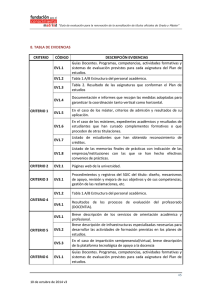
10. Composición de medios de cultivo utilizados Todos los
Anuncio

10. Composición de medios de cultivo utilizados Todos los medios de cultivo y soluciones stock se esterilizan a 120 ºC durante 20 minutos Medio YMA modificado Medio original de Vincent, 1975; Ferrera Cerrato et al., 1993. (ELMARC) K2HPO4 0,655 g MgSO4.7H2O 0,2 g NaCl 0,1 g Manitol 2,5 g Sacarosa 7,5 g Extracto de levadura 0,5 g Agar 15 g Soluc. Rojo congo * 1 ml pH 6.5 - 6.8 Agua destilada 1000 ml * Rojo congo 1g Agua destilada 400 ml En los cultivos líquidos se emplea el mismo medio sin la solución de rojo congo. 44 Medio glucosa peptona agar (GPA) Ferrera Cerrato et al., 1993 Glucosa 10 g Peptona de carne 5g Púrpura de Bromocresol* 10 ml Agar 15 g pH 7.0 * Púrpura de Bromocresol 0.1% p/v Agua destilada 1000 ml Medio Triptona – Extracto de levadura (TY) Bacto-Tryptona 5g Extracto de levadura 3g CaCl2. 2H2O 0,87g Agar 15g pH 6.7 Agua destilada Medio M9 (Beringer, 1974) 1000 ml (Miller, 1972) Na.2HPO4 5,8 g KH2PO4 3,0 g NaCl 0,5 g *NH4Cl 1,0 g Agar 15 g pH 6.7- 6.8 Agua destilada 1000 ml * Se añade solo para probar fuentes carbonadas 45 Soluciones que se preparan por separado y esterilizadas a 120ºC por 20 minutos Compuesto MgSO4 1mM Biotina 0,5µg/ml **Sacarosa 0,2% Stock 6,05mg/25ml Volumen para 100ml 50,15µl/100ml 10mg/25ml 125µl/100ml 10g/150ml 1ml/100ml ** Se añade solo para probar fuentes nitrogenadas Medio YM modificado para fuentes carbonadas (Marquina, 2006) K2HPO4 0,655g MgSO4.7H2O 0,2g NaCl 0,1g NH4NO3 * 0,1g Azul de bromotimol - Agar - Manitol ** 9g ***Fuente carbonada 0,1% pH Agua destilada 6.7- 6.8 1000 ml * Se incorpora para probar fuentes carbonadas ** Se incorpora para probar fuentes nitrogenadas *** Se incorpora al medio esterilizado por filtración 46 Solución Hoagland modificada (Fuente: Eipsten 1972 citado por Taiz y Zeiger, 2002) Macronutrientes Concentración de sol. Stock Volumen de sol. stock (g/L) (ml/L) KNO3 101,10 6 Ca(NO3)2 236,10 4 NH4H2PO4 115,8 2 MgSO4.7H2O 246,49 1 CaCl2.2H2O* 58,9 10 KH2PO4* 54,4 5 KCl* 7,864 2 * Se agregan solo para sustituir las fuentes de Nitrógeno Micronutrientes CuSO4.5H2O 0,062 2 ZnSO4.7H2O 0,288 2 NaMoO4.2H2O 0,107 1 MnCl.2H2O 0,396 1 H3BO3 0,773 2 FeSO4.7H2O 0,557 1 Na2EDTA.2H2O 0,745 1 pH 6,5 - 6,8 Los macronutrientes se agregan al momento de enrazar, para evitar su precipitación en el medio. 47 RESULTADOS Y DISCUSIÓN 1. Aislamiento y purificación de cepas rizobiales a partir de suelos tratados con y sin carbamatos 1.1 Captura de rizobios a partir del suelo empleando como cultivo trampa plantas de Phaseolus vulgars L. y Macroptilium lathyroides L. En el ensayo de captura de rizobios a partir del suelo cultivado con Solanum tuberosum L. (papa), se observó al cabo de 30 días, la formación de 10 nódulos en total de coloración blanca en 2 de las 5 plantas de P. vulgaris L. indicando una distribución heterogénea de la población rizobial en las muestras de suelo recolectadas (Anexo 4), situación similar que se observó en Macroptilium lathyroides L., leguminosa utilizada también como cultivo trampa. Estas leguminosas son huéspedes promiscuos de amplio espectro para las bacterias rizobiales (Bromfield y Barran, 1990; Michiels et al., 1998). En el caso de P. vulgaris, se conoce que puede ser nodulada por varios géneros rizobiales, aunque no siempre son nódulos efectivos en cuanto a la fijación y puede mostrar un grado de preferencia por ciertos rizobios (Pacovsky et al. 1984; Aguilar et al. 2001). Los resultados fueron poco alentadores en la captura efectuada en el suelo cultivado con Allium sativum L. ya que se formó una sola estructura similar a un nódulo de 1 mm de diámetro (que pudo haber sido un nódulo incipiente, el cual no concretó su