citología ii - Colegio Maravillas
Anuncio

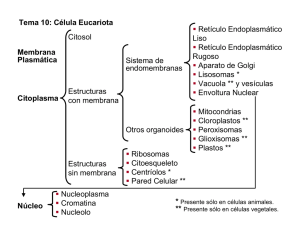
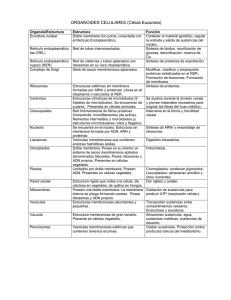
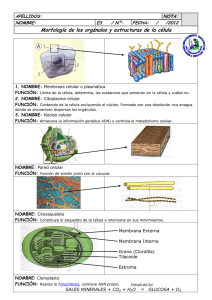
CITOLOGÍA II 3. 3 Citosol y ribosomas. Citoesqueleto. Centrosomas. Cilios y flagelos. CITOESQUELETO. El citoesqueleto es propio de las células eucarióticas. Está compuesto por proteínas que forman estructuras filamentosas, dando lugar a una red tridimensional dinámica que se extiende a través del citoplasma. Esta red está continuamente organizándose y desorganizándose, en un constante dinamismo de ensamblaje de las proteínas que lo conforman. Estructura y funciones Existen 3 tipos de filamentos que, según su grosor y constitución química, se denominan: microfilamentos, filamentos intermedios y microtúbulos Los microfilamentos: Fibras delgadas y flexibles que tienen un diámetro de unos 5 a 7 nm Están formadas por una proteína globular llamada actina. Funciones: Contracción muscular: interacción de la actina con la miosina. (células musculares) Formación de extensiones o protuberancias de la célula: aumentan la superficie de absorción (microvellosidades intestinales). Formación del anillo contráctil de estrangulación durante la citocinesis: para separar las dos células durante la mitosis. Formación de pseudópodos: movimiento ameboide. Corrientes citoplasmáticas: que permiten desplazar los orgánulos por el citoplasma. Filamentos intermedios: Son filamentos de proteína fibrosa que van de 8 a 11nm de diámetro, con un grosor intermedio entre los microfilamentos y los microtúbulos. Son los componentes del citoesqueleto más estables, dando soporte a los orgánulos y proporcionan resistencia estructural a la célula. Las proteínas que conforman estos filamentos dependen del tejido en el que se hallen (queratinas: en células epiteliales; proteínas de neurofilamentos: en neuronas; vicentinas: en tejidos conjuntivos o leucocitos…) Microtúbulos: son cilindros huecos y rígidos de 25 nm de diámetro, son los componentes de mayor grosor. Están compuestos por unas proteínas denominadas tubulinas (alfa, beta y en el centrosoma también la gamma). Se pueden polimerizar y despolimerizar según las necesidades de la célula. Los microtúbulos son más flexibles pero más duros que la actina. Funciones: Movimiento de orgánulos y vesículas. Formación del huso acromático. Movimiento celular. Las células eucariotas presentan en su citoplasma un centro organizador de los microtúbulos: El centrosoma. Centrosomas Un centrosoma es un centro organizador de los microtúbulos que se encuentra en las células animales; se localiza en el citoplasma cerca del núcleo. Forman una parte del citoesqueleto de la célula. Su composición sigue este orden (desde lo más grande hasta lo más pequeño): Los centrosomas están compuestos por diplosomas que a su vez lo componen dos centriolos. Los centriolos constan de microtúbulos y áster. Los microtúbulos son proteínas cilíndricas compuestas por subunidades llamadas tubulina. Esta se divide en tres clases distintas: α β γ. El áster es una estructura que existe en el medio de los microtúbulos. Un centriolo lo forman 9 grupos de 3 microtúbulos cada uno. La estructura es la siguiente: A los grupos de 3 microtúbulos (A, B, C) se les denomina triplete. El espacio central donde se encuentra el áster se llama eje tubular. Los centriolos se encuentran enfrentados perpendicularmente. Cuando la tubulina no se encuentra formando los microtúbulos, están dispersas en el citoplasma, pero siempre disponibles para una futura formación de dichas estructuras. Estos centros organizadores se caracterizan por tres funciones: movimiento, transporte y la formación del huso mitótico. El transporte se produce por medio de la adhesión de una vesícula a una proteína, y esta se une al microtúbulo. A esto se le denomina motor celular. Durante el proceso de división celular (exactamente en la interfase) los centrosomas se replican dando lugar a dos centrosomas con su par de centriolos. Estos se desplazan hasta situarse a lados opuestos de la célula, entonces de cada uno surge un racimo de filamentos denominados áster. Posteriormente, se forma el huso mitótico entre los centriolos por medio de los filamentos. Estos filamentos están compuestos de proteína y por