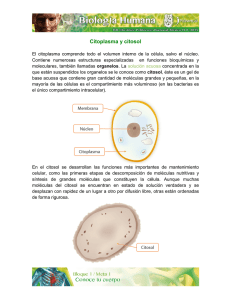
Organización general de las células: Citosol y sistema de
Anuncio

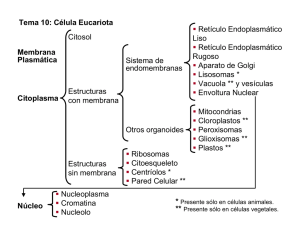
Organización general de las células: Citosol y sistema de endomembranas 1 Tema 15. Peroxisomas. Vacuola vegetal. Almacenamiento intracelular de sustancias o paraplasmas. Glúcidos intracelulares. Acúmulos de naturaleza proteíca. Depósitos de lípidos. Pigmentos y sustancias minerales. Resumen general del metabolismo celular citosólico. 1. Peroxisomas Los peroxisomas, junto con las mitocondrias, son los orgánulos celulares que desempeñan un papel primordial en la utilización del O2. 1.1 Estructura Morfológicamente son partículas esféricas, limitadas por membrana, de diámetro variable 0,2-1,5 µm y con un contenido enzimático, rico en enzimas que intervienen en el metabolismo del H2O2, colaborando con mitocondrias y cloroplastos en algunas funciones. Se han observado en casi todos los tipos celulares animales y en algunos tipos celulares vegetales de plantas superiores que realizan la fotorrespiración y en el endosperma de semillas (glioxisomas). Con microscopía electrónica presentan un contenido granular fino y pueden ser identificados por técnicas citoquímicas específicas (Diaminobenzidina y H2O2) que reaccionan con la intervención de una de las enzimas del peroxisoma, la catalasa. En el interior de algunos peroxisomas pueden observarse estructuras cristalinas que generalmente corresponden a la enzima urato oxidasa. La estructura de estos nucleoides paracristalinos varía según el tipo celular; en general, los nucleoides aparecen formados por estructuras en forma de túbulos, formando distintos tipos de redes. 1.2. Función 1.2.1. Actividad enzimática Los peroxisomas se denominan así porque contienen enzimas que utilizan el oxígeno molecular para eliminar átomos de hidrógeno de substratos específicos, a través de una reacción oxidativa que produce H2O2. La reacción global sería: RH2 + O2 ⇒ R + H2O2 Siendo RH2 el substrato oxidable (aminoácidos, cetoácidos α , ácido úrico, alantoína, acil-CoA, ácido glicólico, ácido glioxílico, enoil-CoA, etc.). Las enzimas que catalizan está reacción son: la oxidasa de aminoácidos, oxidasa de hidroxiácidos α y oxidasa de urato. Organización general de las células: Citosol y sistema de endomembranas 2 El H2O2 resultado de la reacción es un producto altamente tóxico que es eliminado por otra enzima del peroxisoma, la catalasa, bien directamente según la reacción: 2H2O2 ⇒ 2H2O + O2 o utilizando este H2O2 para oxidar diversas sustancias (alcoholes, fenoles, ácido fórmico, formaldehído y acetaldehído), según la reacción: 2H2O2 + R'H2 ⇒ 2H2O + R' Siendo R'H2 una de las diversas sustancias mencionadas anteriormente. No siempre las cuatro enzimas (catalasa, oxidasa de aminoácidos, oxidasa de hidroxiácidos α y oxidasa de urato) aparecen en todos los peroxisomas, pero al menos la catalasa y oxidasa de hidroxiácidos α están siempre presentes. Mediante su dotación enzimática, variable de unos tejidos a otros, los peroxisomas pueden intervenir en diversas vías metabólicas, que difieren según los organismos y tipo de tejido. 1.2.2. Catabolismo de las purinas En la degradación de las bases púricas (adenina y guanina) intervienen diversas enzimas del peroxisoma. El H2O2 que se libera con estas oxidaciones es descompuesto por la catalasa. El producto final de la degradación es: - Acido úrico: Primates, aves e insectos. - Alantoína: Mamíferos, tortugas y moluscos. - Acido alantoíco: Algunos peces teleósteos. - Acido glioxílico y urea: Muchos peces, anfibios e invertebrados marinos. 