metabolismo iii: fotosíntesis
Anuncio

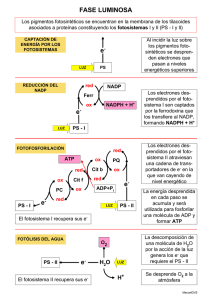
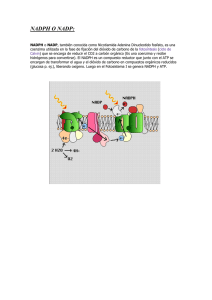
METABOLISMO III: FOTOSÍNTESIS METABOLISMO III: FOTOSÍNTESIS FOTOSÍNTESIS. En un sentido estricto, la fotosíntesis es un proceso consistente en la conversión de energía lumínica en energía química. La realizan ciertos organismos, tales como, plantas, algas, bacterias fotosintéticas y algunos protozoos. Estos organismos captan la radiación lumínica y la transforman en compuestos de alto poder reductor (NADPH) y compuestos con enlaces ricos en energía (ATP) . Estos productos que se forman durante la fase lumínica de la fotosíntesis, se utilizan durante la fase oscura para la síntesis de materia orgánica a partir de materia inorgánica. 1. FASE LUMÍNICA DE LA FOTOSÍNTESIS. 1.1. Absorción de energía lumínica: Fotosistemas. Se realiza en la membrana de los tilacoides del interior de los cloroplastos, donde se localizan pigmentos fotosensibles (clorofilas a y b, carotenos, xantofilas etc), formando antenas solares denominadas complejos colectores de luz ( CCL 1 / 17 METABOLISMO III: FOTOSÍNTESIS ), que son verdaderas trampas para los fotones. Cuando un fotón choca con un electrón de un pigmento fotosintético, la energía del fotón (E=hv) es absorbida por el electrón el cual salta a un nivel más alejado del núcleo, pudiendo dejar ionizado al átomo. El pigmento queda de esta forma con un defecto de electrones es decir oxidado. Los pigmentos fotosintéticos están asociados a proteínas formando los fotosistemas, constituidos por la antena y el centro de reacción . Los pigmentos de la antena encauzan la energía fotónica hasta el centro de reacción , el cual esta constituido por una clorofila diana a la que va a parar la energía de excitación, transfiriendo así electrones al aceptor primario de electrones , que a su vez los transfiere fuera del fotosistema. La clorofila diana queda así oxidada, recuperando los electrones de una molécula denominada dador primario de electrones. Según la longitud de onda a la que se excitan los pigmentos fotosintéticos podemos diferenciar dos fotosistemas diferentes: Fotosistema I: capta luz de £ 700 nm. La antena esta constituida por clorofilas a, b y carotenos. La clorofila diana es un dímero de clorofila a I que absorbe a 700 nm, antes conocida como P700. El aceptor primario es poco conocido y se llama aceptor X y el dador es la plastocianina . 2 / 17 METABOLISMO III: FOTOSÍNTESIS Fotosistema II: capta luz de £ 680 nm. La antena esta constituida por clorofilas a, b y xantofilas. La clorofila diana es un dímero de clorofila a II que absorbe a 680 nm, antes conocida como P680. El aceptor primario es el aceptor Q (probablemente una plastoquinona) y el dador primario se denomina dador Z y es también poco conocido. 1.2. Transporte de electrones: cadena fotosintética. La energía lumínica se va transformar en moléculas de alto poder reductor NADPH. El transporte de electrones desde el agua hasta el NADPH se realiza a través de una cadena de transportadores situada en la membrana de los tilacoides, en la que interviene de forma activa los dos fotosistemas. Esta cadena fotosintética se puede dividir en tres segmentos: (Figura) I Transporte de e - desde el agua hasta el Fotosistema II. La fotolisis del agua es un proceso poco conocido que libera 2 electrones que son transportados hasta 2 moléculas de Chl a II del fotosistema II (que se encuentran oxidadas por una reacción fotoquímica anterior). Los electrones son transportados a través de un intermediario Z que es considerado el dador primario de electrones del fotosistema II. La fotolisis del agua ocurre en el espacio intratilacoidal, donde quedan liberados el oxígeno y los protones. H 2 O --luz---® ½ O 2 + 2 H + + 2 e - 3 / 17 METABOLISMO III: FOTOSÍNTESIS II Transporte de e - desde el fotosistema II hasta el fotosistema I. La reacción de oxidación fotoquímica consiste en que la energía de 2 fotones de £ 680 nm hacen pasar 2 electrón desde 2 moléculas Chl a