Formación del embrión tridérmico EMBRIOLOGÍA DEL SISTEMA
Anuncio

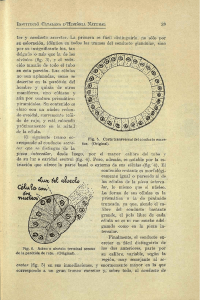
Capítulo 39 583 Embriología e Histología de las Vías Urinarias 6 3 1 1 2 7 Fig. 39.5 2 5 Epiblasto (1), hipoblasto (2), mesoderma extra-embrionario (3). 3 5 4 Fig. 39.4 Cavidad amniótica (1), cavidad vitelínica primitiva (2), sinciciotrofoblasto (3), citotrofoblasto (4), blatocisto con mesoderma extra-embrionario (5), epiblasto (6), hipoblasto (7), y membrana de Heüser (8). Con la aparición del celoma extra-embrionario el mesodermo extra-embrionario queda dividido en dos porciones, una que reviste internamente el citotrofoblasto, y una capa que envuelve externamente las vesículas amniótica y vitelínica primitiva. El crecimiento comparativo del didérmico al de los anexos embrionarios es pequeño, teniendo el disco didérmico en esta fase el tamaño de 0,1 a 0,2 mm (Langman, 1985). Formación del embrión tridérmico La gastrulación es el proceso por el cual el embrión didérmico es convertido en un embrión tridérmico. La gastrulación se inicia con la formación de la línea primitiva en la superficie del epiblasto del disco embrionario. En seguida las células abandonan la superficie profunda de la línea primitiva, formando una malla de tejido embrionario, que, ingresando en el hipoblasto, promueve la aparición del endodermo definitivo y queda entre las dos capas, produciendo el mesoderma intra-embrionario (Fig. 39.5). La línea primitiva, que formóel mesoderma activamente hasta el inicio de la cuarta semana, disminuye gradualmente a su producción, desapareciendo al final de la cuarta semana. El mesoderma, rellena el espacio entre el ectoderma y el endodermo, no formándose en dos puntos, donde estos continúan adheridos: en las membranas bucofaríngea y cloacal. Algunas células migran cefálicamente del nudo primitivo (nudo de Hensen), formando un cordón primitivo llamado notocordio o notocorda, que se localiza entre las células del mesoderma intra-embrionario. Este notocordio, además de definir el eje primitivo del embrión, induce la formación de la placa neural en el ectoderma, que da origen a la corredera neural, en seguida a las crestas neu- Part III (39-47).indd 583 rales y al tubo neural. Las crestas neurales darán origen a los ganglios sensitivos de los nervios craneanos y espinales y a otras varias estructuras y el tubo neural origina el sistema nervioso central, es decir, el encéfalo y la médula. El mesoderma forma, de cada lado del notocordio, condensaciones cilíndricas conocidas como mesoderma paraxial, condensaciones menos pronunciadas, conocidas como mesoderma intermediario y, externamente, las placas del mesodermo lateral. El mesoderma paraxial va a originar los somitos, el mesodermo intermediario va a originar parte de los sistemas urinario y genital, y el mesodermo lateral se dividirá en dos placas: una, en contacto con el ectodermo, formará la somatopleura, y la otra en contacto con el endodermo, irá a originar la esplacnopleura (Fig. 39.6). EMBRIOLOGÍA DEL SISTEMA URINARIO Parte del sistema urogenital tiene origen en el mesoderma intermediario, habiendo una relación entre los desarrollos urinario y genital, de tal forma, que los componentes de uno son usados más tarde en la construcción de otro sistema. Embriología de los riñones y uréter El sistema adquiere su forma antes del desarrollo de las gónadas, surgiendo subsiguientemente tres grupos de estructuras: pronefro, mesonefro y metanefro. Estas estructuras surgen jun- 6 1 3 7 4 2 1 8 2 Fig. 39.6 Ectoderma tegumentario (1); endodermo (2), mesoderma paraxial (3); mesoderma intermediario (4); mesoderma lateral (5); ectodermo neural formando la corredera neural (6); tubo neural (7); notocordio (8). 