17.- FISIOLOGÍA CELULAR, LAS MOLÉCULAS EN ACCIÓN
Anuncio

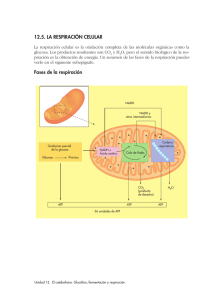
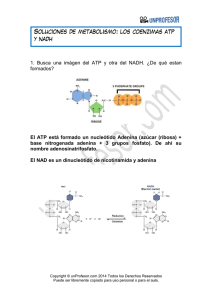
FISIOLOGÍA CELULAR, LAS MOLÉCULAS EN ACCIÓN INTRODUCCIÓN • El metabolismo es un circuito integrado de reacciones químicas la célula transforma los nutrientes captados en: Energía (ATP) Moléculas de recambio Nuevas moléculas Trabajo mecánico Síntesis de bio-macromoléculas Transporte activo Todos los seres vivos necesitan materia y energía para vivir Renovarse Crecer 3 TIPOS DE NUTRICIÓN AUTÓTROF Fuente de carbono: CO2 Fuente de energía : luz Fuente de energía : reacciones redox FOTOSÍNTESIS QUIMIOSÍNTESIS HETERÓTROF Fuente de carbono y energía: moléculas orgánicas 4 ORGANISMO FUENTE DE CARBONO FUENTE DE ENERGÍA EJEMPLOS FOTOLITÓTROFO CO2 Luz Células vegetales, algas verdes, bacterias fotosintéticas FOTOORGANÓTROFO Compuesto s orgánicos Luz Bacterias púrpura no sulfuradas QUIMIOLITÓTROFO CO2 Reacciones redox Bacterias desnitrificantes del hierro y del azufre QUIMIORGANÓTROFO Compuesto s orgánicos Reacciones redox Animales, hongos y m.o. METABOLISMO Reducción Oxidación NH3, CO2, Urea 6 Polisacáridos, lípidos, proteínas,... Reacciones exergónicas NADPH2 ! Reacciones endergónicas NADP 7 8 CARACTERÍSTICAS Y FUNCIONES Las reacciones metabólicas están acopladas energéticamente mediante el ATP ADP ! ATP Fosforilación a nivel de sustrato: se forma un compuesto intermedio “rico en energía” Ac.1,3 diP glicérico Ac.3P glicérico ADP ATP • Fosforilación en el transporte de electrones Los coenzimas reducidos NADH, FADH y NADPH ceden los electrones a otras moléculas hasta la molécula final aceptora. La energía transferida a una cadena de electrones es utilizada por la ATPasa, para la síntesis de ATP. La fosforilación oxidativa (O2 último aceptor de e-) membrana interna mitocondrial La fosforilación fotosintética (luz impulsa el transporte de e-) membrana tilacoidal de cloroplastos Las reacciones metabólicas son reacciones de óxido-reducción Los coenzimas transportan H+ y electrones de alto potencial energético desde las reacciones catabólicas a las anabólicas Compuesto orgánico Deshidrogenación reducido Compuesto orgánico oxidado Catabolismo NADP,NAD,FMN NADPH2, NAD+H+, FMNH2 Anabolismo Compuesto biosintético Hidrogenación reducido Compuesto biosintético oxidado Las reacciones metabólicas están ligadas entre sí y catalizadas por enzimas RUTAS METABÓLICAS: sucesión de reacciones encadenadas ( el producto de una es el sustrato de otra) en las cuales se producen metabolitos y cada una está catalizada por una enzima diferente ! FASE I Las rutas catabólicas son convergentes FASE III Ruta anfibólica: aquella que se puede utilizar tanto con fines anabólicos y catabólicos (C. de Krebs) FASE II Las rutas anabólicas son divergentes En rojo: rutas anabólicas En negro. rutas catabólicas Las reacciones metabólicas están compartimentadas La gran cantidad de reacciones simultáneas que tienen lugar en la célula ocurren en compartimentos diferentes, de esta manera se aumenta la eficacia enzimática ( más fácil que E y S interactúen) - En el citoplasma: glucolisis, gluconeogénesis - En la mitocondria: ciclo de Krebs, beta oxidación de ac.grasos - En el R.E.: síntesis de lípidos y proteínas - En el núcleo: duplicación y transcripción 14 TIPOS DE CATABOLISMO Respiración celular El dador es un compuesto orgánico y el aceptor final de electrones un compuesto inorgánico O2, NO3, SO4 Síntesis de ATP en la cadena de transporte de electrones (fosforilación oxidativa) Oxidación total (hasta CO2) . Se libera mucha energía R. Aerobia: aceptor de hidrógenos es el O2 R Anaerobia: aceptor es S, NO3, SO4,... Fermentación Dador y aceptor final de electrones una molécula orgánica pequeña (ac. pirúvico) Oxidación parcial, se libera poca energía Síntesis de ATP a nivel de sustrato. No hay cadena de trasnporte de electrones, ni fosforilación oxidativa. Levaduras, bacterias y células animales y vegetales en condiciones especiales Tipos de células según su catabolismo Células aerobias Necesitan el O2 para realizar la respiración aeróbica Células anerobias estrictas El O2 es tóxico. Realizan las fermentaciones y respiración anaerobia Células anerobias facultativas Pueden vivir con y sin O2. Pueden hacer fermentaciones y 17 respiración aerobia según la disponibilidad del O2 CATABOLISMO DE LOS GLÚCIDOS Reacciones para producir energía, tienen carácter oxidativo GLUCOLISIS Reacción anaeróbica,catabólica oxidativa. La glucosa proviene de la digestión, de las reservas almacenadas en el organismo o de la fotosíntesis No requiere oxígeno Proporciona 6 precursores metabólicos, 2 ATP y poder reductor Tiene lugar en el hialoplasma de las células La llevan a cabo todos los seres vivos (aerobios y anaerobios) Glucosa + 2NAD+ + 2ADP + 2Pi 2 NADH + 2H+ + 2H2 O 2 Piruvato + 2ATP + 1 1º Etapa de activación: La glucosa se rompe en dos triosas: se gastan 2 ATP 2º Etapa de degradación.Mediante reacciones de oxidación se transforma en dos moléculas de ac. pirúvico. El coenzima NAD+ se reduce a NADH +H+. La energía liberada se utiliza para fabricar ATP (4 moléculas) Para que el proceso no se detenga hay que aportar continuamente glucosa o algún compuesto intermedio y oxidar el NADH 2 2 2 2 2 La glucosa reacciona con el ATP y se forma glucosa 6-fosfato La glucosa 6-fosfato se isomeriza a fructosa 6-fosfato La fructosa 6-fosfato reacciona con el ATP y da fructosa 1,6 difosfato La fructosa 1, 6-difosfato se rompe y da lugar al aldehído 3, fosfoglicérico y la dihidroxiacetonafosfato El aldehído 3 fosfoglicérico se oxida gracias al coenzima NAD+ y se fosforila por el ácido fosfórico dando ácido 1,3 difosfoglicérico 2 2 2 2 El ácido 1,3 difosfoglicérico reacciona con el ADP para dar ATP y y ácido 3 fosfoglicérico 2 2 2 2 Fosforilación a nivel de sustrato El ácido 3- fosfoglicérico reacciona con el ADP y se forma ATP y ácido pirúvico 2 2 2 2 BALANCE GLOBAL DE LA GLUCOLISIS Glucosa + 2NAD+ + 2ADP + 2Pi 2 Piruvato + 2ATP + 2 NADH+ 2H+ + +2H2 O GLUCOSA Oxidación vía glucolisis DESTINO DEL PIRUVATO PIRUVATO Condiciones aerobias R.AEROBIA ACETIL CoA CO2 + H2O Condiciones anaerobias FERMENTACIONES LACTATO ETANOL + CO2 FERMENTACIÓN Vía catabó lica anaer obia Citosol o h ialoplasm a El H+ es cedido