II hasta el aceptor primario de electrones Q . Las moléculas de clorofila quedan oxidadas, pasando a su forma reducida gracias a los electrones procedentes del agua. Gracias a esta activación fotoquímica, los electrones aumentan su nivel de energía pasando de un potencial redox + 1,0 V a un potencial de 0 V. Los electrones son transportados hasta el fotosistema I a través de una cadena de transportadores constituida en orden creciente de sus potenciales redox, plastoquinona ( PQ ), citocromo f y plastocianina ( PC ) siendo este último, considerado el dador primario del fotosistema I, cede los electrones a 2 moléculas de Chl a I que se encuentran oxidadas por una segunda reacción fotoquímica anterior. El transporte de electrones a lo largo de esta cadena supone una perdida de energía que es empleada en la síntesis de ATP . III Transporte de e - desde el fotosistema I al NADPH De nuevo la energía de 2 fotones, esta vez de £ 700 nm, va a hacer que dos electrones 4 / 17 METABOLISMO III: FOTOSÍNTESIS procedentes de dos moléculas de Chl a I se activen reduciendo una molécula del aceptor de electrones X. Las moléculas de clorofila oxidadas pasan a su forma reducida al captar los electrones procedentes del fotosistema II. Los electrones son cedidos desde el aceptor X a la ferredoxina FD, luego a la reductasa , y por último al NADP + que queda reducido a NADPH más H + . Globalmente el proceso se resume: H 2 0 + NADP + + 4 fotones ----------® ½ O 2 + NADPH + H + Son necesarias la fotolisis de 2 moléculas de agua para que se forme una molécula de oxígeno. Los cuatro fotones actúan dos sobre el fotosistema II y dos sobre el fotosistema I, de manera 5 / 17 METABOLISMO III: FOTOSÍNTESIS que los 2 electrones que provienen del agua son impulsados energéticamente "cuesta arriba" en dos ocasiones. Así, a partir de una molécula como el agua con potencial redox positivo (escasamente reductora), se obtiene una molécula de NADPH de potencial redox negativo (reductor fuerte), útil es las reacciones de reducción de la fase oscura de la fotosínteis. Comparativamente el transporte de electrones por la cadena fotosintética es contrario al transporte en la cadena respiratoria donde los electrones eran transportados desde el NADPH hasta el agua. Transporte cíclico de electrones A nivel de los tilacoides existe un segundo proceso de transporte de electrones que requiere una sola reacción fotoquímica, la del fotosistema I. El aceptor primario X transfiere los electrones provenientes de la Chl a I hasta la ferredoxina la cual en lugar de transferirlos al NADP, lo hace al citocromo b 6 , el cual lo transfiere de nuevo a la Chla I a través de la plastoquinona, citocromo f y plastocianina. En el transporte cíclico no hay producción de NADPH ni se libera oxígeno pues no hay fotolisis del agua. Este proceso cíclico tiene como única misión la síntesis de ATP . El que se de este transporte cíclico o no depende de la cantidad de NADPH, si hay poco los electrones son transportados de manera no cíclica, si hay mucho siguen este transporte cíclico. (Para algunos autores este transporte cíclico no es más que un artefacto experimental). Los procesos fotosintéticos donde el agua actúa como molécula reductora y en los que se libera oxígeno, se denominan fotosínteis oxigénica, para diferenciarlos de otros procesos fotosintéticos donde el dador de electrones no es el agua y por tanto no liberan oxígeno, que se denominan fotosíntesis anoxigénica. 6 / 17 METABOLISMO III: FOTOSÍNTESIS La fotosínteis anoxigénica la realizan algunos grupos de bacterias como las sulfobacterias purpúreas que utilizan H 2 S como dador de electrones; las sulfobacterias verdes que también utilizan H 2 S pero no acumulan azufre y las bacterias verdes no sulfúreas , que utilizan moléculas orgánicas como agentes reductores. De cualquier forma, en estas bacterias se producen moléculas reductoras de NADPH, que al igual que las producidas en la fotosíntesis oxigénica se emplean en la síntesis de ATP y reducción de materia inorgánica. 