TEMA 7: Proliferación celular.
Anuncio

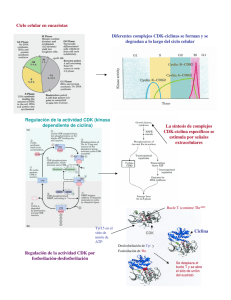
L. Zedón Muñoz TEMA 7: Proliferación celular. Enero de 2007 78 Ciclo celular y regulación de la multiplicación celular. - Fases del ciclo celular. Componentes del sistema de control del ciclo celular. Sistema de control del ciclo celular. Puntos de restricción del ciclo celular. Proteínas quinasa reguladoras del ciclo celular: Ciclinas y Quinas dependites de ciclinas (Cdk). El sistema de control del ciclo celular depende de la proteolisis cíclica. El sistema de control del ciclo celular depende de la regulación transcripcional. Inhibición del crecimiento celular. Señales negativas de regulación de los puntos de restricción. Proteínas reguladoras del ciclo celular. Control extracelulular de la división celular: Mitógenos del ciclo celular. Factores de crecimiento. Factores de supervivencia. Matriz extracelular e interacciones célula-matriz. La matriz extracelular inuye en la forma celular, la supervivencia y la proliferación. PROLIFERACIÓN CELULAR De acuerdo con la teoría celular establecida por el biólogo alemán Rudolf Vircho en el siglo XIX, las células sólo provienen de células. La vida de una célula se inicia con su formación a través de la división de una célula madre y termina con la formación de sus células hijas o con su muerte. Las etapas a través de las cuales pasa la célula desde una división celular a la siguiente constituyen el ciclo de la célula. Sin embargo, este ciclo no lo realizan todas las células: Células lábiles : son células que se dividen y mueren continuamente. Ej: epidermis, epitelios de mucosa, tracto gastrointestinal. Células estables : suelen estar en reposo G0, teniendo una escasa capacidad de división. Sin embargo, pueden entrar en el ciclo celular tras una agresión. Ej: hepatocitos, epitelio tubular renal, acinos pancreáticos. Células permanentes : son aquellas que pierden su capacidad de división al nacer el individuo, presentando una diferenciación terminal. Ej: neuronas, cardiomiocitos, músculo esquelético. La función básica del ciclo celular es la de duplicar en forma exacta la gran cantidad de DNA cromosómico y luego distribuir las copias en células hijas genéticamente iguales. La duración del ciclo celular varía de manera signicativa en los distintos tipos celulares. Ej. células epiteliales del intestino (12h), células hepáticas humanas (1 año). (5) Fases del ciclo celular. El ciclo celular se divide en dos fases principalmente: 1. Interfase, que consta de: Fase de síntesis (S) : En esta etapa la célula duplica su material genético para pasarle una copia completa del genoma a cada una de sus células hijas. Fase G1 y G2 (intervalo) : Entre la fase S y M de cada ciclo hay dos fases denominadas intervalo en las cuales la célula está muy activa metabolicamente, lo cual le permite incrementar su tamaño (aumentando el número de proteínas y organelos), de lo contrario las células se harían más pequeñas con cada división. 79 2. Fase M - Mitosis (M): En esta fase se reparte a las células hijas el material genético duplicado, a través de la segregación de los cromosomas. La fase M, para su estudio se divide en: Profase Metafase Anafase Telofase Citocinesis Cuando ya no se requieren más células, estas entran en un estado denominado G0, en el cual abandonan el ciclo celular y entran en un periodo de latencia. Esto no signica que entren en reposo, ya que éstas células siguen presentando un metabolismo activo. Ahora bien, si estas células reciben el estímulo adecuado abandonan el estado G0 y entran al G1. Algunas poblaciones celulares altamente especializadas como las bras musculares o neuronas al entrar en estado G0 abandonan indenidamente el ciclo celular. COMPONENTES DEL SISTEMA DE CONTROL DEL CICLO CELULAR Puntos de control del ciclo celular. En el ciclo celular hay tres puntos controlados por sistemas de vigilancia del núcleo celular, diseñados para evitar que las células se puedan replicar si están dañadas, sobre todo a nivel del ADN. Una célula detenida en el ciclo por estos mecanismos puede: activar los mecanismos de reparación del ADN o, si fallan, empezar el proceso de apoptosis para eliminarse. 1. Punto de control G1 : Revisa que las composiciones del medio sean favorables para la proliferación de la célula (temperatura adecuada, presencia de nutrientes, sales), que esta haya crecido lo suciente y que el material genético esté intacto. El punto de control G1 es el más importante ya que coincide con el punto de Restricción del ciclo celular. 