Regulación enzimática
Anuncio

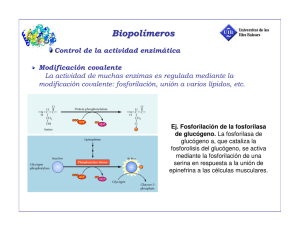
Tema 13 Regulación enzimática Ya sabemos que todos los procesos celulares están regulados, esta regulación necesaria, se basa en la posibilidad de aumentar o disminuir el flujo a través de una vía. El flujo se entiende como la velocidad de conversión del primer metabolito en el último. La regulación de una etapa no es más que modificar la actividad de la enzima que la controla. Mecanismos de regulación • Generales: Se basan en la disponibilidad de sustrato o producto. Por ejemplo, la disponibilidad de glucosa aumenta el flujo de la glucolisis, de igual forma que si eliminamos el piruvato. • Específicos: La regulación de la actividad de una o varias de las enzimas de la vía. Ya que lo que se modifica es la velocidad, veamos las ecuaciones que la rigen. V = K [S]; Vruta = [E] [metabolíto]. A través de ella podemos hacernos una idea de lo que se modifica. Enzimas reguladoras del flujo En una ruta hay dos tipos principales de enzimas, las que denominamos reguladoras del flujo, que son aquellas que al aumentar su cantidad de enzima activa aumentamos el flujo, y aquellas que no son reguladas de forma específica. El grado de influencia sobre el flujo a través de la vía se denomina Coeficiente de control de flujo Esto es así porque no podemos aumentar más el flujo de la vía que la actividad de la enzima que estamos regulando específicamente. Para poner un ejemplo: Una hormona puede aumentar la cantidad de enzima activa de 100 a 500 veces, de modo, que el denominador sería 400/100 = 4. Como mucho, entonces (recordemos que no puede ser mayor que 1), el numerador puede ser 4, pero pongamos que es 2. De esta forma tendríamos un de 2/4 = 0,5. Cuanto mayor coeficiente presente, más importante será la enzima en el control del flujo a través de la vía metabólica. Estas enzimas reguladoras catalizan etapas lentas ya que la cantidad de la cantidad de enzima activa que está presente es menor que el de la s demás de la ruta, por ello, las etapas lentas (limitantes) son irreversibles. Un ejemplo claro de esto es la etapa de la glucolisis catalizada por la PFK, que requiere ATP. La bajada de la concentración del producto debido al drenaje del a vía haría que faltara mucho para conseguir el equilibrio y, por tanto, la reacción es espontanea. En cuanto a la ruta en general, el paso de Glu a Pyr produce ATP con una *Go' de "15,3 Kcal/mol, y el inverso, consumiendo ADP de .4,2 Kcal/mol. Ambas son espontaneas, por lo que se estarían dando a la vez, algo realmente estúpido para la célula. En la realidad, algunas de las enzimas que catalizan etapas irreversibles están reguladas, como por ejemplo la que veíamos antes, la PFK, que requiere otra enzima para la reacción inversa, la F1,6Bpasa, de esta forma, cuando se requiere glucosa aumenta la actividad de la fosfatasa y disminuye la de la kinasa, de forma inversa a lo que ocurre cuando hay demanda de 1 energía. La regulación de la enzima se puede conseguir a dos niveles: • Regulación de la cantidad de proteína enzimática: mediante la síntesis y degradación de la enzima. Este control como es lógico, es a largo"medio plazo. • Regulación de la actividad catalítica: Se trata de un control muy fino y a muy corto plazo. Esta a su ves se subdivide en: • Cambios en la estructura covalente de la enzima de forma reversible, como por ejemplo la fosforilación, metilación o farnesilación. • Cambios conformacionales de la enzima por la unión de moléculas reguladoras. Se trata del modelo típico de regulación alostérica, también es reversible. De este modo, recordamos los distintos tipos de regulación: • General • Específica • Cantidad de proteína activa • Regulación de la actividad • Modificación covalente de la enzima • Unión de ligandos Tipos de enzimas y su importancia en regulación Podríamos preguntarnos si una enzima michaeliana puede llevar a cabo el control a través de una vía metabólica. Para intentar dilucidarlo, no hay más que escoger un valor para [S] pequeño y un grande en relación con Km (de forma equivalente a como lo hacíamos para determinar [S] para Vmax, en función de Km). La diferencia de coger [S] como Km/9 o 9·Km es de 0,1Vmax a 0,9 Vmax. Esto que significa, pues que un aumento de aproximadamente 9 veces requiere añadir sustrato en 81 veces, algo que fisiológicamente no pasa, ya que las concentraciones de