Descarregar PDF
Anuncio

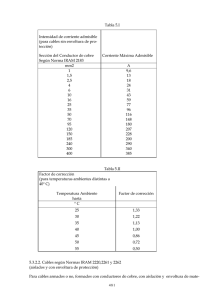
Documento descargado de http://www.elsevier.es el 19/11/2016. Copia para uso personal, se prohíbe la transmisión de este documento por cualquier medio o formato. Actualidad científica Cosmética al día LLORENÇ PONS CONSULTOR FARMACÉUTICO. MIEMBRO EXTERNO DEL COMITÉ EUROPEO DE COSMETOLOGÍA (BRUSELAS). Transglutaminasas (II) Sustratos proteínicos identificados en la envoltura proteínica córnea En la primer parte de este artículo se abordó el papel de las transglutaminasas y su modulación, lo que permite identificar moléculas capaces de compensar su déficit y favorecer su activación, un hecho que favorece la normalización de algunas importantes disfunciones del metabolismo epidérmico, causantes a su vez de enfermedades cutáneas y problemas estéticos. En esta segunda parte se analizan los sustratos proteínicos identificados en la envoltura proteínica córnea, el cornified envelope. n una cuidadosa investigación, Nemes y Steinert1 cuantificaron las numerosas proteínas presentes en la envoltura densa proteínica de los corneocitos. El primer precursor identificado en la envoltura proteínica córnea fue la involucrina. Esta proteína ya se expresa en los estratos epidérmicos suprabasales y contiene 37 secuencias de 10 aminoácidos, cada una de ellas con 3 residuos de glutamina y 2 de glutamato2. Su estructura es compleja, pero predominantemente alfahelicoidal. Aunque su presencia en la envoltura densa no se considera que supere el 5% de las proteínas, conforma un primer «andamio» proteínico a través de enlaces cruzados, catalizados por las transglutaminasas. Otra proteína, denominada loricrina y rica en azufre, se acumula en los gránulos de queratohialina, y su marcada insolubilidad es responsable de que sólo se libere de ellos en el momento de alcanzar la zona de transición. Entonces se desplaza hasta la cara interna del andamio, que se ha formado con la involucrina, y se fija mediante enlaces cruzados catalizados, sobre todo por la TGase 1, situada en este espacio celular periférico. Su presencia en la envoltura densa es muy importante, ya que alcanza el 80% de su contenido proteínico. Se conocen mutaciones de un dominio rico en glicina, que causan algunas enfermedades cutáneas, como la queratodermis3. También hay una familia de pequeñas proteínas ricas en prolina (SPR), que forman 3 subgrupos diferentes (SPR1, SPR2 y SPR3), también conocidas con los nombres de cornifinas o pancornulinas, cuyos residuos de glutamina y lisina forman enlaces isopeptídicos que se incorporan a la envoltura densa gracias a la actividad enzimática de las transglutaminasas. E 128 O F F A R M Su expresión epidérmica depende del calcio y su estructura incluye, en posición central, varias secuencias de proteínas ricas en prolina. En posiciones terminales hay péptidos que tienen, además, residuos de glutamina y de lisina. Estas proteínas, de bajo peso molecular, están presentes en la envoltura densa a una concentración que oscila entre el 3-5%. Se supone que su función es la de actuar de puente para facilitar la unión de otras proteínas4. Otra proteína de bajo peso molecular (unos 12 KDa) conocida con el nombre de cistatina o queratolinina, regula la actividad proteasa que se precisa en el entorno de la envoltura densa, y colabora así en el proceso de maduración de esta trama proteínica. Aunque su presencia es inferior al 5%, un déficit de este tipo de proteínas reduce la función barrera del estrato córneo y causa su deshidratación5. Una proteína de bajo peso molecular (unos 10 KDa), denominada proelafina, se expresa a partir de un precursor tras la eliminación de una secuencia de 25 aminoácidos. Mediante un segundo proceso de hidrólisis, se eliminan dos proteínas que tienen actividad inhibidora de elastasa y proteinasa 3, que se conocen con los nombres de elafina y cementoína. Ambas son buenos sustratos para las trasnglutaminasas, pero su presencia en la envoltura densa es inferior al 1%. La profilagrina es una proteína de muy elevado peso molecular (más de 400 KDa), que mediante hidrólisis li- VOL 24 NÚM 11 DICIEMBRE 2005 Documento descargado de http://www.elsevier.es el 19/11/2016. Copia para uso personal, se prohíbe la transmisión de este documento por cualquier medio o formato. Actualidad científica Cosmética al día bera numerosas unidades de filagrina. Esta filagrina agrega de forma intensa los filamentos intermedios de queratina, para formar así la matriz proteínica intracelular de los corneocitos. Pero su parcial incorporación a la envoltura proteínica refuerza la conexión entre la matriz proteínica y la envoltura densa. Según Presland et al6, la filagrina también puede favorecer, en ocasiones, el desarrollo de procesos apoptóticos en los queratinocitos. Los desmosomas situados en las membranas plasmáticas de los queratinocitos son agregados proteínicos que no sólo conectan con los filamentos intermedios de queratinas, sino que son conexiones puntuales con las células que se hallan en su entorno. Debido a ello, durante la formación de la envoltura densa, algunos