pequeñas cantidades de ácido ribonucleico. Los cromosomas se adhieren a estos filamentos por el centrómero. En cuanto al movimiento celular, necesita un aparato locomotor intracelular que le ayude a desplazarse y detenerse para poder relacionarse con su alrededor. La creación de cuerpos basales (cilios o flagelos) en las membranas celulares le aporta movilidad. El desplazamiento de las proteínas específicas por el microtúbulo se hace en direcciones opuestas. También se cree que estos orgánulos están relacionados con la movilidad y localización de los demás orgánulos principales internos dentro del citoplasma. Transporte de vesículas por medio de los microtúbulos Cilios y flagelos. Los cilios y flagelos son prolongaciones de la membrana plasmática formadas por microtúbulos y proteínas asociadas, responsables del movimiento de ciertos tipos celulares. Tienen una estructura común, pero se diferencian en su patrón de movimiento, y en que los cilios son numerosos y cortos y los flagelos son pocos, más largos y más gruesos debido a que tienen otro tipo de estructuras añadidas, como mitocondrias o fibras. El estudio de estas estructuras se basa en: • Morfología y composición. Estructura y funcionamiento. Función. Cilios. Del latín “cilium”, que significa pestaña, son unos pequeños apéndices motores propios de células desnudas o sin pared, que pueden cubrir, de manera total (a modo de césped) o de manera parcial la superficie celular y, son responsables del movimiento de la célula. Morfología y composición Como ya ha sido mencionado antes, son unos apéndices móviles, cuya membrana está formada por proteínas (más de 250 proteínas diferentes) y lípidos. Dentro de la membrana tiene una serie de filamentos proteicos responsables del movimiento en sí. Estructura y funcionamiento. A menudo son definidos como evaginaciones de la membrana plasmática que albergan contenido citoplasmático. Tienen una forma cilíndrica con el mismo diámetro en toda su extensión, y acaban en una punta redondeada. Consta de varias partes: la primera y más fundamental es el anoxema, que es el esqueleto interno (cuando está bajo la membrana plasmática se denomina cuerpo basal) y consta de 9 microtúbulos exteriores que rodean a 2 microtúbulos interiores, que forman el conocido patrón de ‘9+2’. En el corpúsculo basal, los periféricos tienen 9 fibras densas asociados a ellos que intervienen en el movimiento. Los dos microtúbulos internos están rodeados por una vaina interna. Los microtúbulos periféricos están formados por uno completo (A) y otro incompleto (B), mientras que los dos centrales son completos. Los periféricos están unidos a los centrales por filamentos radiales, y los periféricos están unidos entre sí gracias a una proteína llamada nexina. También entra en juego otra proteína llamada dineína que sale a modo de dos brazos del microtúbulo periférico A, y que es la encargada de transformar la reacción ATPADP+P en trabajo mecánico. En la parte basal (dentro de la membrana plasmática) el par central desaparece y se observan 3 tubos por cada par que había antes de microtúbulos periféricos, ya que se le une una fibra a cada par asociada con el movimiento. Función El funcionamiento se basa en el deslizamiento de los dobletes externos, unos respecto a otros. Aquí entra en juego la dineína . En presencia de ATP, los brazos de dineína del microtúbulo A se deslizan por encima del microtúbulo B contiguo. Como los microtúbulos están unidos entre sí gracias a la nexina, esto provoca la flexión de los microtúbulos. Esta flexión es la base del batido de cilios y flagelos. Este movimiento tiene dos momentos: el primero es el del golpe efectivo, en el que se flexionan los cilios. El segundo es denominado ‘fase de recuperación’, y ocurre después de que los flagelos hayan producido su golpe efectivo y vuelven a prepararse para realizarlo. Es importante conocer el mecanismo de coordinación de los cilios. Entra el juego el metacronismo. La coordinación de los cilios entre sí, al fustigar el agua sobre la superficie de una célula, viene dada por la misma agua, movida por el cilio precedente. El que sigue en fila halla así una dirección favorecida y se mueve por ella con un pequeño retraso. Su función básica es la del movimiento de las