los metabolitos son pequeñas y no pueden variar tanto ya que romperían el equilibrio osmótico. De hecho lo que se observa es que pequeñas variaciones en [S] promueven variaciones de hasta 200 veces en la actividad de una enzima. De este modo parece quedar claro que la regulación por [S] pertenece casi exclusivamente a los enzimas alostéricos que, además, también se regulan por efectores. Una de las características más importantes de estos enzimas es la cooperatividad, de la que hablaremos más adelante. Estudio de primeras enzimas de una ruta Monod, Changeux y Jacob estudiaron las características comunes de enzimas que catalizaban la primera reacción de una vía o ruta metabólica inhibida por el producto final de la misma: • El ligando regulador (efector activador o inhibidor) es distinto estructuralmente al sustrato o al producto de la reacción. • Muchas de ellas presentaban cinética sigmoide (algunas enzimas son reguladoras pero con cinética michaeliana), pero hay que tener en cuenta que hay enzimas con cinética sigmoide per no son reguladoras. • Varios tratamientos fisico"químicos (Tº, reducción de puentes disulfuro) desestabiliza la enzima frente al efector, sin pérdida de la actividad catalítica. Esto es debido a la menos posibilidad de transmitir cambios conformacionales en estructuras más flexibles, (similar a la mejor transmisión de los sonidos por sólidos o líquidos que por el aire). • Suelen ser hetero u oligoméricas. 2 La conclusión a sacar es que el efector se une a un sitio distinto del C.A. generando cambios conformacionales en la enzima que causa un cambio en la estructura del C.A., lo que provoca cambios en las propiedades cinéticas de la enzima. De este trabajo surgió en término alosterísmo, no como respuesta a una cinética sigmoide, sino a una regulación por ligando. Cooperatividad Se trata de la propiedad que permite responder con gran sensibilidad a cambios relativamente pequeños en la [S], O2 para nuestro ejemplo, la Hg y Mg. Como hacemos siempre al describir una propiedad cinética, hay que caracterizarla matemáticamente, por ello definimos Y como el grado de saturación, definido por el nº de sitios ocupados por el ligando dividido entre el nº total de sitios de unión. Mioglobina Se trata de una única cadena polipeptídica con cinética hiperbólica y un único sitio de unión por cadena. Mediante el desarrollo de las constantes de la reacción (constante de disociación, de equilibrio, Y) llamando P a la proteína y A al O2 se llega a la conclusión de que Y no puede ser mayor que 1, acercándose sólo en el caso de [A] , cuando todos los sitios están ocupados por el ligando. Hemoglobina Se trata de un tetrámeno (2, 2) con un sitio de unión por subunidad, lo que hace un total de 4. Para esta proteína , que se denomina curva de saturación sigmoide. El exponente h para la hemoglobina es de 2,6. Cooperatividad infinita Imaginamos que inmediatamente y simultaneamente, las subunidades unen ligandos (algo que no ocurre en la Hg). La 1ª molécula que se une aumenta la afinidad de unión del resto por parte de la proteína. El desarrollo de K, nos lleva a una ecuación, similar a la de Hill, pero que ahora presenta n en lugar de h, definiendo n como el nº e sitios de unión dividido entre la molécula (P). es el caso de la Hg, n = 4, que obviamente es distinto de 2,6. Esto es debido a que no hay un equilibrio todo o nada en la unión de ligando. Hay estadíos intermedios (Hg con 1, 2, 3, 4 sitios ocupados). Afirmamos que en la Hg hay cooperatividad positiva, teniendo en cuenta, además, que h<n, nunca igual. Definimos h como una medida de lo fuerte que es la cooperatividad, en mis palabras, la fuerza que hay que ejercer para provovar un estado más receptivo en las subunidad de al lado. Cuando h sea próximo a n, indicará una cooperatividad infinita, que indicará que inmediatamente de la unión del ligando se induce la unión del resto. Para h>1 es positiva, para h=1 no hay cooperatividad (recordad las prácticas) y para h<1 es negativa, lo que significa que la unión de un ligando disminuye la afinidad del resto de las subunidades por el ligando. Determinación del valor de h Al igual que sucedía con la cinética michaeliana, la linearización de la curva de velocidad permite obtener unos valores mucho más reales. Por ello, comenzamos describiendo 1"Y como la fracción de sitios de unión libres, al ser 1 el máximo