30/01/2009 05:56:11 p.m. 584 Parte III Uroginecología to con la formación de un conducto que drena la orina producida por la cloaca (conducto de Wolf o conducto mesonéfrico), surgiendo de él un brote ureteral que originará el uréter. Al inicio de la cuarta semana embrionaria, el mesoderma intermediario comienza a fragmentarse craneocaudalmente y los cinco o siete primeros pares de fragmentos van a constituir el pronefro, que en la embriología humana es rudimentario y no funcional, desapareciendo con la aparición de la fragmentación posterior formadora del mesonefro. Los fragmentos del pronefro, conocidos como nefrótomos, se conectan lateralmente con un par de conductos pronéfricos, que crecen en dirección a la cloaca (seno urogenital) y también degeneran, siendo continuadas en la misma dirección por los conductos mesonéfricos o conductos de Wolf. Al final de la cuarta semana los conductos mesonéfricos ya están presos en la cloaca, habiendo surgido, bien cerca de la inserción en la cloaca, en cada conducto de Wolf, un brote ureteral. En el mesonefro se forman cerca de 40 túbulos mesonéfricos en una gradiente cefalocaudal, que evolucionan hacia vesículas mesonéfricas abrazando de un lado un mechón capilar y del otro lado desembocando en el conducto de Wolf, produciendo la orina mesonéfrica. Algunas de esas vesículas mesonéfricas degeneran al final del primer trimestre y otras van a participar de la génesis de constituyentes de los genitales masculinos. Dado que las vesículas mesonéfricas más craneales regresan cuando se forman las caudales, no se encuentran más de 30 vesículas mesonéfricas funcionantes en un momento determinado. Estas vesículas mesonéfricas se diferencian en unidades excretoras, siendo una versión abreviada de la nefrona adulta. En ellas encontramos inclusive una estructura llamada cápsula de Bowman, que envuelve una red de capilares llamada glomérulo renal mesonéfrico, formando un corpúsculo renal mesonéfrico. Estos capilares son originados en la aorta dorsal del embrión. Cada corpúsculo renal, con su tubo mesonéfrico, desembocando en el conducto mesonéfrico de Wolf, forma una unidad excretora mesonéfrica (Fig. 39.7). Las unidades excretoras mesonéfricas están funcionantes entre la sexta y décima semana, produciendo su orina mesonéfrica. Después de la décima semana ellas regresan. En la mujer también regresa totalmente el conducto mesonéfrico. En el hombre persiste el conducto de Wolf originando el epidídimo, el conducto deferente y las vesículas seminales. En el hombre, algunas vesículas mesonéfricas van a originar los conductos eferentes del testículo. El metanefro o riñón permanente inicia su desarrollo al comienzo de la quinta semana y su funcionamiento cerca de cuatro semanas más tarde (Behrmann et al., 1996). La orina continúa siendo arrojada en la cavidad amniótica durante toda la vida fetal. Las células caudales del mesoderma intermediario se condensan a cada lado del brote ureteral para formar el blastema metanéfrico. La base morfológica del desarrollo de un riñón me- Part III (39-47).indd 584 Crispi 1 2 4 3 8 6 7 5 Fig. 39.7 Conducto pronéfrico que regresará (1) ; nefrotomos que regresarán en la embriología humana (2) ; mesonefro que se fragmentará formando aproximadamente 40 unidades (3) ; conducto mesonéfrico o de Wolf (4) ; metanefro (5); seno urogenital (6); brote ureteral (7); alantoides (8). tanéfrico es el alargamiento y la ramificación (hasta 14 ó 15 días) del brote uretral o divertículo metanéfrico, que se transforma en el sistema de ductos colectores (metanéfricos) del metanefro, y la formación de las vesículas metanéfricas renales a partir de condensaciones mesénquimales (blastema metanéfrico) situado a cada lado de las extremidades de las ramas. El mecanismo subyacente a estos eventos está constituido por una serie de interacciones inductoras recíprocas entre las extremidades de las ramas de los ductos metanéfricos y las células del