células. Se mueven a forma de remo en el agua mediante las dos fases anteriormente dichas. Algunos cilios como los de los bronquiolos en la garganta de nuestro organismo se encargan de captar partículas perjudiciales que hay en el aire. Flagelos Morfología y composición. En cuanto a filamento y su membrana es exactamente igual que los cilios, pero en su parte basal posee un motor molecular compuesto por diferentes tipos de proteínas (anillos S, M, C, L Y P). También están compuestos por fibras que contienen flagelina. Son más largos y gruesos que los cilios. Dependiendo de su situación, encontramos cuatro casos que se pueden ver en la 2 imagen (A)Monótrica, B)Lofótrica, C)Anfítrica, D)Perítrica. Funcionamiento. El estator, formado por el anillo C y los pasadores, se ensambla con la membrana plasmática y establece el motor molecular. El rotor, formado por los anillos M y S, gracias a la hidrólisis del ATP en ADP + energía que se utiliza para producir el movimiento, empieza a funcionar y a girar y hace que gire el cojinete constituido por los anillos L y P. El codo o gancho es el encargado de transmitir el movimiento al filamento, que se moverá helicoidalmente. La diferencia fundamental en el movimiento de en cilios y flagelos es el patrón de movimiento, ya que los flagelos se mueven helicoidalmente y los cilios de atrás hacia delante. Ribosomas Estructura y composición Este orgánulo está formado por dos subunidades (una grande y otra pequeña) formadas por ARN ribosómico y proteínas. El espacio que queda entre las dos subunidades tiene componentes enzimáticos que colaborarán con el ARNr y las proteínas en la síntesis proteica. Son orgánulos SIN membrana. Se encuentran tanto en procariotas como en eucariotas, diferenciándose en tamaño y número de proteínas. El tamaño viene determinado por el coeficiente de sedimentación (unidades Svedberg). Este coeficiente nos da una idea aproximada del peso y volumen del orgánulo. Los cromosomas eucariotas miden 80S y los procariotas 70S. Localización Cada subunidad se forma en el núcleo, concretamente en el nucleolo, pero son exportadas separadas al citoplasma, donde estas dos subunidades se ensamblan dando lugar a un ribosoma funcional. Pueden encontrarse de forma libre formando polirribosomas (leen el ARNm); asociados al retículo endoplasmático o a la membrana externa de la envoltura nuclear; también pueden encontrarse dentro de las mitocondrias y cloroplastos, en este caso de r. de 70S (procariotas) ya que las mitocondrias y plastos tienen origen bacteriano (Teoría de Lyn Margulis). Función biológica Su principal función es la biosíntesis de proteínas. Los ribosomas son los encargados de traducir el mensaje genético aportado por el ARNm transformándolo en las cadenas de aminoácidos que forman las proteínas. Para ello los ribosomas: • Permiten el acoplamiento del ARNm junto al ARNt por la complementariedad de las bases del codón y anticodón. • Hacen posible la formación del enlace peptídico entre los aminoácidos aportados por el ARNt al estar presente el enzima péptido-sintasa. • Actúan en la iniciación, elongación y terminación de la cadena de aminoácidos, dando lugar a una proteína. El ribosoma es la principal diana de algunos antibióticos. Estos se unen a algunas de las subunidades del ribosoma o interfirieren en la síntesis de proteínas en bacterias, ejerciendo un efecto bactericida. 3.4. Orgánulos celulares: mitocondrias, peroxisomas, cloroplastos, retículo endoplasmático, Complejo de Golgi, lisosomas y vacuolas. Mitocondrias Son orgánulos celulares presentes en todas las células eucariotas aerobias. Están implicadas en la mayor parte de las reacciones oxidativas celulares y obtienen de ellas energía que almacenan sintetizado ATP. Su forma es variable, aunque suelen tener una forma de cilindro alargado. El número de mitocondrias por célula es variable, dependiendo de su actividad metabólica. Estructura: - - - Membrana externa: tiene una composición similar de proteínas y lípidos y parece bastante permeable, debido fundamentalmente a las porinas que poseen canales de paso. Membrana interna: muy rica en proteínas (70-80% de su composición) y manifiesta una elevada impermeabilidad debido al tipo de lípidos que posee, y a la ausencia de porinas