2. Punto de control G2-M : garantiza que las células no ingresen en la fase de Mitosis hasta que no se haya reparado el ADN y se complete su replicación. 3. Punto de control M : se encuentra en la fase de mitosis, entre la metafase y la anafase. Se encarga de revisar que todos los cromosomas se hayan unido al huso mitótico. Sustancias que controlan el ciclo celular. CICLINAS: la entrada y la progresión de las células en el ciclo celular está controlado por cambios en las concentraciones y actividades de una familia de proteínas denominadas ciclinas. Las concentraciones de las distintas ciclinas ascienden en fases especícas del ciclo celular y después se degradan con gran rapidez a medida que la célula atraviesa en ciclo. Estas proteínas no ejercen actividad enzimática por sí mismas, pero se han de unir a las cinanas (quinasas) para que estas últimas se activen. Hay al menos 6 tipos de ciclinas distintas en mamíferos: A, B, C, D, E, F CDK o QUINASA. Las cdk junto con ciclinas forman complejos, siendo los mayores controladores del ciclo celular. En los seres superiores se identicaron dos principales: Cdc Cdk (quinasa dependiente de ciclina) FPM o Factor Promotor de la Maduración : El FPM está formado por dos subunidades: cdk y ciclinas. 80 Para que la célula abandone la fase G1 e ingrese a la fase S, es decir, inicie la replicación del ADN, la ciclina G1 aumenta su concentración a partir del punto R y activa la quinasa cdk2. A partir de este momento ambas moléculas proteicas conforman un FACTOR PROMOTOR DE LA REPLICACIÓN (FPR) que activa la síntesis del ADN. Cuando la concentración de ciclina decrece la cdk2 se libera y el complejo FPR se desactiva. Los niveles de cdk2 son constantes todo el ciclo. Superada la fase G2, se activa el inicio de la mitosis. Al nal de la G2 aumenta la concentración de ciclina mitótica y al alcanzar una determinada concentración se une a la cdc2 componiendo el FACTOR PROMOTOR DE LA MITOSIS (FPM) que se encarga de fosforilar proteínas con funciones esenciales durante la mitosis. Cuando todos los cinetocoros se han ligado a las bras del huso se desactiva este complejo. Punto de Restricción del ciclo celular El punto de restricción se encuentra casi al nal de G1, se conoce así puesto que si la célula lo pasa, se encuentra comprometida irreversiblemente a entrar en ciclo celular, independientemente de lo que suceda en el exterior. Ello ocurre a través de este proceso: Las ciclinas D y E aumentan su nivel y se combinan con quinasas dependientes de ciclinas (enzimas fosforilantes). Las quinasas activas transeren fosfatos del ATP a la proteína pRB (el "freno" del ciclo celular). Cuando la pRB se fosforila libera factores de transcripción que actúan sobre los genes. Estos genes estimulados producirán las proteinas necesarias para que avance el ciclo celular. El sistema de control del ciclo celular depende de la proteólisis cíclica. La fosforilación y la desfosforilación representan uno de los métodos utilizados con mayor frecuencia por las células para estimular o inhibir la actividad de una proteína, y el sistema de control del ciclo celular utiliza este recurso de forma repetida. Las reacciones de fosforilación que controlan el ciclo celular son llevadas a cabo por un conjunto especíco de proteincinasas (Cdk), enzimas que transeren un grupo fosfato a una célula diana. Estas Cdk, se encuentran presentes durante todo el ciclo celular en las células en proliferación. Sin embargo, estas proteínas sólo se activan en momentos determinados del ciclo, cuando se unen a las ciclinas, pasando con rapidez después al estado de desactivación. Por lo tanto la actividad de estas cinasas uctúa cíclica en cuanto a su activación y desactivación. La activación de las Cdk va a depender de la variación cíclica de concentración de las quinasas, las cuales están presentes en todo el ciclo, pero a mayor o menor concentración: Cuando la concentración de las quinasas en el ciclo celular es alta, las Cdk se unen a ellas activándose y formando el complejo ciclina-Cdk. Cuando la concentración de las quinasas en el ciclo celular es baja, las quinasas y Cdk están separadas, sin poderse unir, estando estas últimas inactivas. INHIBICIÓN DEL CRECIMIENTO CELULAR Además de la síntesis y degradación de las ciclinas, en la regulación de los