BIOGEOGRAFIA HISTORICA DE LA BIOTA DE LOS DESIERTOS
Anuncio

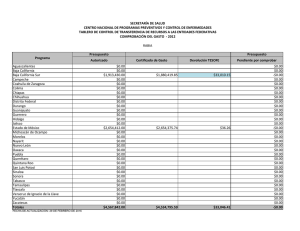
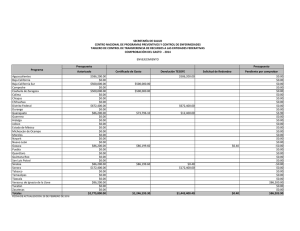
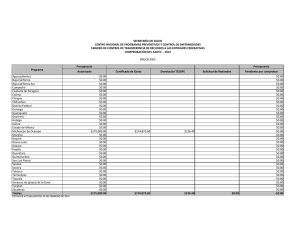
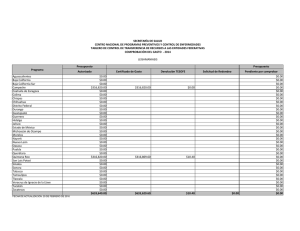
BIOGEOGRAFIA HISTORICA DE LA BIOTA DE LOS DESIERTOS CALIDOS DE NORTEAMERICA Libia Catalina Salinas Castellanos código 2061450 INTRODUCCION Los desiertos de Norte América incluyen el noroeste de México y el suroeste de Estados Unidos. La biogeografía histórica de México es especialmente compleja porque el territorio nacional forma parte de dos regiones biogeográficas: La Neártica y la Neotropical. Refleja los contactos de estas dos grandes regiones con las consiguientes posibilidades de expansión de biotas, así como de evolución en aislamiento durante los periodos geológicos en que no ha habido contacto (Halffter et al. 2008). Hubbard (1973) desarrolló un modelo vicariante del Pleistoceno tardío para explicar la evolución de la fauna de aves de zonas áridas, al imaginar una biota de zonas áridas del suroeste, distribuida ampliamente y de manera continua durante periodos interglaciares, seguida de una fragmentación en dos o tres refugios con poblaciones aisladas, las cuales posteriormente se diferenciaron unas de otras durante los periodos glaciares. Morafka (1977) mencionó el aislamiento y divergencia Pleitocénicos entre los desiertos de Chihuhua y Sonora para la herpetofauna. El objetivo de este trabajo es analizar las relaciones históricas entre las regiones desérticas del noroeste de México y el suroeste de Estados Unidos, con base en los patrones de distribución y la información filogenética de 10 taxa de aves, reptiles y plantas. MATERIALES Y METODOS Las áreas utilizadas en este estudio fueron definidas por Riddle et al. (2006). Como grupo interno se utilizaron 7 regiones ubicadas en el noroeste de México y el suroeste de Estados Unidos distribuidas de la siguiente manera: norte de California (caf), Sur de California (bcaf), desierto de Sonora (dson), desierto de Chihuahua norte (dchin), desierto de Chihuahua sur (dchis), Zonas arbustivas de Sinaloa (zsina) y la meseta sur de México (mex) (Figura 1). Se realizó un análisis de compatibilidad de trazos con una búsqueda heurística de 1000 replicas utilizando el software TNT v1.1 (Goloboff et al., 2008) y Winclada 1.00.08 (Nixon 2002) con lo que se obtuvo la hipótesis primaria de homología. Para establecer el patrón en las relaciones jerárquicas entre las áreas se usó el método transparente o de subárboles libres de paralogía implementado en Nelson 05 utilizando como Libia Catalina Salinas Castellanos Código: 2061450 área externa el oriente de Estados Unidos. Se determinaron las dispersiones y los eventos de vicarianza presentes en el cladograma óptimo de áreas y análisis de ajuste de árboles con el programa Treefitter 1.3B1 (Ronquist, 2002) bajo el modelo de máxima codivergencia (MC), reconciliación y el modelo por defecto (modelo de 4 eventos desarrollado por Ronquist) Se realizó consenso estricto de las topologías resultantes en MC con COMPONENT 2.0 (Page, 1993). Por último se realizó dos análisis de Parsimonia basada en eventos el análisis de dispersiónvicarianza en DIVA 1.2 (Ronquist, 2001). RESULTADOS Y DISCUSION Con el análisis de trazos manual se obtuvieron tres trazos generalizados (figura 1). El trazo uno comprende a las dos áreas de la península de California (caf, bcaf); el trazo dos a los desiertos de Sonora y Chihuahua (dson, dchin y dchis) y el trazo tres al área correspondiente a Sinaloa y la meseta del sur de México; lo cual es congruente con el análisis de compatibilidad de trazos en el que se observa el mismo patrón (figura 2). Riddle en el 2006 explica que estás áreas de semidesierto alcanzaron su máxima extensión durante el principio de Plioceno y disminuyeron de tamaño durante el Plioceno tardío, mucho más húmedo y durante los intervalos pluviales del Pleistoceno. Los desiertos regionales se terminaron de formar durante los periodos interglaciales, y alcanzaron su máxima extensión después del periodo glacial Wisconsiniano. Así el mismo explica que los taxa hermanos presentes en estos desiertos en épocas alternadas podrían compartir ancestría común en una o al menos dos etapas en grandes periodos debido a los aislamientos de especies en la etapa del Mioceno tardío al Plioceno, junto con eventos tectónicos. El cladograma general de áreas resultante del análisis de subárboles libres de paralogía (Figura 