a un sustrato orgánico La oxidación es parcial, se obtiene poca energía(2ATP/glc) Los productos finales son ac. orgánicos y etanol La síntesis de ATP a nivel de sustrato Es propia de microorganismos (bacterias y levaduras) y cuando no hay suficiente O2 en las células musculares FERMENTACIÓN LÁCTICA St La ct ob ac ill to co us s cc us p sp rep ✴Se utiliza a nivel industrial para la obtención de yogurt, quesos,… ✴Las bacterias fermentan la lactosa y la bajada de pH desnaturaliza la caseína que precipita 2 2 2 2 Citosol 2 2 Cuando no llega suficiente O2 a las células musculares, La glucosa se degrada a ac.láctico FERMENTACIÓN ALCOHÓLICA El ac. pirúvico se transforma en alcohol etílico o etanol. Proceso llevado a cabo por levaduras del género Saccharomyces. Se utiliza para la fabricación de bebidas alcohólicas (cerveza, vino, sidra,...) En la fabricación del pan la fermentación del almidón de la harina hace que el pan sea esponjoso por las burbujas de CO2. El alcohol producido desaparece durante el tiempo de cocción. La fermentación alcohólica tiene el mismo objetivo que la fermentación láctica: la recuperación del NAD+ en condiciones anaeróbicas 2 2 2 2 (acetaldehído) 2 2 Citosol 2 2 2 2 RESPIRACIÓN AEROBIA DESTINO DEL PIRUVATO Tiene lugar en la mitocondrias. Las enzimas de la cadena respiratoria se localizan en la membrana interna Glucosa Glucolisis(hialoplasma) 2 Piruvato Oxidación vía pentosas fosfato (mitocondria) Ribosa 5-P NADPH Descarboxilación oxidativa 2 Ac.acetil-CoA Ciclo de Krebs Transporte de electrones (Membrana interna mitocondrial) Matriz mitocondrial Descarboxilación oxidativa s a l e o d t a a Ví s fosf a s o t n e p 37 Formación del acetil CoA (Descarboxilación oxidativa) 2 2 Piruvato descarboxilasa 2 CO2 2 2 Piruvato + NAD+ + HS-CoA ------> Acetil-CoA + CO2 + NADH + H+ CICLO DE KREBS O CICLO DEL ÁCIDO CÍTRICO CICLO DE KREBS CO2 CO2 Fosforilación a nivel de sustrato 1) El acetil-CoA se une al ácido oxalacético y forma el ácido cítrico. en este proceo se recupera el CoA-SH 2) El ácido cítrico se isomeriza a ácido isocítrico 3) Descarboxilación del ácido isocítrico que se transforma en ac. alfa-cetoglutárico, liberándose CO2 y NADH 4) Descarboxilación del ac. alfa-cetoglutárico, se libera CO2, NADH + H + y se forma 1 GTP (ATP). El ac. alfa cetoglutárico se transforma en ac. succínico 5) Oxidación del ac. succínico a ac. fumárico. Se forma un doble enlace. Los electrones son transferidos al FAD que se reduce a FADH2. 6) Adición de agua al doble enlace formándose el ac. málico. 7) Oxidación del alcohol del ac. málico a ac. oxalacético. EL NAD+ se reduce a NADH. el ciclo se completa Acetil CoA C2 CoA Ac. Oxalacético C4 NADH2 C6 Ac. cítrico NADH2 FADH2 C. de KREBS CO2 GTP CoA C4 C5 Ac. Α cetoglutárico Succinil CoA ATP CO2 NADH2 Balance global del Ciclo de Krebs Acetil-CoA + 3 NAD+ + FAD + GDP + Pi + 2H2O --------> 2CO2 + CoA-SH + 3(NADH + H+) + FADH2 + GTP Acetil CoA CoA 2CO2 ATP 3 NADH2 FADH2 Electrones de alta energía ATP Cadena transporte de electrones Oxígeno v Intermediarios metabólicos Biosintéticas precursores Rutas CADENA DE TRANSPORTE DE ELECTRONES - Se