complejos ciclina-CDK también interviene su unión a los inhibidores de CDK. Estos adquieren una importancia especial en la regulación de los cambios de etapa del ciclo celular (G1->S Y G2->M), momentos en los que la célula 81 percibe si ya existe una duplicación suciente de ADN y si se han reparado todos los errores antes de proseguir. Si la delidad de la duplicación del ADN no se controla adecuadamente, se producirá una acumulación de mutaciones y será posible una transformación maligna. Por ejemplo, cuando hay una lesión del ADN (p.ej., por radiación ultravioleta), la proteína supresora de tumores TP53 (antes p53) se estabiliza e induce la trascripción de CDKN1A (antes p21), un inhibidor de la CDK. Este inhibidor detiene las células en G1 o G2 hasta que se produce la reparación del ADN; cuando ello ocurre, la concentraciones de TP53 descienden, así como las de CDKN1A, y la célula continúa el proceso. Cuando una lesión del ADN es demasiado extensa, TP53 inicia una cascada de acontecimientos destinados a convencer a la célula para que se suicide (apoptosis). Proteínas inhibidoras del ciclo celular p53 (proteína supresora de tumores): Es una proteína que funciona bloqueando el ciclo celular si el ADN está dañado, codicada por un gen supresor tumoral. Si el daño es severo, esta proteína puede provocar la apoptosis o muerte celular programada. A la p53 se la conoce como el guardián del genoma. Los niveles de p53 están aumentados en células lesionadas como por ejemplo por radiaciones ionizantes, con lo que se aumenta el tiempo para reparar el ADN por bloqueo del ciclo celular. Las mutaciones de la p53 son las más frecuentes encontradas en el cáncer. Las mutaciones de la p53 heredables producen el síndrome de Li Fraumeni que conduce a una alta frecuencia de cáncer en los individuos afectados. p27 : Es una proteína que se une a ciclinas y cdk bloqueando la entrada en fase S y está bajo el control de la "proteína supresora de tumores (p53). Se ha demostrado que niveles bajos de p27 predicen un mal pronóstico para las pacientes con cáncer de mama. p21 : puede actuar inhibiendo la duplicación en células que ya se encuentran en fase S. Rb (Proteína del retinoblastoma) y Factor de transcripción E2F: la pRb forma un complejo con miembros de la familia del factor de transcripción E2F. Cuando la pRb es fosforilada por las CDKs se libera el E2F, que induce el paso de G1 a S al activar la maquinaria de síntesis del ADN. p15 y p16 : ambas están bajo el control de la p53 y bloquean la actividad del complejo CDK-ciclina D impidiendo que el ciclo progrese de G1 a S. CONTROL EXTRACELULAR DE LA DIVISIÓN CELULAR Los organismos unicelulares, como las bacterias y las levaduras, tienen que crecer y dividirse con la mayor rapidez posible. En cambio, las células de un organismo pluricelular deben ser controladas de manera que se dividan sólo y cuando el organismo necesite otras células, ya sea para permitir el crecimiento de los tejidos o reponer las células perdidas. En consecuencia, para que una célula animal se divida, no es necesario únicamente nutrientes, sino también señales químicas provenientes de otras células, generalmente de células vecinas.(5) La mayoría de las moléculas de señalización extracelular son proteínas solubles secretadas por otras células o proteínas unidas a la supercie de otras células o a la matriz extracelular. La mayoría de ellas actúan positivamente para estimular determinados procesos, pero otras inhiben el proceso. (5) Estas proteínas se dividen en tres grupos dependiendo de su función: Mitógenos Factores de crecimiento Factores de supervivencia 82 Mitógenos Los mitógenos son proteínas que estimulan la división celular, contrarrestando los mecanismos intracelulares de freno (Rb) que bloquean la progresión del ciclo. Ejemplos de mitógenos: Somatomedina Eritropoyetina (EPO) Factores de crecimiento (FGF, PDGF, EGF) Estas proteínas, actúan en la fase G1 para permitir la entrada de la célula a la fase S. Los mitógenos actúan uniéndose a receptores de la membrana con actividad tiroxina-quinasa, los cuales activan la proteína G ?Ras? cambiándola de su estado unido a GDP por GTP (activación). Esta activación desencadena una cascada de fosforilaciones a través de las proteínas MAPK (quinasas activadas por mitógenos), las cuales transmiten el estímulo a diversas moléculas efectoras de trascripción. Esta cascada de fosforilaciones ocasiona la