1.2.3. Metabolismo de los lípidos Los peroxisomas intervienen en el metabolismo de los lípidos mediante los siguientes procesos (aunque no todos ellos ocurren en todos los organismos): - a) β-oxidación de los ácidos grasos Aproximadamente un 25% de los ácidos grasos se degradan en peroxisomas y el resto en mitocondrias. En ambos orgánulos este proceso de degradación se denomina β-oxidación y conduce a la formación de acetil-CoA. La diferencia reside en que, mientras en las mitocondrias la primera reacción oxidativa es catalizada por una deshidrogenasa, en los peroxisomas esta oxidación la realiza una oxidasa flavínica. La oxidación del acil-CoA por el O2 forma H2O2, que es descompuesto por la catalasa. Los acetilCoA formados pasarán a diferentes rutas (biosíntesis de azúcares, ciclo de Krebs). - b) Conversión de grasa en carbohidratos: Ciclo del glioxilato El acetil-CoA producido en la degradación de ácidos grasos puede seguir una ruta metabólica importante en algunos órganos vegetales (endosperma de semilla de ricino, Euglena sp.). Las moléculas de acetil-CoA producidas en la degradación de los ácidos grasos en el peroxisoma (glioxisoma) se utilizan para Organización general de las células: Citosol y sistema de endomembranas 3 producir ácido succínico, en el proceso conocido como ciclo del glioxilato. El ácido succínico producido en este ciclo abandona los peroxisomas y penetra en las mitocondrias, en cuya matriz es oxidado por las enzimas del ciclo de Krebs a ácido oxalacético, que abandona las mitocondrias y se convierte en glucosa en el citosol. 1.2.4. Metabolismo del ácido glicólico En órganos de plantas en los que se realiza la fotorrespiración, en presencia de abundante luz, O2 y baja tensión parcial de CO2 se produce ácido glicólico, que es un subproducto metabólico de los cloroplastos producido por fijación de O2 en la ribulosa-1,5-diP. El ácido glicólico entra en los peroxisomas y es oxidado a ácido glioxílico, que se convierte en glicina, que pasa de los peroxisomas a las mitocondrias donde se transforma en serina y CO2. 1.2.5. Otras funciones Las reacciones oxidativas de los peroxisomas son muy importantes en hígado y riñón, donde detoxifican gran cantidad de moléculas tóxicas que entran en la circulación sanguínea (p. ej. el etanol). 1.3. Origen de los peroxisomas Junto con las mitocondrias, los peroxisomas son los principales lugares de utilización del O2. Una hipótesis es que los peroxisomas son un vestigio de un orgánulo antiguo, que en los antecesores primitivos de las células eucarióticas realizaba todo el metabolismo del O2. Cuando el O2 producido por las bacterias fotosintéticas empezó a acumularse en la atmósfera podría haber resultado altamente tóxico para la mayoría de las células. Los peroxisomas pudieron haber contribuido a disminuir la concentración de O2 en estas células, a la vez que permitían utilizar su reactividad química para realizar reacciones oxidativas útiles. De acuerdo con ésto el desarrollo posterior de las mitocondrias hizo que los peroxisomas se volvieran altamente obsoletos; dado que muchas reacciones que ellos realizaban sin producir energía fueron acopladas a la formación de ATP por medio de la fosforilación oxidativa. Por tanto, las reacciones oxidativas que realizan los peroxisomas actualmente podrían ser las que siguen siendo útiles a pesar de la presencia de las mitocondrias. Se pensaba que los peroxisomas podían originarse por gemación del retículo endoplasmático rugoso desprovisto de ribosomas y que almacenarían enzimas peroxisómicas, sintetizadas por ribosomas libres. Estas proteínas están provistas de un péptido señal carboxilo terminal, de tres aminoácidos, que es reconocido por receptores específicos de la membrana del peroxisoma. Estos receptores se sintetizarían por el retículo endoplasmático rugoso, al originar la membrana del peroxisoma, quedarían alojados en el lado citosólico de dicha membrana. Actualmente, se piensa que los peroxisomas son capaces de reproducirse como las mitocondrias y cloroplastos. En este caso los componentes de membrana serían importados del citoplasma. Tienen una vida media de aproximadamente 4 días y medio, y se destruyen por autofagia. 