grado de saturación. A partir de ahí y mediante una serie de desarrollos matemáticos llegamos a una ecuación logarítmica: 3 La representación del primer término Vs log [A] nos da una pendiente igual a n y un corte en ordenadas que nos da el "log de K. A este representación se la denomina representación de Hill. Como ya vimos en prácticas, los datos que obtenemos experimentalmente no es Y, sino la curva de velocidad, por ello, cogiendo los puntos del tramo central, más lineal, representamos la siguiente ecuación: El cambio de n por h se debe a la partida de un supuesto falso, una cooperatividad infinita, al coger el tramo lineal. Definimos ahora K = K0,5 como análoga a Km, es decir, la concentración de sustrato para alcanzar la mitad de la velocidad máxima. Modelo concertado o de simetría Monod, Wyman y Changeux dilucidan este modelo para explicar el alosterísmo. En el propio título se explican varias cosas, concertado implica que las dos subunidades cambian de conformación a la vez y la simetría nos indica que las dos subunidades tienen la misma conformación y la misma constante de disociación. Para enunciarlo partieron de las siguientes premisas: • La proteína es un oligómero. • Existe en dos posibles estados conformacionales T de tenso y R de relajado. La primera predomina cuando el ligando no está unido. • La conformación R tiene más afinidad por el ligando, lo que significa que la constante de disociación de T es mayor que la de R y, por lo tanto, el ligando se une preferentemente a R. • Todos los sitios de unión en cada estado son equivalentes (ya lo habíamos dicho, es la condición de concertado) y tienen idénticas constantes de disociación por los ligandos (resultado de la simetría). En definitiva, todas las subunidades cambian de conformación a la vez Parámetros ♦ ♦ ♦ : El grado de saturación. : Fracción de proteína en estado R ♦ ♦ . En ausencia de ligando L>1. . De modo que, en concordancia con lo que dijimos antes C<1. Origen de la cooperatividad 4 La base es que la proteína predomina en ausencia de ligando en estado T, pero el ligando se une preferentemente a R. Por esto, nos preguntamos: ¿Cómo puede aumentar ? • Aumentando la concentración de ligando • Aumentando el estado R • Las dos Al aumentar el ligando, se satura el estado R, más receptivo, lo que lleva al equilibrio entre las dos formas a desplazarse, aumentando a su vez el nº de sitios activos. De este modo a mayor L (más T y menos R en situación inicial, necesaria para que se produzca un desplazamiento, ya que si toda la proteína se encuentra en un solo estado no hay posibilidad de cooperatividad) y menor C (KT > KR), mayor será el efecto cooperativo (mayor h) Regulación por interacciones alostéricas El efecto alostérico bien sea positivo o negativo, modifica la razón del equilibrio R"T estabilizando uno u otro. ♦ Activador: se une preferentemente a R aumentando su concentración, lo que provoca los efectos mencionados en el apartado anterior. ♦ Inhibidor: se une preferentemente al estado T disminuyendo R y, por tanto, el nº de sitios activos, lo que conlleva la disminución de . El efecto que tienen estos dos tipos de efectores sobre las curvas de saturación alostéricas ya los conocemos por prácticas. Si el efector alostérico se une sólo (no preferentemente) a uno de los dos estados se pierde la cooperatividad, obteniendo una curva de saturación hiperbólica. Efectos alostéricos ♦ Homotrópicos: Son los producidos por interacciones entre moléculas idénticas del sustrato. Se trata de cooperatividad. ♦ Heterotrópicos: Son los producidos por interacciones entre los efectores alostéricos (moléculas distintas ala sustrato) y el sustrato. Se trata de la regulación alostérica en sentido estricto. Enzimas alostéricas Hay dos sistemas, el K y el V, que detallaremos a continuación: Sistemas K Se caracteriza por tener afinidades diferentes tanto por sustrato como por ligando. Su actividad se regula (positiva o negativamente) por cambios en la afinidad por el sustrato. Kcat no cambia, siendo Vmax constante, sólo cambia K0,5, como se observa en la gráficas de las hojas. Sistemas V Se caracteriza porque tanto R como T presentan la misma afinidad por el sustrato, con lo que se da en enzimas alostéricas sin cooperatividad (recordemos las premisas). LA diferencia, por tanto, entre las dos conformaciones es di distinta Kcat. Es esto lo que permite que estos