1.3. Traslocación de protones y fotofosforilación El ordenamiento de los trasportadores de electrones en la membrana tilacoidal tiene como consecuencia el que el transporte de electrones se acompañe de una liberación de protones en el espacio intratilacoidal, procedentes del estroma. Al igual que en la fosforilación oxidativa, la energía liberada en el transporte fotosintético de electrones desde el fotosistema II al I se emplea para generar un gradiente electroquímico capaz de generar una fuerza protonmotriz, que sirve para que los sistemas ATPasa localizados en la membrana tilacoidal sinteticen ATP. Parece que son necesarios la traslocación de tres protones para la síntesis de una molécula de ATP (no hay unanimidad en este número). La fotolisis de una molécula de agua y el consiguiente transporte de los electrones hasta el NADPH, libera 4 H + en el espacio intratilacoidal, 2 provenientes del agua y dos son bombeados desde el estroma por la plastoquinona. De esta forma se formaría una molécula de ATP por cada molécula de agua que se rompe. Sin embargo, para algunos autores el transporte de electrones desde el agua hasta el fotosistema II libera suficiente energía como para la síntesis de otra molécula de ATP, de manera que hay dos posibles balances de la fotofosforilación acoplada al transporte no cíclico: 7 / 17 METABOLISMO III: FOTOSÍNTESIS a) 2 H 2 O + 2 NADP + + 2 ADP + 2 Pi + 8 fotones ----------® O 2 + 2 NADPH + 2 H + + 2 ATP b) 2 H 2 O + 2 NADP + + 4 ADP + 4 Pi + 8 fotones ----------® O 2 + 2 NADPH + 2 H + + 4 ATP En el caso a, la relación NADPH/ATP es 1/1, sin embargo, durante la fase oscura de la fotosíntesis hacen falta 3 ATP por cada 2 NADPH. Para los científicos que proponen este balance, esta mayor demanda de ATP se compensaría con el producido por el transporte cíclico. En el caso b, la relación NADPH/ATP es 1/2. Sería suficiente para la fase oscura y el transporte cíclico solo sería un modo de control del NADPH o incluso un artefacto experimental. Comparación entre fosforilación oxidativa y fotofosforilación Ambos mecanismos, siempre según la hipótesis de Mitchel, son muy similares, teniendo en común las siguientes características: - Las reacciones ocurren en una membrana poco permeable a los protones: la membrana del tilacoide y la membrana interna de la mitocondria que delimitan un compartimento cerrado: el espacio intratilacoidal y la matriz. 8 / 17 METABOLISMO III: FOTOSÍNTESIS - En ambos casos los transportadores de electrones se alternan con transportadores de protones que bombean protones desde la matriz al espacio intermembrana y del estroma al espacio intratilacoidal. - La traslocación de protones genera en ambos casos un gradiente electroquímico que tiene dos componentes: un gradiente de pH y un gradiente de cargas eléctricas (una diferencia de potencial). De esta forma los espacios intratilacoidal e intermembrana se hacen más ácidos y más positivos que el estroma y la matriz, respectivamente. - El gradiente electroquímico genera una fuerza llamada protonmotriz que impulsa los protones hacia el estroma o la matriz. - Los protones solo pueden atravesar la membrana a través de las ATPasas, que en su base hidrófoba se comportan como transportadores de protones, los cuales impulsados por la fuerza protonmotriz pasan a través de la esfera de la ATPasa permitiendo la síntesis del ATP. 2. FASE OSCURA DE LA FOTOSÍNTESIS Durante la fase oscura el NADPH y el ATP producidos durante la fase lumínica son utilizados en el estroma para la síntesis de moléculas orgánicas por reducción de moléculas o iones inorgánicos como el anhídrido carbónico, el nitrato o el sulfato. 