflejándose en este mapa la procedencia original de cada sensación.La correspondencia entre sensaciones superficiales y corteza cerebral se denomina somatotopia cortical. La superficie dedicada a cada órgano depende del número de terminaciones nerviosas en el mismo, siendo muy distinta para unas partes del cuerpo que para otras y especialmente grande para la boca, lengua y dedos de las manos. Organización en columnas de la corteza El cortéx somatosensorial, al igual que el córtex visual y auditivo, está organizado en columnas, cada una de ellas especializada en las diferentes sensaciones procedentes de una misma zona del cuerpo. Sin embargo no percibimos las características de los objetos en “listas” sino que percibimos objetos completos. Esto es porque hay zonas donde convergen aferencias. Por ejemplo en las capas 3ª y 3b se procesa la información que llega de los dedos centrales de la mano por separado. En las capas siguientes del córtex (1 y 2) la información se integra de manera que se perciban objetos, más que características separadas. El nivel de procesamiento más elaborado se produce en la corteza parietal posterior, que integra también la información de otros sistemas, como el visual, en la que las sensaciones de textura, tamaño, forma se entienden como características de un mismo objeto. Las lesiones en esta área provocan síndromes muy complejos como la astereognosia o la hemineglicencia. Existen muchos paralelismos con la percepción visual. Las sensaciones se transmiten pasando por el tálamo hacía el córtex. En éste se analizan inicialmente las características independientes que son proyectadas en módulos corticales y que posteriormente se van a ir integrando hasta tener percepción de los objetos del entorno. 3.3.- VIAS DE MODULACIÓN SENSORIAL: LA MODULACIÓN DE LA PERCEPCIÓN DEL DOLOR Las investigaciones han puesto de manifiesto la existencia de mecanismos neurales que reducen la sensibilidad al dolor, de modo que la información nociceptiva puede ser modulada tanto por las aferencias sensoriales nociceptivas que recibe la médula espinal como por la activación de vías descendentes procedentes de diferentes estructuras del encéfalo que alcanzan la médula espinal y el tronco del encéfalo. Las fibras nociceptivas llegan al asta dorsal de la médula espinal donde establecen sinapsis con diferentes tipos de neuronas, parte de cuyos axones constituyen las vías ascendentes que transmiten la información nociceptiva a centros superiores del SNC. Especialmente importante en este sentido es el tracto espinotalamico si bien otros sistemas somatosensoriales intervienen también (por ejemplo el sistema lemniscal interviene en la localización del estímulo doloroso). No obstante los núcleos talámicos que reciben información nociceptiva la proyectan de manera difusa, es decir que no parece existir un mapa nociceptivo similar a los mapas somatotopicos de las aferencias táctiles. Todos hemos oído hablar de situaciones: soldados en la batalla, competiciones deportivas, momentos de alto estrés en que se reduce la percepción al dolor y esto es un hecho comprobado, aunque sorprendente. Pero también en situaciones cotidianas se puede producir analgesia. Pensemos por ejemplo en lo común que es frotar o masajear la parte dolorida reduciendo así el dolor. Esta reducción se produce porque En la médula espinal, la actividad de las neuronas que reciben información nociceptiva es modulada por otras aferencias sensoriales que convergen sobre ellas así la estimulación de las fibras procedentes de los mecanorreceptores hace disminuir la respuesta de las neuronas del asta doral a la información que llega a través de las fibras nociceptivas (de forma semejante a lo que ocurría con la inhibición lateral) En este sentido se ha se ha propuesto la existencia de circuitos neurales específicos que modulan la transmisión de la información nociceptiva y que se conoce como Teoría del control de entrada del dolor como vemos en la imagen: Especial importancia tienen las llamadas vías descendentes de la información nocioceptiva. Se ha comprobado con animales que la estimulación de la sustancia gris periacuedutal produce una analgesia lo suficientemente profunda como para servir de anestésico en una operación. Especialmente importantes son en estas vías descendentes, la sustancia gris periacueductal (GPA), el núcleo magno del rafe (NMR) y el núcleo reticular paragigantocelular (NRPG) Los axones que parten de estos núcleos se dirigen hacia la médula espinal formando parte del fascículo dorsolateral y alcanzando el asta dorsal a diferentes niveles medulares, donde modulan la transmisión de la información nociceptiva. ¿Cómo inhiben estas vías el dolor? Pues a través de la intermediación de los péptidos opioides endógenos que se unen a receptores específicos ampliamente distribuidos por todo el SNC, los receptores opioides Las neuronas de la GPA reciben a su vez proyecciones descendentes que proceden de distintas zonas encefálicas como el hipotálamo, diferentes núcleos del tálamo, la amígdala y la corteza frontal lo que podría estar implicado en el hecho de que en determinadas situaciones de estrés se suprime el dolor Por otra parte la selección natural habría primado este sistema ya que cuando un animal tiene una herida el dolor le lleva a generar respuestas de descanso destinadas a recuperarse pero este tipo de respuestas podría interferir con conductas más adaptativas como buscar comida o cuidar a las crías. A este tipo de activación de los sistemas analgésicos endógenos se le ha llamado analgesia inducida por el estrés. 4.- ALGUNOS EJEMPLOS SOBRE LAS FUNCIONES BIOLOGICAS DE LOS SENTIDOS En todas las culturas humanas una buena comida se asocia al placer. Y no nos pasa solo a nosotros, las porciones de sabroso alimento son utilizadas por los investigadores como refuerzo para conseguir motivar a las ratas a realizar tareas que requieren aprendizaje. La llamada fase cefálica es la responsable del aspecto placentero de la digestión y consiste en un conjunto de reflejos (secreción de saliva, secreciones gástricas…) que se ponen en marcha al ser estimulados los receptores sensoriales del olfato y el gusto, junto con otras áreas de la boca y el encéfalo. Son hechos demostrados que a mejor calidad de la comida más activación de los reflejos cefálicos y que si se eliminan farmacológicamente o mediante lesión estos reflejos la ingesta deja de proporcionar placer. El sentido del gusto también ha evolucionado para ayudar a los animales a escoger adecuadamente la dieta como demuestran: - Las hambres especificas: Como el hambre de sal en el ganado ya que el sodio es un elemento esencial o el general gusto por los dulces y el chocolate ya que la glucosa también es esencial -La aversión gustativa: consiste en que un animal aprende a evitar una comida determinada si en alguna ocasión su ingestión vino seguida al cabo de minutos o incluso horas, de sensaciones desagradables (mareos, vómitos). La aversión gustativa es un tipo de asociación entre el sabor de la comida ingerida (EC), sea el que sea, con el malestar posterior (EI) que se da en un lapso de tiempo mucho mayor de lo que las teorías conductistas postulaban por lo que se conoce como aprendizaje biológicamente preparado. Por su parte el olfato juega un papel muy importante en el despliegue de la conducta sexual de casi todos los mamíferos, incluido el hombre, así como en otro tipo de conductas esenciales: marcación del territorio, conductas agresivas, conducta maternal No es extraño pues que (pese a que la especie humana no es muy ducha en el sentido del olfato) que algunos aromas como el almizcle o la algalia, nos resulten cautivadores, ya que sus principios esenciales proceden de glándulas secretoras de algunos animales, que utilizan estos componentes como reclamo sexual. Las sustancias odoríferas o feromonas son mensajeros químicos que interactúan con receptores específicos desencadenando determinados procesos fisiológicos o promoviendo conductas específicas. Las feromonas afectan directamente a la secreción de hormonas hipotalámicas e hipofisarias y provocan efectos muy característicos: -Efecto Bruce: ha sido estudiado fundamentalmente en roedores. En esta situación, un macho que se encuentra con una hembra embarazada por otro macho, puede emitir feromonas de tal intensidad que la hembra pierde el embarazo y queda en poco tiempo con capacidad de aparearse nuevamente -Efecto Vandenberg: también ha sido comprobado que el inicio de la pubertad también puede acelerarse por el olor de un macho En cuanto a nuestra especie, todas las mujeres sabemos que cuando convivimos con otra mujer, nuestros periodos tienden a sincronizarse. Estos son solo algunos ejemplos de los cientos que hay, para hacernos una idea de lo útiles que son nuestros sentidos para adaptarnos al ambiente.