sistemas tenga representación 5 sigmoide. El trabajo del efector consiste en desplazar el equilibrio R"T, de modo que si predomina la forma de menor Kcat, disminuye la velocidad, siendo entonces un inhibidor y viceversa. Modelo secuencial Este modelo más complicado enunciado por Koshland, Némethy y Filmer parte de dos premisas: • En ausencia de ligando, la proteína existe en una única conformación (al contrario que ocurría con el modelo de simetría). • En presencia de ligando, el cambio de conformación es secuencial, subunidad por subunidad. Origen de la cooperatividad En este modelo, son las interacciones entre las subunidades, bien positivas o negativas, las que favorecen o impiden la unión de ligando a la adyacente. De este modo llamamos cooperatividad positiva a la interacciones favorecedoras de la unión de ligando y negativa a las desfavorecedoras. Cooperatividad negativa o anticooperatividad Este comportamiento sólo se puede explicar con el modelo secuencial. No se ajusta la modelo concertado porque A estabiliza el estado de alta afinidad, por lo que no puede aumentar el estado de baja afinidad. Se caracteriza por una curva Vs [A] sin tendencia a la saturación, con lo que no tiene nada que ver ni con la cinética hiperbólica ni con una sigmoide, ya que a medida que se añade más sustrato hay que ir añadiendo más y más para conseguir una subida que antes o en otros modelos se consigue con poca variación de concentración. Se obtienen resultados equivalentes a diferentes sitios de unión de ligando con distintas afinidades y no interactivos. No es lo mismo cooperatividad negativa que regulador alostérico negativo, ya que el primero se produce por la unión de sustrato y la segunda por la de in inhibidor distinto al sustrato. Además, un efector alostérico negativo no produce cooperatividad negativa. Representación de Scatchard Una forma de dilucidar si nos enfrentamos a una cooperatividad es esta representación. Se basa en estudios de unión de ligando a receptor, y suele utilizarse para ver si un mismo ligando se une a dos o más tipos de receptores. Si partimos de una proteína con un solo tipo de unión, podemos hallar la cantidad de ligando unido, a través de filtrado y posterior contabilización (como el centelleo). El ligando unido depende de la cantidad total de proteína y de la fracción de saturación. De ahí se obtiene la ecuación de una hipérbola. Ya que [E] << [A] podemos considerar, de forma equivalente a como lo hacíamos en temas anteriores que la concentración de A libre es la total. Si consideramos ahora una proteína oligomérica con n sitios de unión por molécula, se nos transforma la ecuación anterior incorporando el término n multiplicando el segundo término. A partir de esta ecuación y mediante un procesamiento se consigue linearizarla, representando la fracción de ligando unido frente al total Vs lo unido. De esta forma, podemos calcular (como se ve en la gráfica de las hojas) n y K, la constante de disociación. 6 Una vez conocida esta linearidad, podemos comparar lo que sucede si cogemos una población de dos proteínas con distintas n y K (siendo K1 < K2). En este caso, la ecuación que nos define el ligando unido será la suma de las dos hipérbolas. Si esto lo representamos podremos obtener dos tipos de curvas, una descendente desde el primer momento, que nos indicará que hay dos poblaciones y que también se corresponderá con una única población de una enzima alostérica con cooperatividad negativa, ya que al cambiar K con la añadidura de ligando, la representación parece indicar que se trata de dos poblaciones. Si se obtiene una curva ascendente en primer lugar, indicará una verdadera suma de afinidad conforme se aumenta el ligando unido, o lo que es lo mismo, una población con una cooperatividad positiva. Si se pudiera observar las dos poblaciones de forma independiente, éstas se corresponderían con las dos rectas discontinuas. Evidentemente aquí pasa algo, ya que si recordamos el las prácticas ese término era S0,5 y, además, de la propia representación, es imposible que K tenga unidades de concentración ya que se trata de un término adimensional. Por ello, y porque así lo demostró en el examen, designamos a partir de ahora S0,5 como la concentración de sustrato para conseguir ½ de Vmax. K es la constante de disociación No entra la ecuación de Adair. ENZIMOLOGÍA 98 Regulación enzimática 7