2.1. Reducción fotosintética del anhídrido carbónico: Ciclo de Calvin. Los trabajos de Calvin, Basshan y Benson realizados entre 1946 y 1953, demostraron que el C O 9 / 17 METABOLISMO III: FOTOSÍNTESIS 2 es incorporado por fijación, sobre un aceptor de C5: la ribulosa 1,5-difosfato , dando un compuesto intermediario de C6 muy inestable, el cual se escinde en dos moléculas C3 de ácido 3-fosfoglicérico ( APG ). Ambas reacciones, fijación y escisión, son catalizadas por la enzima ribulosa-1,5-difosfato carboxilasa-oxigenasa ( Rubisco ). En el estroma las moléculas de APG son fosforiladas y reducidas mediante dos reacciones consecutivas de fosforilación y oxido-reducción, que emplean la misma cantidad de ATP y NA DPH , produciéndose osas de 3 carbonos -el gliceraldehído 3-fosfato y su isómero hidroxicetona fosfato -, que pueden seguir dos caminos: - una parte son exportadas al citoplasma, donde sirven para la síntesis de hexosas (glucosa) o almidón. - otra parte es utilizada en el estroma para regenerar el aceptor C5, mediante un conjunto de complejas reacciones en las que intervienen triosas, tetrosas, pentosas, hexosas y heptos as que intercambian fragmentos de dos o tres átomos de carbono mediante reacciones de condensación y transcetolización, cuya última reacción es una fosforilación que requiere ATP. Todo este conjunto de reacciones constituyen el llamado ciclo de Calvin que podemos ver resumidamente en la figura. Tal y como podemos ver la fijación de 3 CO 2 requieren 9 ATP y 6 10 / 17 METABOLISMO III: FOTOSÍNTESIS NADPH para dar una molécula de gliceraldehído 3-fosfato . Por tanto para la síntesis de una molécula de glucosa harán falta 18 ATP y 12 NADPH . La ecuación global de síntesis fotosintética de una molécula de glucosa será: 6 CO 2 + 12 H 2 O -----LUZ----® C 6 H 12 O 6 + 6 H 2 O + 6 O 2 * Fotorrespiración La enzima Rubisco es bifuncional, actuando como carboxilasa cuando [CO 2 ] ñ [O 2 ] o como o xidasa cuando [CO 2 ] á [O 2 ]. Como carboxilasa fija 1 molécula de CO 2 en la ribulasa 1,5-difosfato para dar 2 moléculas de APG , tal y como hemos visto anteriormente. 11 / 17 METABOLISMO III: FOTOSÍNTESIS RuDP + CO 2 -------------® 2 APG Como oxidasa NO fija CO 2 y la ribulasa 1,5-difosfato se oxida (consume O 2 ) produciendo 1 mo lécula de APG , que sigue el ciclo de Calvin, y 1 molécula de ac. fosfoglicólico que es transportado a los peroxisomas donde a partir de 2 moléculas se obtiene 1 molécula de APG y una 1 de CO 2 . 2 RuDP + 2 O 2 -----------® 2 APG + 2 PGli 2 PGli ------------------------® 1 APG + 2 CO 2 Globalmente: 2 RuDP + 2 O2 ------------------------® 3 APG + 2 CO 2 12 / 17 METABOLISMO III: FOTOSÍNTESIS Debido a que es un proceso que consume oxígeno y libera CO 2 , pero esta asociado a la fotosíntesis, se denomina fotor respiración. Supone una perdida de eficacia en la fijación fotosintética del CO 2 durante el ciclo de Calvin, siendo un problema grave cuando las plantas están sometidas a elevadas temperaturas y sequedad (verano y climas cálidos), pues en estas condiciones la planta cierra sus estomas dificultando la entrada de CO 2 y la salida de O 2 , lo cual favorece la fotorrespiración. Sin embargo, la fotorrespiración no es un problema para algunas géneros de plantas que viven en climas cálidos y que han adaptado su metabolismo fotosintético con el fin de evitar la fotorrespiración. A este grupo de plantas, entre las que se encuentra la caña de azúcar, se llaman plantas C4 debido a que el CO 2 se fija previamente sobre una molécula de tres carbonos para dar una molécula de 4 carbonos (C4), a diferencia de la gran mayoría de las plantas que como hemos visto fija el CO 2 sobre una molécula de cinco carbonos para dar dos moléculas de 3 carbonos (C3), razón por la que se las llama plantas C3. * Ciclo de Hatch-Slack En las plantas C3 el ciclo de Calvin y la fotorrespiración ocurren simultáneamente, mientras que en las plantas C4 apenas se produce fotorrespiración. Esto es posible gracias a la estructura anatómica de las hojas de estas plantas. Las células fotosintéticas se distribuyen en dos capas concéntricas alrededor de los vasos transportadores de savia. En la corona externa, las células del mesófilo realizan la fijación del CO 2 sobre para dar moléculas de 4 carbonos, mientras que en la corona interna, las células de la vaina perivascular donde se realiza el ciclo de Calvin. Las interrelaciones entre ambas coronas se conoce como ciclo de Hatch-Slack (Fig). 