Tema 6: Apéndices bacterianos. Flagelos, fimbrias y pili.
Anuncio

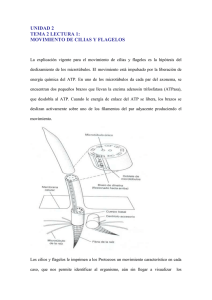
Tema 6: Apéndices bacterianos. Flagelos, fimbrias y pili. Fimbrias y pili Las fimbrias son apéndices cortos, finos, más delgados que los flagelos y muy similares a pelitos que rodean a la bacteria. Se observan principalmente en Gram +. Cada célula puede estar rodeada por unas 1000 fimbrias, pero sólo son visibles al microscopio electrónico por su pequeño tamaño. Se estructuran en base a unos tubos huecos compuestos por fimbrina, organizados helicoidalmente. No tienen función en la movilidad celular, sino que se encargan de fijar las bacterias a superficies sólidas, o a tejidos del huésped. Los pili sexuales son un tipo especial de fimbria. En una célula suele haber 1 – 10 pilis. Sirven para unir la célula a su compañera durante la conjugación bacteriana, y a través de ellas se transmite la información genética de uno a otro individuo. Flagelos Son los dedicados al movimiento bacteriano. La mayor parte de los microorganismos son móviles, por lo que se trata de una ventaja frente a los que no tienen movilidad. · Flagelos perítricos Los flagelos perítricos se distribuyen de forma uniforme por toda la superficie bacteriana. · Flagelos polares Los flagelos presentes se encuentran agrupados en un polo de la bacteria. Pueden ser: · Bacterias monótricas: Solo tienen un flagelo. · Bacterias anfítricas: Tienen 2 flagelos, uno en cada polo. · Bacterias lofótricas: Presentan un grupo de flagelos en un polo, a modo de penacho. Estructura del flagelo bacteriano · Filamento Es la parte más larga, y se extiende desde la superficie celular hasta la punta del flagelo. Es un cilindro hueco constituido por flagelina, y se dispone helicoidalmente. · Gancho Se trata de un segmento curvado y corto que une el filamento al cuerpo basal. Permite el movimiento porque actúa como acoplamiento flexible. Noemie Martínez Biron – Grupo 121 – Microbiología 2010 / 2011 1 · Cuerpo basal Se encuentra anclado a la pared celular. Es el motor del flagelo. Tiene distintos anillos según se trate de un organismo Gram + o Gram -. En la mayoría de Gram - tiene 4 anillos unidos por un vástago central. Los anillos externos están asociados a la capa en la que se insertan, el anillo L con los liposacáridos, y el anillo P con el peptidoglicano. Los anillos internos SM y C se insertan en la membrana plasmática. En Gram + solo aparecen los anillos internos. Tipos de movimiento · En medios sólidos El movimiento puede producirse por la rotación de un enjambre de flagelos, por el movimiento de pilis sexuales, o por deslizamiento sobre una superficie. · En medios líquidos · Movimiento flagelar básico: Los flagelos producen el movimiento al rotar. El filamento tiene forma de hélice rígida, y la bacteria se mueve. El sentido de rotación determina la dirección que toma la bacteria. Las bacterias que se mueven en un medio isotónico suelen parar y girar (cambiando el sentido de rotación del flagelo) al azar. Este movimiento se denomina taxia. Si introducimos un nutriente, la bacteria lo puede detectar y moverse hacia él. Si lo que introducimos es un compuesto tóxico, la bacteria se moverá en dirección contraria. En bacterias monótricas con su flagelo en posición polar, el flagelo gira en un sentido, y puede cambiar de dirección haciéndolo girar en sentido opuesto. En bacterias monótricas con el flagelo apolar, los cambios de sentido no pueden realizarse como los anteriores. La bacteria se para, se deja llevar por el medio, y cuando está orientada hacia donde quiere, vuelve a mover su flagelo. En bacterias perítricas, basta con que uno de los flagelos cambie su sentido de rotación para que la bacteria voltee hacia otra dirección. Taxias · Quimiotaxia Desarrolla una respuesta de la bacteria frente a algún componente químico. E.coli posee 4 quimiorreceptores aceptores de metilos (MCPs): · Tsr: Receptor para la serina. · Tar: Receptor para el aspartato. · Trg: Receptor para la