sitúa en la membrana interna mitocondrial (c.eucariotas) Los electrones y los H+ de los coenzimas reducidos (NADH2 y FADH2 ) se transfieren a uno de los complejos proteicos que forman la cadena transportadora de electrones, hasta llegar al O2 Son reacciones REDOX, en las cuales intervienen dos constituyentes de diferente potencial redox. Los electrones van desde la molécula con menor potencial redox a la molécula con mayor potencial redox. ESTRUCTURA DE LA MEMBRANA DE LAS CRESTAS MITOCONDRIALES La membrana de las crestas mitocondriales tiene la estructura de membrana de mosaico fluido. Empotradas en la doble bicapa lipídica se encuentran las proteínas transportadoras de los electrones. Forman tres grandes complejos: . Complejo I (NADH deshidrogenasa) - Complejo II (citocromo bc1) - Complejo III (citocromo oxidasa) El transporte de electrones comienza cuando el NADH + H+ cede los e- al complejo I y a través del Co Q, pasan al ¿CÓMO SE2 PRODUCE EL TRANSPORTE complejo II. El FADH (menor potencial redox) DElos ELECTRONES? cede directmente al complejo II. Luego se tranfieren a través del citocromo C al complejo II y por último son aceptados por el oxígeno (mayaor potencial redox)que se reduce a agua. 2H+ + 1/2 O2 + 2e - H 2O Mecanismo de la cadena respiratoria. Oxidación del NADH y síntesis de ATP Mecanismo de la cadena respiratoria. Oxidación del FADH2 y síntesis de ATP HIPÓTESIS QUIMIOSMÓTICA Acoplamiento de los procesos redox y trasnporte de H+ A la vez que se realiza el transporte de electrones se genera un transporte de protones por parte de los complejos I, II y III desde la matriz hacia el espacio intermembranoso Como la membrana interna es impermeble a los H+, se genera un gradiente electroquímico entre el espacio intermembranoso y la matriz mitocondrial,que es aprovechado por la bomba ATP-asa para devolver de nuevo los protones y generar ATP( por cada dos protones que retornan se sintetiza un ATP). Se calcula que el NADH cede 2 electrones y en su transporte se libera energía para bombear 6H+ ( por cada FADH2 se bombean 4 H+). De manera que : 1NADH+H+/3ATP 1 FADH2/2ATP La hipótesis que explica el acoplamiento de los procesos redox y el transporte de H+ es la hipótesis quimiosmótica, y su resultado es la fosforilación oxidativa. BALANCE EN MOLES DE ATP EN LA RESPIRACIÓN CELULAR ( Una molécula de glucosa) EUCARIOTAS PROCARIOTAS Coenzimas reducidos y ATP Moles de ATP (totales) Moles de ATP (totales) 2 NADH 2 ATP 4 ATP 2 ATP 6 ATP 2 ATP Descarboxilación 2 NADH del ac. pirúvico 6 ATP 6 ATP 18 ATP 4 ATP 2 ATP 18 ATP 4 ATP 2 ATP 36 ATP 38 ATP Proceso Glucolisis C. de Krebs Balance global 6 NADH 2 FADH2 2 GTP ¿QUÉ SUCEDE CON EL NADH DE ORIGEN HIALOPLASMÁTICO EN LOS EUCARIOTAS? Hemos visto que por cada NADH que ingresa en la cadena de electrones se obtienen 3 ATP. En los organismos eucariotas, el NADH que se origina en la glucolisis (hialoplasma) por cada glucosa sólo puede originar 2 ATP. Esto es debido a que el NADH no puede atravesar directamente la membrana mitocondrial y utiliza una lanzadera que cede los electrones al FAD que hay en el interior de la mitocondria, reduciéndose