trascripción de genes tempranos (entre los que destacan los que codican a las ciclinas de G1) y algunos de estos genes a su vez activan la trascripción de otros genes denominados genes tardíos. De esta manera, la vía de señalización Ras-MAPK trasmite señales extracelulares al núcleo activando la maquinaria del ciclo celular. Factores de crecimiento. El crecimiento de un organismo o de un órgano depende tanto del crecimiento como de las divisiones celulares. Si las células se dividiesen sin crecer, se tornarían cada vez más pequeñas. Los factores de crecimiento estimulan el crecimiento celular (aumento de la masa celular) mediante la promoción de la síntesis y la inhibición de proteínas y otras macromoléculas. El crecimiento de las células no depende del ciclo celular. Por ejemplo, las células nerviosas y musculares, crecen sobretodo después de la división celular. Al igual que la mayoría de los mitógenos los factores de crecimiento se unen a receptores de supercie celular que luego activan diversas vías de señalización intracelular, las cuales podrán inducir: un aumento de la síntesis proteica o bien una disminución de la degradación proteica. Todo ello conducirá al crecimiento celular. Factores de crecimiento: Factor de crecimiento epitelial (EGF): estimula la división al unirse a un receptor de tiroxina quinasa de la membrana celular. Causa el crecimiento de los dientes. Factor de crecimiento transformante-alfa (TGF-alfa): es homólogo al anterior y se une a su mismo receptor. Factor de crecimiento derivado de plaquetas (PDGF): ocasiona que durante el desorden embrionario se produzca crecimiento desproporcionado del encéfalo en comparación del crecimiento del resto del cuerpo. Estimula la migración y proliferación de broblastos, células musculares lisas y monocitos, pero también tiene otras propiedades proinamatorias. Factor de crecimiento broblástico (FGFs): se unen a la heparina y a otras moléculas aniónicas (y, por tanto, presentan una intensa anidad por la MB). Poseen otras actividades, además de estimular el crecimiento celular. Factor de crecimiento transformante- beta (TGT-beta) Citoquinas: IL-1, TNF...: estos factores no solo inhiben a las células epiteliales, sino que dependen del tipo de célula en la que incidan, pudiendo favorecer el crecimiento, como en las células mesenquimatósas. En concentraciones bajas induce la síntesis de PDGF, siendo mitógeno indirecto. 83 Factores de supervivencia. Promueven la supervivencia celular por supresión de los programas de suicidio intracelular o apoptosis. Esta necesidad de señales de otras células para la supervivencia, contribuye a que las células sólo sobrevivan cuando y donde se las requiera. Por ejemplo, las neuronas son producidas en exceso y entre ellas compiten en la obtención de factores de crecimiento, siendo las que más tengan las que sobrevivirán. Al igual que los mitógenos y los factores de crecimiento, los factores de supervivencia suelen unirse a receptores de la membrana celular, lo cual activa vías de señalización intracelular que inhiben la muerte celular, en general mediante la regulación de proteínas pertenecientes a la familia Bcl-2. Algunos factores de supervivencia son: IL-3 SCF IGF-1: Es secretado principalmente por el hígado en respuesta a señales de la hormona del crecimiento (GH). MATRIZ EXTRACELULAR E INTERACCIÓN CELULA-MATRIZ La Matriz extracelular (MEC) es un complejo macromolecular, dinámico y en constante remodelación que se sintetiza a nivel local y que constituye una proporción importante de cualquier tejido. Las macromoléculas que constituyen la matriz extracelular son de cuatro grandes tipos: Sistema colágeno. Sistema elástico. Proteoglicanos. Glicoproteínas multifuncionales (laminina, bronectina, tenascina, trombospondina y otras). Cada una desempeña funciones de manera integrada con las demás; esto hace que la matriz sea calicada como un verdadero complejo funcional. Los colágenos y el sistema elástico constituyen la arquitectura de la matriz extracelular. Las glicoproteínas actúan como moléculas de adhesión del sustrato intercelular, importantes en las interacciones célula-célula y célula-matriz. Los glicosaminoglicanos y proteoglicanos tienen un papel fundamental en el equilibrio hidroelectrolítico y ácido básico. Con todo ello la MEC proporciona turgencia a los tejidos blandos, rigidez al hueso, sustrato para la adherencia celular y es imprescindible para