2. Vacuola vegetal Organización general de las células: Citosol y sistema de endomembranas 4 En las células vegetales jóvenes (meristemáticas) existen numerosas vesículas y vacuolas de pequeño tamaño (provacuolas), que sólo se aprecian bien con el microscopio electrónico. Al crecer estas células, las provacuolas adquieren mayor tamaño y se fusionan hasta formar una gran vacuola central, que en algunas células, como las parenquimáticas, llega a ocupar más del 90 % del volumen celular. La vacuola se halla limitada por una membrana citoplasmática, que se denomina tonoplasto, con estructura trilaminar, de menor espesor que la membrana plasmática; es decir, como las membranas citoplasmáticas, aunque en algunos casos se ha descrito que su espesor es igual al de la membrana plasmática. Mediante criofractura se observa una imagen similar a la de la membrana plasmática: en la hemimembrana externa se aprecian numerosas partículas pequeñas que hacen protrusión, mientras que en la interna hay menos partículas pero de mayor tamaño. El examen de cortes al microscopio electrónico muestra que la hemimembrana interna es más gruesa que la externa, al igual que ocurre en la membrana plasmática. En la vacuola hay un jugo vacuolar de apariencia amorfa incluso con microscopía electrónica; pero a veces también pueden observarse estructuras de mayor tamaño, cristalinas o no, según los productos que almacene la vacuola, y que guardan relación con su función. 2.1. Funciones La vacuola lleva a cabo funciones fisiológicas importantes, que podemos resumir en las siguientes: 2.1.1. Facilitar el intercambio con el medio externo Como el intercambio de sustancias del interior de la célula con el medio externo sólo puede realizarse a través de la membrana plasmática, cuando las células animales desean aumentar dicho intercambio, cambian de forma aumentando la relación superficie/volumen, como ocurre, p. ej., en las microvellosidades. En las células vegetales, la gruesa pared celular no permite esa modificación, y el problema se resuelve gracias a la vacuola. Si ésta no existiera, la célula tendría todo el citoplasma y orgánulos ocupando el mismo volumen que ahora tiene, pero con una superficie de dimensiones mucho más reducidas. Al aparecer la vacuola, el volumen del citoplasma no aumenta, pero se extiende ocupando una fina capa entre la pared celular y la vacuola. De esta manera, la célula vegetal se hace grande y desarrolla una gran superficie de membrana plasmática en relación con el pequeño volumen que ocupa el citoplasma, si no incluimos en este volumen el que ocupa la vacuola. Esa misma disposición aumenta la eficacia de los cloroplastos, evitando en gran medida que se hagan sombra unos a otros. 2.1.2. Turgencia celular La vacuola se encuentra a una gran presión osmótica debido a la alta concentración de azúcares y sales que contiene. Su membrana presenta particularidades especiales de permeabilidad y transporte. Esto hace que la célula se mantenga turgente, pues el agua tiende a penetrar en la vacuola para equilibrar la presión osmótica. El citoplasma queda así apretado contra la pared celular. Organización general de las células: Citosol y sistema de endomembranas 5 La presión de turgencia de una vacuola puede sufrir cambios controlados como respuesta a fluctuaciones ambientales. La presión se incrementa al aumentar la concentración de solutos, bien en el hialoplasma (mediante la importación de solutos desde el espacio extracelular a través de la membrana plasmática), o bien directamente en la propia vacuola (mediante la síntesis de estos solutos por hidrólisis de sustancias almacenadas). Cuando estos mecanismos actúan en sentido inverso causan la disminución de la presión. El control de la presión de turgencia está regulado por receptores de la membrana plasmática que responden a los cambios de presión induciendo un bombeo de K+ hacia el citoplasma (para contrarrestar la disminución de la presión) o la difusión de K+ hacia el exterior a favor de gradiente (para contrarrestar el aumento de presión). La presión de turgencia varía ampliamente de unas plantas a otras: desde 0,5 hasta 50 atmósferas. En plantas de hábitat salino se requiere una alta concentración de solutos para aumentar la turgencia. Un + mecanismo rápido para aumentar la presión sería la incorporación de K ; sin embargo, altas concentraciones de este ion no serían