13 / 17 METABOLISMO III: FOTOSÍNTESIS En las células del mesófilo, el CO 2 se fija sobre el ac. fosfoenolpirúvico (C3) para dar ac. oxalacético ( C4 ). La enzima que cataliza esta reacción es una carboxilasa con más afinidad por el CO 2 que la Rubisco. El ac. oxalacético se transforma en ac. aspártico o en ac. málico, que son transportados a las células de la vaina, donde sufren descarboxilación para dar por un lado ac. pirúvico ( C3 ) que es exportado a las células del mesófilo para cerrar el ciclo como fosfoenolpirúvico, y por otro lado CO 2 que se incorpora en el ciclo de Calvin. Por tanto, podemos considerar que este ciclo sirve como lanzadera de CO 2 hacia las células de la vaina con el fin de que la [CO 2 ] sea mucho mayor que la [O 2 ] y así la Rubisco actúe como carboxilasa y no como oxidasa evitando la fotorrespiración. 2.2. Reducción fotosintética del nitrógeno. Se realiza en tres fases; 1) Los iones nitrato (NO 3 - ) son reducidos a nitrito (NO 2 - ) por la enzima nitrato reductasa. 14 / 17 METABOLISMO III: FOTOSÍNTESIS NO 3 - + NADPH + H + --------® NO 2 - + NADP + 2) Los iones nitrito son reducidos a amoníaco por el enzima nitrito reductasa. NO 2 - + NADPH + H + --------® NH 3 + NADP + 3) el amoníaco se incorpora como grupo amino del ácido glutámico que posteriormente sirve en la síntesis de otros ami noácidos . ac. ?-cetoglutárico + NH 3 + NADPH + H + --------® ac. glutámico + NADP + + H 2 O. 2.3. Reducción fotosintética del azufre. También se realiza en tres fases: 1) Los iones sulfato (SO 4 2- ) son reducidos a sulfito (SO 3 2- ). SO 4 2- + NADPH + H + + ATP --------® SO 3 2- + NADP + + ADP + Pi 2) Los iones sulfito son reducidos a sulfuro de hidrógeno (H 2 S). 15 / 17 METABOLISMO III: FOTOSÍNTESIS SO 3 2- + NADPH + H + + ATP --------® H 2 S + NADP + + ADP + Pi 3) El sulfuro de hidrógeno se incorpora como grupo tiol (-SH) de la cisteína. Acetilserina + H 2 S --------® Acetato + Cisteína 3. FACTORES QUE INFLUYEN EN LA FOTOSÍNTEIS a) Intensidad luminosa: La actividad fotosintética aumenta con la intensidad, hasta un límite que depende de la especie vegetal. Así, hay especies de penumbra y especies fotófilas. En general las plantas C4 presentan mayor rendimiento fotosintético para una misma intensidad luminosa que las plantas C3. b) Color de la luz: Las plantas precisan luz roja, cuya longitud de onda es superior a 680 nm, y luz azul de longitud de onda inferior a 680. Cuando sólo se ilumina con luz roja solo funciona el fotosistema I, lo cual desencadena el transporte cíclico de electrones y por tanto el rendimiento fotosintético disminuye. Esto se conoce como declive del rojo y se produce al atardecer. c) Fotoperiodo: El rendimiento fotosintético esta relacionado con el tiempo de luz y el tiempo de oscuridad. En general a mayor cantidad de luz mayor rendimiento fotosintético. Sin embargo, aunque las fases luminosa y oscura ocurren simultáneamente, son conveniente periodos de oscuridad en los que la planta regenera el NADP reducido durante la fase luminosa. d) Temperatura: Dentro del intervalo de temperatura al que cada especie esta adaptada a vivir, existe una temperatura óptima a la cual las enzimas funcionan a pleno rendimiento. 16 / 17 METABOLISMO III: FOTOSÍNTESIS d) Humedad ambiental: La humedad ambiental condiciona la apertura y cierre de los estomas. Si el ambiente es seco se cierran si es húmedo se abren. Esto afecta a las plantas C3, pues el cierre de los estomas produce un incremento de la fotorrespiración. e) Concentración de O 2 : Cuando aumenta la concentración de oxígeno, se incrementa la fotorrespiración y por tanto disminuye el rendimiento fotosintético. d) Concentración de CO 2 : En este caso ocurre lo contrario. Sin embargo, hay un concentración máxima de CO 2 por encima de la cual no se incrementa el rendimiento fotosintético, debido a la saturación del enzima Rubisco. (saturación por sustrato) 17 / 17