3) muestra una relación entre las áreas de la península de california con el desierto de Sonora y el desierto de Chihuahua. Este análisis soporta lo expuesto por Brooks en su artículo del 2001 en el que afirma en cuanto a los grupos de aves que las comunidades en baja California y California contiene siete especies una endémica para cada uno de los seis clados resultado de la vicarianza. Adicionalmente las especies endémicas posiblemente resultaron de especiaciones aisladas periféricas como es el caso de T. redivivum. La comunidad hermana de la península de California es el desierto de Sonora el cual tiene mayor complejidad y riqueza de especies ya que está compuesto por especies endémicas de cada uno de los seis clados producto de eventos vicariantes y dos adicionales resultado de especiaciones aisladas (T. curvirostre y P.fucus). Con el ajuste de árboles basado en parsimonia se obtuvo una topología significativa (p=0) (Figura 4.) bajo el modelo de Ronquist y para el modelo de máxima codivergencia se obtuvieron 5 árboles no significativos con los que se realizó el árbol consenso (Figura 5) que no resuelve las relaciones entre áreas debido a que muestra una politomía entre el norte de California, El desierto de Libia Catalina Salinas Castellanos Código: 2061450 Sinaloa y Chihuahua (norte) y la meseta del sur de México. En el análisis por eventos se observaron dispersiones entre las áreas de la Tabla 2; se encontró un gran número de eventos de dispersión entre las áreas individuales (12 en total), sin embargo el mayor porcentaje de dispersión fue de de 12.5% y se dio en tres de los eventos dispersivos indicando que la dispersión no explica claramente las relaciones entre todas las aéreas de este estudio. De igual manera se observaron 4 eventos de vicarianza dos de los cuales presentaron un porcentaje alto cercano al 43% (Tabla 3). Lo anterior se sustenta por la historia biogeografía de esta zona. Murphy (1983), Grismer (1994), Upton y Murphy (1997) y Murphy y Aguirre-Leon (2002) ampliaron el rango temporal de la historia biogeografica en los desiertos mas orientales para incluir los eventos vicariantes del final del Mioceno y Plioceno propuestos para explicar la formación de la fauna de la península de california. Ciertos eventos específicos incluyen: (1) formación del Mar de Cortés (golfo de california) en el mioceno tardío o Plioceno temprano debido a la actividad orogénica y volcánica a lo largo del borde de las placas de norte América y Pacifico; (2) La inundación en el Plioceno del Istmo de La Paz entre la región de los cabos y el resto de la península; (3) La transgresión hacia el norte de las aguas marinas en la entrada del golfo hacia los desiertos de zonas bajas del norte del lago Mojave y el noroeste de San Gorgonio durante el plioceno o pleistoceno temprano y (4) La creación en el Pleistoceno del paisaje marino del Vizcaino a través de la porción central de la península. Los eventos que dieron lugar a la distribución de la herpetofauna a lo largo de los desiertos del oeste (Sonara y Sinaloa) y del este (Chihuahua) fueron asociados con el surgimiento de la sierra madre occidental y la Meseta Mexicana y a cambios climáticos del pleistoceno y holoceno asociados a la apertura y desaparición de la zona de contacto entre los desiertos del oeste y del este. Adicional a esto Van Devender y colaboradores (1983) han enfatizado repetidamente los eventos Pleistocénicos en el desarrollo de la vegetación de los desiertos de Norte América, particularmente sobre la del desierto de la región sonorense. Por tanto la estructura de los desiertos cálidos de norte América se debe principalmente a eventos de vicarianza debido a que estas áreas comparten una historia en común a pesar de los eventos geológicos que dieron lugar al levantamiento de montañas entre otros y en menor proporción por eventos de dispersión entre áreas. . Libia Catalina Salinas Castellanos Código: 2061450 BIBLIOGRAFIA PUBLICACIONES Brooks, D., & Mclenn, D. 2001. A comparison of a discovery-based and an eventbased method of historical biogeography. Journal of Biogeography 757-767. Halffter, A., Llorente, J., Morrone, J. 2008. La perspective biogeográfica histórica. Capital natural de México, vol.I: Conocimiento actual de la biodiversidad. CONABIO, México, pp.67-86. Leaché, A., Mulcahy. 