viables y, por eso, estas plantas acumulan solutos orgánicos en las vacuolas que pueden alcanzar concentraciones de hasta 0.5 M sin dañar el metabolismo. Entre estos solutos están los polifosfatos, compuestos polihidroxílicos como el glicerol y el manitol, aminoácidos como la prolina, o derivados N-metilados de aminoácidos como la glicinbetana. La vacuola regula también las concentraciones de determinados iones y el pH del hialoplasma, que + alcanzan valores diferentes a los encontrados en la vacuola. La concentración de Na dentro de la vacuola + es entre 4 y 5 veces mayor que en el hialoplasma, debido a una bomba de Na presente en el tonoplasto. + Cuando desciende el pH del medio, la vacuola bombea iones H hacia su interior para ejercer un efecto tampón. Los cambios en la turgencia pueden generar alteraciones en la forma celular si la pared es plástica, como ocurre en los estomas. Durante el día los estomas están abiertos y la luz activa bombas de K+ de la membrana plasmática para mantener alta la presión de turgencia. Por la noche se invierte el proceso y se cierran los estomas. Cambios similares son los responsables de movimientos de la hoja de Mimosa pudica, del cierre de las trampas en las hojas de ciertas plantas carnívoras y de los movimientos de partes florales en la polinización de algunas plantas. 2.1.3. Digestión celular En el interior de la vacuola hay enzimas lisosómicas. De este modo, la vacuola se comporta como fagosoma y citolisosoma. 2.1.4. Acumulación de sustancias de reserva y subproductos del metabolismo En la vacuola hay agua y determinadas sustancias que pueden estar disueltas, floculentes o cristalinas. Entre estas sustancias están: Organización general de las células: Citosol y sistema de endomembranas - Aniones y cationes, como: Cl-, SO2-4 PO3-4, Na+, K+, Ca++ y Mg++. - Hidratos de carbono: • 6 Monosacáridos: Los más abundantes son las hexosas, como la glucosa (en uvas), fructosa (en melocotones), galactosa, manosa y sorbosa. • Disacáridos: Como sacarosa y maltosa (que se forma por el desdoblamiento de polisacáridos, especialmente del almidón). • Polisacáridos: No cuentan ni el almidón (que está en los plastidios), ni la celulosa (que está en la pared celular). El más abundante es la inulina, que por hidrólisis da fructosa. El glucógeno sólo es frecuente en hifas de hongos y algunos líquenes. - Aminoácidos: Son eslabones en la elaboración de materias proteicas que también se almacenan en las vacuolas. El más difundido es la asparagina. En menor cantidad existen leucina, tirosina y ácido glutámico. - Polipéptidos y proteínas: Incluyen: • Enzimas: Como la amilasa (que degrada el almidón en dextrina y luego en maltosa), y diversas lipasas y proteasas. Muchos vegetales contienen en las vacuolas proteínas que inhiben las enzimas que digieren proteínas en animales herbívoros (como el factor inhihidor 1), constituyendo una defensa contra su voracidad. Son muy abundantes en las hojas de solanáceas. Pueden observarse con el microscopio electrónico en cortes como partículas floculentas. • Granos de aleurona: Son reservas proteicas que llenan las vacuolas y son visibles incluso al microscopio óptico. Forman granos regulares (cristaloides proteicos) o irregulares. En el ricino, sobre el cristaloide hay gránulos irregulares que forman el globoide. Se encuentran en gran proporción en el endosperma de semillas y pueden estar en las vacuolas de estas células durante años hasta que, al germinar la semilla, se hidrolizan las proteínas sirviendo de alimento al embrión. - Alcaloides y glucósidos: Se consideran productos de degradación de la actividad metabólica del citoplasma celular y se almacenan en la vacuola. - Pigmentos antociánicos: De naturaleza glucosídica. Dan color rojo a corolas, hojas, algunos órganos caulinares y subterráneos. - Pigmentos flavónicos: Parecidos a los anteriores. Se encuentran en pétalos y dan color amarillo combinados con xantofila. - Taninos: Dan precipitados azules o negros en presencia de sales férricas. Con dicromato potásico dan precipitados