blastema metanéfrico circundante. Sin el sistema de ductos metanéfricos las vesículas metanéfricas no se forman. De modo inverso, el mesoderma metanéfrico actúa sobre el sistema de ductos metanéfricos, induciendo su ramificación característica (Carlson MB, 1996). El divertículo metanéfrico o brote ureteral es el primordio del uréter, pelvis renal, cálices y túbulos colectores y su pedículo se vuelve el uréter, y su extremidad cefálica expandida forma la pelvis renal. Las cuatro primeras generaciones de túbulos aumentan de tamaño y se vuelven confluentes, formando los grandes cálices, y las cuatro generaciones siguientes coalescen formando los pequeños cálices. Las generaciones siguientes forman los túbulos colectores, con sus diversos colectores, teniendo en la extremidad en la porción correspondiente que generará las vesículas mesonéfricas que se alargan gradualmente hasta transformarse en un nefrón, conteniendo el corpúsculo renal (gloméru- 30/01/2009 05:56:11 p.m. Capítulo 39 585 Embriología e Histología de las Vías Urinarias lo y cápsula de Bowman), el túbulo contorneado proximal, el asa de Henle y el túbulo contorneado distal. Al final, este nefrón se abre hacia el túbulo colector lanzando su orina metanéfrica. El blastema metanéfrico se fragmentará tantas veces como veces se ramifiquen los ductos metanéfricos (Fig. 39.8). El aumento de tamaño del riñón después del nacimiento resulta principalmente del alargamiento de los túbulos contorneados proximales y del aumento del tejido intersticial. La formación de los nefrones nacidos a término está completa al nacimiento. La maduración funcional de los riñones ocurre después del nacimiento. La filtración glomerular comienza alrededor de la novena semana fetal, y la velocidad de filtración aumenta después del nacimiento (Arant, 1987). Durante la fase final del periodo embrionario y el inicio del periodo fetal los riñones se desplazan superiormente entrando 2 en contacto con las glándulas adrenales, que forman un capuchón de tejido glandular en el polo cefálico de cada riñón. Aunque normalmente irrigado por una gran arteria renal que sale directamente de la aorta, el riñón adulto consiste en cinco lóbulos vasculares. Las arterias que alimentan cada uno de esos lóbulos eran originalmente vasos segmentarios que suplían el mesonefro y que fueron asimilados por el metanefro en migración. Son de orígenes aórticos, son típicamente reducidos a un único par de arterias renales, pero son comunes las variaciones anatómicas (Carlson, 1996). Embriología de la vejiga y uretra El estudio de la embriología de la vejiga comienza por la septación de la cloaca a través del septo urorectal, formando posteriormente el conducto anorectal y anteriormente el seno urogenital, que se continuará superiormente por el alantoides (Fig. 39.9). El alantoides se originó en la porción superior y posterior de la vesícula vitelínica. La membrana cloacal, que es uno de los puntos donde el mesoderma no se forma entre el ectodermo y el endodermo, se divide en membrana urorectal y membrana anal. El seno urogenital está dividido en tres partes 1. Una porción superior, la vejiga urinaria, que se continúa por el alantoides y posee una base expandida continua con el seno urogenital y un proceso tubular delgado, que se extiende por un pedículo del embrión en la otra extremidad. Inicialmente la vejiga es continua con el alantoides, pero cuando la luz del alantoides es obliterada, un cordón fibroso 1 3 4 5 2 Fig. 39.8 Fig. 39.9 (A y B) Fragmentación del blastema mesonéfrico (1); túbulos colectores (2); otra generación de tubo colector (3); vesícula metanéfrica en formación (4). Conducto mesonéfrico o de Wolf (1); brote uretral (2); alantoides (3); somitos originados del mesoderma paraxial (4); seno urogenital (5) Part III (39-47).indd 585 30/01/2009 05:56:12 p.m.