ribosa, la glucosa y la galactosa. Noemie Martínez Biron – Grupo 121 – Microbiología 2010 / 2011 2 · Tap: Receptor para dipéptidos. Las MCPs no actúan directamente sobre la rotación flagelar. Actúan a través de una serie de proteínas citoplasmáticas llamadas Che. Las MCP son proteínas transmembrana, por lo que dan a parar al citoplasma y al periplasma. En el lado periplasmático de las MCP, tiene lugar la unión con moléculas atrayentes. Un atrayente favorecerá la rotación normal, mientras que un repelente favorecerá un volteo. El lado citoplasmático de las MCP (se llama PQM) interactúa con las proteínas Che W (unida a la Che A, una quinasa capaz de autofosforilarse usando ATP). · Cuando le célula camina: El receptor MCP (PQM) se encuentra en un estado conformacional en el que no hay posibilidad de interactuación con Che W. De esta forma, Che W tampoco interactúa con Che A. · Cuando la célula voltea: El receptor MCP (PQM) se encuentra en un estado conformacional ideal para la interacción con Che W, y ésta con Che A. Al tener capacidad de autofosforilación, esta proteína fosforilada cede su fosfato a Che Y, que interacciona (una vez fosforilado) con una proteína flagelar específica (Fli M). Esta proteína se encuentra en la base del flagelo, y es la que induce una rotación, provocando el giro. Cuando la concentración de nutrientes en el medio es grande, Che A se mantiene desfosforilada, lo que provoca que no haya demasiados giros. En cambio, en un medio pobre en nutrientes, Che A se mantiene fosforilada, provocando cambios continuos. En una situación de equilibrio, se mantienen niveles intermedios de fosforilación de Che A y Che Y para favorecer los giros aleatorios. También es importante la forma en la que la proteína MCP (PQM) adquiere una u otra conformación. En este proceso están implicadas las demás Che: · Che R: Es una proteína metiltransferasa, que introduce continuamente grupos metilo en MCP (PQM). Cuando llega a un nivel determinado de metilación, cambia su conformación. · Che B: Produce el efecto contrario a la anterior. Esta proteína se fosforila a través de Che A y desmetila a MCP (PQM), cambiando de nuevo su conformación. · Fototaxia Se da principalmente en bacterias fotosintéticas y halobacterias. Las bacterias detectan una luz determinada, y se dirigen a ella (o al contrario, según esa luz les sea o no beneficiosa). · Aerotaxia Se trata de una respuesta por parte de una bacteria a una presencia o ausencia de oxígeno. El proceso se desencadena por una proteína (Aer) unida a un cofactor FAD que se encarga de detectar los cambios en las concentraciones de oxígeno. Noemie Martínez Biron – Grupo 121 – Microbiología 2010 / 2011 3 Si las concentraciones de oxígeno son bajas, un componente del sistema de transporte de electrones interacciona con FAD. Este modifica a Aer, provocando una cascada de fosforilaciones Che, dando lugar al movimiento celular. · Magnetotaxis Permite a los microorganismos terrestres orientarse por el campo magnético terrestre. Existe en organismos con presencia de magnetosomas, los cuales sí pueden verse afectados por los campos magnéticos. Movimiento axial Producido por giro del propio cuerpo celular como consecuencia del movimiento de flagelos internos situados en el periplasma celular. El único grupo de microorganismos que posee este tipo de movimiento son las espiroquetas. Pueden vivir en medios acuosos o como huéspedes, y producen muchas enfermedades (sífilis, leptospirosis, fiebres recurrentes, Lyme). Las espiroquetas son Gram –. El movimiento axial produce un desplazamiento tipo “sacacorchos”. Cuando el microorganismo quiere cambiar de dirección, los flagelos de uno y otro polo se paran, y vuelven a ponerse en marcha en la misma dirección, cambiando de sentido. Noemie Martínez Biron – Grupo 121 – Microbiología 2010 / 2011 4


![[Libro] "El marxismo y la justicia social. La idea de... en Ernesto Che Guevara"](http://s2.studylib.es/store/data/003536401_1-0ffe0dcd9ebcd44131a00eac1da28c4f-300x300.png)