pardos. Se encuentran unidos a materias proteicas a las que vuelven insolubles e imputrescibles; de ahí su utilidad para teñir pieles. Se encuentran en el parénquima, al que dan un color pardo. Derivan del ácido gálico o ellágico en combinación con glucósidos. Se consideran un producto de excreción de la planta. - Ácidos orgánicos y sus sales: Presentes en el parénquima de la hoja, fruto y rizomas. Los más frecuentes son los ácidos cítrico, málico, tartárico y oxálico. Son muy abundantes en las plantas suculentas. El ácido oxálico forma cristales en presencia de Ca++ (oxalato cálcico). Si cristaliza con una molécula de agua, los cristales son monoclínicos. Se observan como agujas muy finas. Se encuentran en bulbos de liliáceas y en las hojas de Aloe sp., y se denominan rafidios. Pero si cristalizan con tres moléculas de agua, los cristales son del sistema rómbico. Frecuentemente estos cristales forman bipirámides sobre cuyas caras se depositan pirámides. El conjunto forma una drusa o macla, frecuente Organización general de las células: Citosol y sistema de endomembranas 7 en todas las partes de la planta. Algunos de estos compuestos tienen una función claramente metabólica. Las plantas suculentas abren sus estomas por la noche, cuando la concentración de CO2 ambiental es menor que durante el día, y el CO2 es incorporado en ácido málico que se almacena en las vacuolas. Durante el día, cuando hay energía luminosa, el ácido málico es transformado en azúcares mientras se mantienen los estomas cerrados. 3. Almacenamiento intracelular de sustancias, paraplasmas o inclusiones intracelulares Por lo general, el citoplasma contiene depósitos temporales transitorios constituidos por una reserva de nutrientes o moléculas de otra naturaleza. Las inclusiones intracitoplasmáticas se consideran parte del citoplasma, pero no son orgánulos o componentes principales de la célula sino productos de su actividad metabólica que han quedado dentro de ella. 3.1. Glúcidos intracelulares Los carbohidratos o glúcidos se almacenan como glucógeno, que es un polímero ramificado de la glucosa. Este es catabolizado, según las necesidades para producir glucosa. La degradación enzimática de la glucosa proporciona, a su vez, energía y cadenas cortas de carbono, que vuelven a emplear en la síntesis de diferentes componentes del citoplasma. Con microscopía electrónica, el glucógeno aparece en forma de gránulos o partículas isodiamétricas de 15-30 nm muy densos y homogéneos, a menudo de perfil ligeramente irregular. Cada gránulo de glucógeno es una sola molécula, consistente en una cadena muy ramificada, rodeada por las enzimas que participan en su formación y degradación. A los gránulos aislados se les denomina glucógeno β. Cuando forman acúmulos a modo de roseta, se le llama glucógeno α. La glucogenogénesis y la glucogenolisis ocurren en el citosol, pero parece que guardan una relación topográfica con el retículo endoplasmático liso, muy abundante en la zona donde ocurren ambos procesos. 3.2. Acúmulos de naturaleza proteíca Se observan como inclusiones de estructura cristalina. Generalmente, se encuentran en el citosol; pero a veces también se pueden observar en el núcleo, mitocondrias, complejo de Golgi, retículo endoplasmático rugoso y gránulos de secreción. Casi nunca se conoce su significado funcional, aunque en algunos casos, como los cristales de Reinke de las células de Leydig, se consideran un subproducto metabólico, que se incrementa con la edad. 3.3. Depósitos de lípidos Con frecuencia, la célula almacena lípidos en forma de gotitas esféricas, siendo típicas de adipocitos. Las inclusiones citoplasmáticas formadas por lípidos son, en su mayoría, lípidos neutros Organización general de las células: Citosol y sistema de endomembranas 8 (triglicéridos de ácidos grasos) con un grado de insaturación suficiente para ser líquidos a temperatura corporal. Constituyen una fuente de energía y cadenas cortas de carbono, al ser degradados en glicerol y ácidos grasos; que pueden ser utilizados en la síntesis de componentes de membrana u otras estructuras ricas en lípidos. Los ácidos grasos penetran en la matriz mitocondrial o en los peroxisomas en forma de moléculas de acil-CoA, donde son degradados hasta acetil-CoA en el proceso conocido como β-oxidación. El acetil-CoA producido es oxidado en el ciclo de Krebs. Las inclusiones lipídicas no están rodeadas por membrana. A veces, en la interfase lípido-citosol se observa una capa de finas fibrillas que contienen enzimas del metabolismo lipídico. Esta estructura es muy patente en los adipocitos, donde hay filamentos de vimentina rodeando la grasa. 