entre sus proteínas. Esta membrana selecciona las sustancias que van a pasar a la cámara interna. presenta numerosos pliegues denominados crestas. Espacio intermembranoso (cámara externa): relleno de un líquido bastante parecido al citosol, aunque con algunas proteínas específicas. Matriz mitocondrial (cámara interna): contiene ADN, ARN y ribosomas propios. Incluye enzimas responsables del ciclo de Krebs y transportadores de electrones. Funciones: - Ciclo de Krebs o de los ácidos tricarboxílicos: ruta del catabolismo de los glúcidos. Se produce en la cámara interna. - Fosforilación oxidativa: Gracias a la cadena de transportadores de electronesprotones en la cadena respiratoria se produce la síntesis de ATP. Se produce en las crestas de la membrana interna. - β-oxidación de los ácidos grasos: catabolismo de lípidos. Se da en la cámara interna. Funciones propias como replicación y transcripción de su ADN y síntesis de sus propias proteínas. Cámara interna. Además, las mitocondrias se dividen, independientemente de la célula, por escisión binaria. Cuando la célula se divide, las mitocondrias se reparten entre las dos células hijas. Perixosomas Son orgánulos citoplasmáticos en forma de vesícula, rodeados de una membrana simple y en cuyo interior se encuentran enzimas oxidativas, donde destaca el enzima catalasa, a veces la concentración de enzimas cristalizadas se puede apreciar a microscopía electrónica. Las enzimas oxidativas intervienen en el metabolismo energético y la catalasa degrada el peróxido de hidrógeno, formando agua y oxígeno. Se replican por división, como las mitocondrias y los cloroplastos, aunque a diferencia de ellos carecen de ADN o de ribosomas propios. Funciones: - β-oxidación de los ácidos grasos, procesos en los que se genera H2O2. - Eliminación del agua oxigenada gracias a la catalasa o usándola para oxidar otras sustancias. - Detoxificación de diversas sustancias. - Síntesis de fosfolípidos, especialmente en el cerebro y en el corazón. Degradación de las bases nitrogenadas y de lípidos. En las células vegetales, los peroxisomas están implicados, además, en: Fotorrespiración: Una de las propiedades más interesantes de la enzima RUBISCO es que además de catalizar la carboxilación de la ribulosa 1,5 bifosfato, también produce su oxigenación; proceso conocido como fotorrespiración: RUBP + O2 + rubisco Ácido fosfoglicérico + Glicolato. La fotorrespiración da como resultado la liberación de CO2, después de una serie de reacciones enzimáticas. (Lo veremos en el capítulo de metabolismo) Ciclo de glioxilato (en las células de las semillas), por el que los ácidos grasos almacenados se convierten en azúcares (ciclo parecido al Krebs). En este caso los peroxisomas se denominan glioxisomas. CLOROPLASTOS: ESTRUCTURA Y COMPOSICIÓN. Son los plastos más complejos, son exclusivos de las células vegetales y bacterias fotosintéticas. Poseen un color verde debido al espectro de absorción de la clorofila, reflejando la onda luminosa que no absorbe (verde) Su forma más frecuente es lenticular, ovoide o esférico. Morfología: Envoltura externa: rodea al cloroplasto y está formada por una membrana externa y otra interna, separadas por un pequeño espacio denominado espacio intermembranoso. Ambas membranas son lisas y no contienen clorofila. La membrana externa es muy permeable, mientras que la interna es casi impermeable, contiene numerosas proteínas transportadoras. Membrana tilacoidal: impermeable a la mayoría de moléculas e iones. Es plegada y se encuentra en el interior del cloroplasto (estroma). Rodea a un espacio interno llamado espacio tilacoidal y al replegarse, forma sacos aplanados denominados tilacoides, que se comunican entre sí. Los apilamientos de tilacoides reciben el nombre de grana (unidos entre sí por estructuras denominadas lamelas). Los tilacoides son vesículas en cuya membrana se encuentran las moléculas que forman los fotosistemas. En el interior de los tilacoides se encuentra la clorofila. En el estroma se encuentra el ADN circular y ribosomas 70S y pueden aparecer gránulos de almidón. Esta membrana está compuesta por: o o o 38% de lípidos 50% de proteínas, las cuales pueden ser de tres tipos: - asociadas a pigmentos fotosintéticos (formando complejos moleculares denominados fotosistemas I y II), - transportadoras de electrones (desde un dador que suele ser el agua hasta el NADP que los capta y reduce) - otros complejos enzimáticos. 