a FADH2. Esto no sucede en procariotas. BALANCE DE LA RESPIRACIÓN AEROBIA DE 1 GLUCOSA (c.eucariota) Proceso Sustancia inicial Sustancia final coenzimas reducidos y ATP Glucosa 2 ac. pirúvico 2 NADH 2 ATP 4 ATP 2 ATP Descarboxilación 2 ac. del ac. pirúvico pirúvico 2 acetilCoA 2 CO2 2 NADH 6 ATP 4 CO2 6 NADH 2 FADH2 2 GTP 18 ATP 4 ATP 2 ATP Glucolisis C. de KREBS Balance global C6H12O6 + 6O2 2 acetilCoA Glucosa 6 CO2 + 636ADP + 36Pi O2 6 H2O Moles de ATP (totales) 36 ATP 6CO2 + 6H2O + 36 ATP ESQUEMA SIMPLIFICADO DE LOS PROCESOS DE RESPIRACIÓN CELULAR 6 6 2 2 ATP NADH2 36 2 CO2 4 2 NADH2 2NADH2 6NADH2 2FADH2 6 6 61 Si comparamos ambos procesos: catabolismo aeróbico y fermentación vemos que el rendimiento del primero es mayor ( 38 ATP por molécula de glucosa) que el del segundo (2 ATP por mol. de glucosa). Evolutivamente, el mayor rendimiento energético de los organismos aeróbicos fue muy importante ya que les permitó conquistar todos los ambientes, produciéndose una gran variedad de formas de vida, cada una adaptada a un nicho ecológico determinado, es lo que se llama radiación adaptativa. RESERVA DE GLÚCIDOS La concentración de glucosa en sangre debe permanecer constante. El exceso de glucosa se acumula en hígado y músculo (animales) en forma de glucógeno. Al completarse esta reserva el resto de glucosa se almacena en forma de grasa. - El glucógeno del músculo aporta el combustible a las células musculares para su contracción. - El glucógeno hepático suministra glucosa al resto de los tejidos ( en ayuno esta reserva se agot en menos de 24h). La principal reserva energética la proporcionan los lípidos que se almacenan en el tejido adiposo (9Kcal/g) CATABOLISMO DE LOS LÍPIDOS En los animales los ac. grasos se almacenan en el tejido adiposo en forma de triglicéridos y se movilizan gracias a las lipasas. Lipasas Citoplasma Ingresa en la glucolisis como gliceraldehído 3 P Matriz mitocondrial Citoplasma 1 ATP R-COOH + CoA- SH Ac. Graso Cadena transporte de electrones R-COS- CoA AcilCoA Acil-CoA Membran externa sintetasa mitocondrial C. de Krebs 1 AMP - 1 P+ 1Pi Para atravesar la membrana mitodcondrial utiliza como lanzadera la carnitina n Acetil-CoA e- H+ e- H+ NADH2 FADH2 Ac. graso (18 C) Hélice de Lynen (8 vueltas) β- Oxidación ó Hélice de Lynen (Matriz mitocondrial y peroxisomas). Vegetales: peroxisomas 146 ATP En los animales constituyen la principal reserva energética CATABOLISMO DE LAS PROTEÍNAS ! Transaminación Proteínas de la dieta -NH2 Α-cetoglutarato AMINOÁCIDOS Transaminasas Coenzima B6 C. KREBS Preferentemente en el hígado Desaminación oxidativa Glutamato dh NH3 Hígado y riñones NH3 Ac. úrico Proteínas corporales Ciclo de Hígado la urea (citosol y mitocondrias) Urea Biosíntesis Nucleótidos Coenzimas Neurotransmisores Hormonas Porfirinas Oxidación de la cadena carbonada Esqueleto carbonado C. de Krebs GLUCOSA ÁCIDOS GRASOS CO2 , H2 O energía POLISACÁRIDOS LÍPIDOS PROTEÍNAS 3º 1º Ac grasos Glicerina Aminoácidos 2º 2º Acetil-CoA CO2 3º ATP H 2O 1º H y eTransportadores de electrones O2 NH3 Anabolismo Catabolismo Pentosas Hexosas ESQUEMA GENERAL DEL CATABOLISMO NADH CO2