3.4. Pigmentos y sustancias minerales Los pigmentos dan un color natural al tejido, sin necesidad de tinciones. Para que se pueda considerar pigmento esa sustancia ha de poseer color durante la vida de la célula. Los pigmentos pueden clasificarse en: a) Pigmentos exógenos: Originados fuera del organismo y que entran en él: - Lipocromos-carotenoídes: Abundantes en vegetales (para ellos son endógenos). Los organismos animales los ingieren de los vegetales y quedan disueltos en sus grasas. Así, dan color amarillo al huevo, a la manteca e incluso a la grasa humana. Algunas formas de caroteno son provitaminas que pueden convertirse en vitamina A. - Minerales: Polvo, tintas y elementos como plata, sílice y carbón que, p. ej., se encuentran formando parte de los tatuajes. b) Pigmentos endógenos: - Hemoglobina: Se encuentra en el interior de los eritrocitos y, cuando éstos se destruyen, dan lugar a: • Hemosiderina: Es el pigmento que contiene el hierro. Es de color pardo-dorado. Se encuentra en el citoplasma de macrófagos y en condiciones patológicas puede aparecer en otras células. • Hematoidina y bilirrubina: Es la parte de la hemoglobina que no contiene hierro. La pequeña diferencia entre hematoidina y bilirrubina es que, en la última, el anillo tetrapirrólico está roto. Ambos pigmentos muestran color amarillo pardusco y se oxidan fácilmente a biliverdina, de color verde. - Melanina: De color pardo o negro. Se encuentra en la piel y anexos, y en el ojo. Se produce en los melanocitos. Una variedad de melanina es la feomelanina, de color anaranjado. - Lipofuscina: Contiene material graso, por lo que se tiñe con los colorantes para grasas. Muestran color pardo. Abundan en neuronas, hepatocitos y miocitos cardíacos. Aumentan con la edad. En realidad son el contenido de los cuerpos residuales. - Otros tipos celulares pigmentados (cromatóforos), que están presentes en algunos vertebrados y contienen pigmentos relacionados con carotenoides, son los xantóforos (amarillos) y eritróforos (rojos). Estas células, junto con los melanóforos, dan coloración a la piel de esos animales. Organización general de las células: Citosol y sistema de endomembranas 9 4. Resumen general del metabolismo celular citosólico El citosol es el medio en el cual están sumergidos los diversos orgánulos citoplasmáticos, los cuales extraen de él las sustancias necesarias para su funcionamiento y le vierten algunos de sus desechos. El citosol y el nucleoplasma son las partes de la célula donde se efectúa la casi totalidad de las reacciones bioquímicas. La pauta metabólica de las células eucarióticas está considerablemente afectada por la existencia de compartimentos. La glucolisis, vía de las pentosas fosfato y síntesis de ácidos grasos tienen lugar en el citosol, mientras que la oxidación de los ácidos grasos, ciclo del ácido cítrico, formación de cuerpos cetónicos y fosforilación oxidativa se realizan en la mitocondria. Algunos procesos, como la gluconeogénesis y síntesis de urea, dependen de un juego de reacciones que transcurren en ambos compartimentos. Si el citosol es el medio celular de donde parten y llegan las principales vías metabólicas, es también el lugar donde se producen las transformaciones de energía química en energía mecánica, transformaciones generadoras de diversos movimientos celulares: movimientos de ciclosis, ameboides, ciliares y de contracción. Finalmente, por medio de algunos de sus constituyentes, el citosol contribuye a la forma celular.