12% de pigmentos fotosintéticos que absorben la energía solar. Son de dos tipos: clorofilas y carotenoides (terpenos). En los cloroplastos se diferencian tres compartimentos: espacio intermembranoso, estroma y espacio tilacoidal. Estroma: parte interna del cloroplasto formado por una molécula de ADN bicatenario y circular que llevan información para la síntesis de proteínas del cloroplasto (la mayor parte de sintetizan en el hialoplasma), los ribosomas y enzimas, los cuales pueden intervenir en la replicación, transcripción y traducción del ADN, o pueden ser responsables de la fase oscura de la fotosíntesis. Funciones FOTOSÍNTESIS: proceso por el cual se sintetiza materia orgánica a partir de la inorgánica, utilizando para ello la energía solar. En este proceso se libera oxígeno molecular y tiene como ecuación global: En presencia de clorofila En este proceso se diferencian dos etapas: • • Fase luminosa, ocurre en la membrana tilacoidal y se necesita energía luminosa. Los cloroplastos de las células vegetales captan por medio de la clorofila la energía lumínica, se fractura la molécula de agua, se libera oxígeno y se liberan protones que reducen la nicotidamina (NADH) y ATP. Estas moléculas aportarán, en la fase siguiente, energía química para la síntesis de glúcidos. Fase oscura, ocurre en el estroma, no se necesita la luz. SE PRODUCE TANTO DE DÍA COMO DE NOCHE. En esta etapa esas moléculas intervienen en la reducción del CO2 mediante una serie de reacciones: el ciclo de Calvin, que lleva a la formación de la glucosa. Este azúcar se disuelve en agua y recorre toda la planta proporcionándole la energía necesaria para crecer. SÍNTESIS DE PROTEÍNAS: En el estroma de los cloroplastos se sintetizan las proteínas del cloroplasto que están codificadas por el ADN del cloroplasto, que tan solo representan una pequeña parte la mayoría se sintetizan en el hialoplasma. FOTORRESPIRACIÓN, junto con los peroxisomas. FORMACIÓN DE AMONIACO, a partir de nitratos y nitritos. SÍNTESIS DE ADN Retículo endoplasmático El retículo endoplasmático se diferencia en dos tipos de sistemas membranosos de distintas características estructurales y funcionales. Su aportación a la vida es fundamental y gracias a su funcionamiento se sintetizan y distribuyen sustancias imprescindibles para el correcto funcionamiento metabólico de la célula. Se encuentra en la célula vegetal y animal pero nunca en la procariota. ESTRUCTURA El retículo endoplasmático es un complejo sistema de membranas dispuestas en forma de sacos aplanados y túbulos que están interconectados entre sí, compartiendo el mismo espacio interno. Sus membranas se continúan con las de la envuelta nuclear y se pueden extender hasta las proximidades de la membrana plasmática, llegando a representar más de la mitad de las membranas de una célula. • El retículo endoplasmático rugoso tiene esa apariencia debido a los numerosos ribosomas adheridos a su membrana mediante unas proteínas denominadas "riboforinas". Tiene unos sáculos más redondeados, cuyo interior se conoce como "luz del retículo" o "lumen" donde caen las proteínas sintetizadas en él. Está muy desarrollado en las células que, por su función, deben realizar una activa labor de síntesis, como las células hepáticas o las células del páncreas. • El retículo endoplasmático liso no tiene ribosomas y participa en el metabolismo de lípidos. FUNCIONES: Biosíntesis proteica: El ARNm proviene de la transcripción del ADN nuclear y es su “imagen especular”. Al llegar al retículo endoplasmático, se fija a unas estructuras específicas llamadas ribosomas, adheridas al retículo endoplasmático. Allí participa en la síntesis de proteínas, determinando el orden en que se unirán los aminoácidos. Metabolismo de lípidos: Dado que no tiene ribosomas, en el retículo endoplasmático liso no se sintetizan proteínas. Pero tiene un papel esencial en la síntesis de lípidos de la membrana plasmática, colesterol y derivados de éste, como los ácidos biliares o las hormonas esteroideas. Destoxificación: Es un proceso que se lleva a cabo principalmente en las células del