2007. Phylogeny, divergence times and species limits of spiny lizards (Sceloporus magister species group) in western North American deserts and Baja California. Molecular Ecology 16, 5216–5233 Marshall, C., Liebherr, J. 2000. Cladistic biogeography of the Mexican transition zone. Journal of Biogeography 27:203-216 Morrone, J. 2005. Hacia una síntesis biogeográfica de México. Revista Mexicana de Biodiversidad 76 (2): 207-252 Riddle, B., Hafner, D. 2006. A step-wise approach to integrating phylogeographic and phylogenetic biogeographic perspectives on the history of a core North American warm deserts biota. Journal of Arid Environments 66: 435–461 Riddle, B., Hafner, D. 2006. Biogeografía histórica de los desiertos cálidos de Norteamérica. New Mexico Museum of Natural History and Science Bulletin No.32 Zink, R., Blackwell-Rago, C. 2000. The shifting roles of dispersal and vicariance in biogeography. The Royal society 497-503 SOFTWARE Ducasse, J. Cao, N. Zaragüeta, R. NELSON 0.5, versión 1.0. 2007.Laboratoire Informatique et systématique Université Paris 6- France. (www.phython.org). Goloboff, P.A., Farris, J.S., Nixon, K.C. 2008. T.N.T. Tree analysis using new technology. Programa Y documentación disponible en: http://www.zmuc.dk/public/phylogeny/TNT/. Nixon, K.C. 2002. WINCLADA (BETA), Versión 1.00.08. Published by the author, Cornell University, Ithaca, New York. Nixon, K.C & Carpenter, J.M. (1993) On Outgroups. Cladistics 9;413–426. Libia Catalina Salinas Castellanos Código: 2061450 Ronquist, F. 1996. DIVA version 1.1. Computer program and manual available by anonymous FTP from Uppsala University (ftp.uu.se o ftp.systbiol.uu.se). Ronquist, F. 2002. TREEFITTER, version 1.3b. Computer program and manual available by anonymous FTP from Uppsala University (http://morphobank.ebc.uu.se/TreeFitter). ANEXO 1. 1. Análisis Manual de trazos Figura 1. Análisis manual de trazos. Trazo 1: California (caf) y baja California (bcaf) Trazo 2: Zonas arbustivas de Sinaloa (zsina) y tierras altas de México (mex). Trazo 3: Desierto de Chihuahua norte (dchin), Desierto de Chihuahua sur (dchis) y Desierto de sonora (dson). Libia Catalina Salinas Castellanos Código: 2061450 2. Análisis de compatibilidad de Trazos Figura 2. Análisis de compatibilidad de trazos utilizando TNT . Trazo 1: California (caf) y baja California (bcaf) Trazo 2: Zonas arbustivas de Sinaloa (zsina) y tierras altas de México (mex). Trazo 3: Desierto de Chihuahua norte (dchin), Desierto de Chihuahua sur (dchis) y Desierto de sonora (dson). OUT corresponde al Oriente de Estados Unidos. Libia Catalina Salinas Castellanos Código: 2061450 3. Método basado en el patrón Figura 3. Cladograma general de áreas libre de paralogía obtenido a partir del programa Nelson 05. Libia Catalina Salinas Castellanos Código: 2061450 4. Métodos basado en el modelo Figura 4. Cladograma general de áreas obtenida en el análisis de Ajuste de árboles basado en parsimonia, bajo el modelo de 4 eventos de Ronquist (default). Costo= 24.95 p=0. Libia Catalina Salinas Castellanos Código: 2061450 Figura 5. Cladograma general de áreas obtenido del consenso estricto de 5 árboles generados con el modelo de Máxima codivergencia. Ajuste de árboles basado en parsimonia. Sin embargo ninguna de las topologías usadas para el consenso son significativas. Costo=-36.99. ANEXO 2. Tabla 1. Taxa Utilizados en este estudio y las fuentes bibliográficas de donde se extrajo su filogenia. TAXA Pipilo-Towhees (5) Toxostoma curvirostre (6) Toxostoma lecontei (4) Polioptila (4) Callipepla (4) Vermivora (5) Abronia (23) Coleonyx (7) Mesaspis (6) Viguiera (14) REFERENCIA Zink et al., 2000 Morrone et al., 2005 Los números en paréntesis indican el número de especies presentes en cada grupo Libia Catalina Salinas Castellanos Código: 2061450 Tabla 2. Eventos de dispersión entre áreas individuales obtenidos en el análisis dispersión/ vicarianza. El porcentaje del total de eventos postulados por DIVA se da en la tercera columna. DISPERSIONES ENTRE AREAS INDIVIDUALES De Hasta Frecuencia % A B 1,500 12,5 A C 0,500 4,167 B A 1,500 12,5 B C 0,500 4,167 C A 1,000 8,333 C B 1,000 8,333 C D 1,250 10,417 C E 1,250 10,417 D C 0,250 2,083 D E 1,500 12,5 E C 0,250 2,083 E D 1,500 12,5 Total 12 100 Tabla 3. Eventos de vicarianza entre áreas individuales obtenidos en el análisis dispersión/ vicarianza. El porcentaje del total de eventos postulados por DIVA se da en la tercera columna. FRECUENCIA DE EVENTOS VICARIANTES Evento Frecuencia % AB-C 0,999 42,857 C-DE 0,999 42,857 AB-CDE 0,167 7,164 ABC-DE 0,167 7,164 Total 2,331 100 Libia Catalina Salinas Castellanos Código: 2061450