la regulación del crecimiento, el movimiento y la diferenciación de las células que residen en ella. Existen dos formas básicas de MEC. - Matriz intersticial: se encuentra en los espacios que separan las células del tejido conjuntivo y entre el epitelio y las estructuras vasculares y musculares lisas de sostén; sintetizada por las células mesenquimatósicas (p.ej., broblastos). Sus componentes principales son los colágenos brilares y no brilares, proteoglucanos y proteínas. - Membrana basal: se encuentra bajo los epitelios y su síntesis depende tanto de estos como de las células mesenquimatósicas subyacentes. Sus principales componentes son el colágeno tipo IV y glucoproteinas no adhesivas. Su función es la de separar el tejido epitelial del conjuntivo y forma parte del aparato ltrador del riñón. En el crecimiento y la diferenciación celular intervienen al menos dos tipos de señales que actúan en concierto: Una procede de moléculas solubles como los factores de crecimiento polipeptídicos y los inhibidores del crecimiento. 84 En la otra participan componentes insolubles de la MEC que interactúan con las integrinas celulares. La interacción entre la MEC y los factores de crecimiento pueden inuir en el crecimiento, la movilidad, la diferenciación y la síntesis proteica celular. Las integrinas se unen a la MEC e interactúan con el citoesqueleto en complejos de adherencia focal. Esto puede iniciar la síntesis de segundos mensajeros intracelulares o mediar directamente las señales nucleares. Los receptores de los factores de crecimiento situados en la supercie celular también pueden iniciar las segundas señales. Todos son integrados por la célula para generar distintas respuestas, como variaciones del crecimiento, la locomoción o la diferenciación celular. La perturbación de este delicado equilibrio de interacciones célula-matriz puede provocar pérdida de la arquitectura tisular con varias consecuencias para el tejido, por ejemplo pérdida en los patrones de diferenciación, proliferación y muerte celular. Un ejemplo de esta perturbación es la que presentan las glándulas salivales de pacientes con síndrome de Sjögren, donde se observan cambios en la estructura, organización y función de acinos y ductos y de la MEC. Por otro lado, la interacción de los leucocitos con la MEC es esencial para la migración, localización en tejidos, desarrollo y función de estas células que residen en ella. EN RESUMEN: El ciclo celular es un conjunto de procesos ordenados, que lleva a cabo la célula cuando se le ha instruido el dividirse; está dividido en interfase y mitosis El control del ciclo celular se presenta a dos niveles, intracelular y extracelular. El control intracelular está a cargo de mediadores proteicos que ejercen un control negativo y positivo cobre el ciclo celular (cdk-ciclinas y CKI). Existe un punto de restricción y tres puntos de control, los cuales son supervisados por distintas combinaciones de Cdks-ciclinas. La entrada al ciclo celular no es una decisión que la célula toma individualmente; se requiere de las señales adecuadas (mitógenos) ya sea del medio extracelular o de otras células. Cuando una célula no es necesaria o es una posible amenaza esta puede morir por apoptosis ya sea por señales intracelulares o extracelulares. Bibliography [1] http://www.facmed.unam.mx/deptos/embrio/Archivos/Docencia/Ciclocel.pdf [2] Karp G. (1998). Biología celular y molecular, Editorial McGraw?-Hill Interamericana Editores S.A., México 580-588. [3] Vinay Kumar MD. Ramzi Cotran S. Stanley Robbins L. (2004). Patología humana, Editorial Elsevier España S.A., Génova, España 61-70, 179-185. [4] Guzmán Ortuño Pacheco. Francisco J. Martínez Díaz, Francisco A. Pastor Quirante, Pedro D. Blanco Muñoz (2005). Lecciones de Anatomía Patológica General para estudiantes de Medicina y Odontología, Editorial DM, Merced, Murcia 148. [5] Alberts B. Bray D. Hopkin K. Johnson A. Lewis J. Ra M. Roberts K. Walter P. (2004). Introducción a la Biología Celular, Editorial Médica Panamericana, Madrid España 613-633. [6] Stevens A. Lowe J. (2001). Anatomía Patológica, Editorial Harcourt, Madrid 93. [7] http://enciclopedia.us.es/index.php/Ciclo_celular [8] http://es.wikipedia.org/wiki/IGF-1 [9] http://ciruelo.uninorte.edu.co/pdf/salud_uninorte/16/2.Matriz_extracelular.pdf [10] http://icbm.cl/prog_biolo_celular/laboratorios/lab1.html [11] http://www.biologia.edu.ar/cel_euca/regulacion.htm