hígado y que consiste en la inactivación de productos tóxicos como drogas, medicamentos o los propios productos del metabolismo celular, por ser liposolubles. Glucoxilación: Son reacciones de transferencia de un oligosacárido a las proteínas sintetizadas. Se realiza en la membrana del retículo endoplasmático. De este modo, la proteína sintetizada se transforma en una proteína periférica externa del glucocálix en la reproducción de lisosomas. Movilización de glucosa: Cuando existe necesidad de glucosa en el organismo entre las comidas o durante el ejercicio muscular, las reservas hepáticas de este monosacárido almacenadas como inclusiones de glicógeno son movilizadas hacia la sangre. Almacenamiento y liberación de calcio: en el músculo estriado se necesita calcio para producir la contracción muscular. En este tipo de células recibe el nombre de Retículo Sarcoplásmico. Almacenamiento de la Glucosa-6-fosfatasa: es una proteína integral del retículo endoplasmático y que está ausente en otras células que almacenan glucógeno. Solo la encontramos en los REL del Hígado. Complejo de Golgi También llamado complejo de Golgi, es un orgánulo de membrana simple que está constituido por una o más pilas de vesículas membranosas aplastadas, denominadas sáculos. Morfológicamente el AG está compuesto por membranas aplanadas que encierran sacos y túbulos (cisternas) asociados con vesículas de secreción y de transición. La unidad básica del orgánulo es el sáculo, que consiste en una vesícula o cisterna aplanada. Cuando una serie de sáculos se apilan, forman un dictiosoma. Además, pueden observarse toda una serie de vesículas más o menos esféricas a ambos lados y entre los sáculos. El conjunto de todos los dictiosomas y vesículas constituye el aparato de Golgi. El aparato de Golgi esta subdividido en tres regiones o fases: la región cis: orientada hacia el núcleo celular, la región intermedia y la región trans; fase de maduración. En la cara cis se encuentran las vesículas de transición, mientras que en la cara trans, se localizan las vesículas de secreción. Las proteínas que vienen del RE entran al AG en su fase cis o fase de entrada por endocitosis, permitiendo la entrada de la proteína. Posteriormente por gemación, sale de la fase cis y entra a la intermedia. Nuevamente por gemación sale de la fase intermedia y entra a la fase trans. Finalmente, por gemación sale la proteína, generándose una vesícula que saldrá a un destino final. Esta última vesícula lleva el direccionamiento o la ruta que debe seguir para depositar la proteína que transporta. Una de las funciones más importantes de las vesículas es transportar materiales hacia la membrana plasmática y desde ella hacia el interior de la célula; constituyendo de este modo un medio de comunicación entre el interior y el exterior celular. Específicamente, hay un intercambio continuo de materiales entre el retículo endoplasmático, el aparato de Golgi, los lisosomas y el exterior celular. Dentro de las funciones que posee el aparato de Golgi se encuentran la glicosilación de las proteínas y lípidos para producir glucoproteínas y glucolípidos, selección (sorting), destinación (targeting) y la síntesis de polisacáridos de la matriz extracelular. El proceso de glicosilación, que la mayoría de las veces se inicia en el retículo endoplásmico es muy importante, pues permite darle a la molécula procesada propiedades especiales. En el caso de las proteínas, por ejemplo, su glicosilación da lugar a los componentes básicos del glucocálix (capa de oligosacáridos ubicada en la cara externa de la membrana plasmática), que posee un papel fundamental en procesos de comunicación celular y transducción de señales. En otros casos permite otorgarle a la molécula una resistencia mecánica adicional, como es común cuando se trata de hormonas o mensajeros a distancia. Desde la cisterna trans se originan vesículas con productos maduros, ya sea a la membrana plasmática o a otros orgánulos tales como los lisosomas. Cada una de las pilas de sáculos (cisternas) que integran el aparato de Golgi recibe el nombre de dictiosoma; cada dictiosoma agrupa unos 6 sáculos. El aparato de Golgi se encuentra estructural y funcionalmente polarizado. Presenta dos caras bien diferenciadas: la cara