componentes de estos desmosomas quedan incorporados a la nueva estructura que sustituye a la membrana plasmática. Según varios autores, entre los que se encuentran Green y Gaudry7, las proteínas desmosómicas implicadas son desmoplacina, envoplacina y periplacina. Además, para Candi et al8 los residuos de lisina en las queratinas de tipo II también son un sustrato de las transglutaminasas y, por tanto, se incorporan a la envoltura densa mediante enlaces isopeptídicos. Por último, entre las proteínas minoritarias de la envoltura densa debe citarse a la denominada anexina 1 (también conocida con el nombre de liportina 1), ya que puede formar un complejo con algunas proteínas dependientes del calcio y colaborar en la apertura de canales de calcio9 durante el proceso terminal de la diferenciación de los queratinocitos, que se desarrolla en un entorno mas ácido y oxidante. tesis de determinadas proteínas precursoras que deben actuar como sus sustratos, en especial cuando falla el nivel de síntesis de alguna de ellas. Otro aspecto que merece reseñarse corresponde a la capacidad de TGase 1 para crear multímeros de loricrina con la utilización de residuos de glutamina y de lisina que pertenecen a diferentes moléculas. Simultáneamente, TGase 3 favorece la formación de enlaces cruzados en el seno de la misma molécula, lo que incrementa de forma muy intensa la resistencia de esta proteína, que sabemos que representa el 80% del material proteínico de la envoltura proteínica córnea. También hay una curiosa especificidad en la formación de puentes isopeptídicos entre las pequeñas proteínas ricas en prolina (cornifinas y pancornulinas), ya que TGase 3 sólo utiliza el dominio A de este tipo de proteínas, mientras que TGase 1 actúa exclusivamente sobre el dominio B. En condiciones normales, la formación de la envoltura densa se realiza con la utilización de los sustratos mediante una secuencia selectiva y ordenada, pero cualquier situación anómala es causante de que las transglutaminasas epidérmicas se adapten a las nuevas circunstancias y solucionen el problema. Por todos estos motivos, el papel de las TGase y su modulación son motivo de diversos estudios que permitirán identificar moléculas capaces de compensar su déficit y favorecer su activación. Es evidente que de esta forma se normalizarán algunas importantes disfunciones del metabolismo epidérmico, causantes de enfermedades cutáneas y de problemas estéticos. ■ Bibliografía Consideraciones finales En esta revisión se pone de manifiesto que la envoltura proteínica córnea posee una estructura muy compleja, en la que no sólo están presentes algunas proteínas específicas (sobre todo, involucrina y loricrina), sino que también las proteínas de la matriz proteínica intracelular de los cornocitos y las proteínas que forman los corneodesmosomas. Además, en el espacio extracelular se sitúa de forma inamovible una empalizada de omega-hidroxiceramidas, que incrementan la hidrofobicidad de los corneocitos y favorecen las uniones hidrofóbicas con los lípidos lamelares. Pero estos datos, en buena parte bien conocidos por los investigadores, se complementan con el decisivo protagonismo que desarrollan en la epidermis las tres transglutaminasas activas, ya que ahora sabemos que su misión no se limita a formar enlaces isopeptídicos, sino que su distribución diferenciada a nivel subcelular regula diferentes aspectos del metabolismo cutáneo. Un aspecto sorprendente reside en el hecho de que algunas de estas TGase son capaces de propiciar la sín- 130 O F F A R M 1. Nemes Z, Steinert PM. Bricks and mortar of the epidermal barrier. Exp Mol Med. 1999;31:5-19. 2. Lambert A, Ekambaram M, Robinson N, Eckert RL. Transglutaminase reactivity of human involucrin. Skin Pharmacol Appl Skin Physiol. 2000;13:17-30. 3. Ishida-Yamamoto A. Loricrin keratoderma: a novel disease entity characterized by nuclear accumulation of mutant loricrin. J Dermatol Sci. 2003;31:3-8. 4. Steinert PM, Candi E, Kartasova T, Marekov L. Small proline-rich proteins are cross-binding proteins in the cornified cell envelopes of strattified squamous epithelia. J Struct Biol. 1998;122:76-85. 5. Zeeuwen PL, Vlijmen-Willems IM, Olthuis D. Evidence that unrestricted legumain activity is involved in disturbed epidermal cornification in cystatin M/E deficient mice. Hum Mol Genet. 2004;13:1069-79. 6. Presland RB, Kuechle MK, Lewis SP. Regulated expression of human filaggrin in keratinocytes results in cytoskeletal disruption, loss of cell-cell adhesion, and cell cycle arrest. Exp Cell Res. 2001;270:199-213. 7. Green KJ, Gaudry CA. Are desmosomes more than tethers for intermediate filaments? Nat Rev Mol Cell Biol. 2000;1:208-16. 8. Candi E, Tarcsa E, DiGiovanna JJ. A highly conserved lysine residue on the head domain of type II keratins is essential for the attachment of keratin intermediate filaments to the cornified cell envelope through isoppeptide crosslinking by transglutaminases. Proc Natl Acad Sci. 1998;95:2067-72. 9. Gerke V, Moss SE. Annexins: from structure to function. Physiol Rev. 2002;82:331-71. VOL 24 NÚM 11 DICIEMBRE 2005