cis se sitúa próxima a las membranas del retículo endoplasmático y está rodeada de pequeñas vesículas, denominadas vesículas de transición, que derivan de él; la cara trans se encuentra próxima a la membrana plasmática y de ella parten unas vesículas más grandes, las vesículas secretoras. Existe un intenso tráfico de sustancias a través del aparato de Golgi que va desde la cara cis hacia la cara trans. Estas sustancias proceden del retículo endoplasmático, llegan a la cara cis en forma de vesículas de transición, van pasando de sáculo en sáculo, y por último salen por la cara trans en forma de vesículas secretoras que se dirigen hacia diferentes destinos celulares. Lisosomas y Vacuolas Los lisosomas son orgánulos de membrana simple que albergan en su interior enzimas hidrolíticos. Se trata de vesículas esféricas rodeada de una membrana. Los lisosomas participan en la digestión celular, y para ello contienen enzimas digestivas en su interior, que digieren la materia orgánica compleja, transformándola en moléculas más sencillas (polisacáridos en monosacáridos, proteínas en aminoácidos…). Se diferencian dos tipos de lisosomas: Lisosomas primarios: son los que salen de las cisternas del aparato de Golgi, contienen enzimas digestivas y todavía no han participado en la digestión. Lisosomas secundarios: se forman al unirse con otras vesículas, por lo que han realizado la digestión. Dependiendo del origen del material que han digerido se pueden denominar de dos maneras: vacuolas heterofágicas o digestivas (bacterias) y vacuolas autofágicas (mitocondria). Observa el esquema al final del párrafo: En él se representa una célula que digiere una bacteria del exterior y una mitocondria de la propia célula. Para digerir la bacteria, la célula la rodea con parte de la membrana, formando una vesícula con la bacteria en su interior (fagosoma). Los lisosomas primarios que salen del aparato de Golgi se dirigen a ella, fusionando su membrana con la del fagosoma y vierten sus enzimas digestivas que digieren la bacteria y se forma así un lisosoma secundario (vacuola heterofágica). Como resultado de la digestión se pueden generar residuos no digeridos, que quedan encerrados en unas vesículas denominadas cuerpos residuales y que son expulsados fuera de la célula por exocitosis. Cuando la célula digiere materia orgánica del exterior, los lisosomas pueden tener función nutritiva, porque ayudan a digerir nutrientes complejos que la célula toma del medio, o como en el dibujo, los lisosomas pueden tener función defensiva, porque ayudan a destruir bacterias y virus que son engullidos por células defensivas. Para poder digerir la mitocondria que no funciona, la célula la rodea con membranas del retículo liso formando una vesícula llamada autofagosoma. Al igual que antes, los lisosomas procedentes del aparato de Golgi se dirigen hacia allí y se fusionan con el autofagosoma, se forma así un lisosoma secundario (vacuola autofágica). Sus enzimas digieren la mitocondria y los restos no digeridos quedan en el cuerpo residual y son expulsados fuera de la célula. Vacuolas: En las células eucariotas existen enclaves citoplasmáticos en los que se acumulan diferentes tipos de sustancias. Si estos enclaves están rodeados de membrana se denominan vacuolas, en caso contrario inclusiones. Las vacuolas tienen su origen en la fusión de vesículas procedentes del aparato de Golgi. Están presentes en todo tipo de células, pero son especialmente abundantes en las células vegetales, en las que generalmente ocupan el 50% del volumen celular llegando en algunos casos al 95%. Las vacuolas de las células vegetales acumulan en su interior sustancias hidrosolubles que de no estar confinadas por una membrana se dispersarían por todo el citoplasma. Entre estas sustancias se encuentran productos de deshecho del metabolismo celular, sustancias de reserva, pigmentos e incluso algunos alcaloides venenosos que la planta utiliza para alejar a los depredadores. Las vacuolas también ayudan a regular el equilibrio osmótico de las células vegetales. En las células animales también existen vacuolas; entre ellas destacan las vacuolas pulsátiles que algunos protozoos que viven en medios hipotónicos utilizan para bombear hacia el exterior el exceso de agua.