formación) en una sola planta en ambos cultivos trampa. En el suelo con vegetación natural (no cultivado) fueron negativos los resultados, es decir no se formaron nódulos en los cultivos trampa, por lo tanto este suelo fue descartado eligiendo posteriormente como control los rizobios aislados de nódulos de plantas de Vicia faba L. (Anexo 4) cultivada en la parcela cercana que no había sido tratada con carbamato. El hecho de no formarse nódulos en el ensayo de captura del suelo con vegetación natural y no cultivado, significa que la población rizobial es muy baja debido probablemente a la ausencia de leguminosas como plantas huésped. Además la densidad poblacional rizobial puede variar enormemente en 48 cuestión de metros, que para una bacteria es una distancia muy grande y está sujeta a cambios por ejemplo de pH, humedad, exposición al sol, contenido de materia orgánica etc, los cuales pueden determinar un ambiente muy diverso para los rizobios en una distancia muy corta (Lodeiro et al., 2003). Con la referencia de estos resultados se hizo un análisis de suelos a las parcelas cultivadas con S. tuberosum L. y A. sativum L. (Tabla 12), en el Laboratorio de Suelos de la Escuela de Geografía de la Facultad de Ciencias Forestales y Ambientales de la Universidad de Los Andes, para verificar si la diferencia de las respuestas fue debida a las diferencias en las características fisicoquímicas de los suelos. Tabla 12. Análisis de suelos de las parcelas cultivadas con Solanum tuberosum L. y Allium sativum L. Granulometría Nombre Suelo (papa) Suelo (ajo) pH agua 1:2 C.O. M.O. Nitrógeno Calcio (meq/ 100g) Relación C/N arena (%) arcill a limo Clase textural 72 12 16 Fa 5.63 1.78 3.06 0.13 2,53 13.64 62 16 22 Fa 5.11 1.52 2.63 0.13 1,85 11.64 (%) Los resultados mostrados en la Tabla 12 no reflejan una diferencia en las concentraciones de nitrógeno en ambos suelos que pueda explicar la inhibición de la simbiosis en las leguminosas- trampa sembradas en suelos cultivados con A. sativum L. (ajo), por posibles excesos de fertilizantes nitrogenados aplicados en esa parcela. Sin embargo, en cuanto al pH determinado por la relación 1:2 proporción suelo : agua en este laboratorio, y según la interpretación de Malagón (1982) los suelos de pH ácido están en el rango de 5.0 - 5.5 y correspondería al suelo donde fue cultivado A. sativum L. (ajo), y los suelos de pH ligeramente ácido se encuentran entre el rango de 5.5 – 6.5 en donde estaría ubicado el suelo donde fue cultivado con S. tuberosum L. (papa)., podría proporcionar la diferencia entre la población rizobial 49 de ambos suelos, pero esto dependiendo de las especies de rizobios que se encuentren en esa zona, debido a que la tolerancia de los rizobios a la acidez es muy variada y depende de la especie y aún de la cepa. En general se considera que los rizobios de crecimiento rápido son menos tolerantes a la acidez que los de crecimiento lento (Graham et al., 1994) y la infección y la nodulación se reduce en suelos ácidos, variando las respuestas dependiendo de las especies leguminosas y los rizobios (Gil, Martínez, 1994). De hecho Evans et al. (1980) encontraron que en el caso de Pisum sativum, la nodulación resultó 10 veces más susceptible a la acidez que la propia multiplicación de los rizobios o el crecimiento de la planta. Además el bajo pH puede conducir a los rizobios a la muerte, detener su metabolismo, o generar cambios mucho más moderados dependiendo del nivel de acidez, por ejemplo en R. leguminosarum se ha descrito la pérdida de movilidad pero no de crecimiento a pH 5.0 (Glenn y Dilworth, 1994). En el laboratorio se determinó la tolerancia a pH ácidos de los aislados ES1 y EV1, y éstos crecieron en el rango de 6.0 a 7.0, lo que permite inferir que las bacterias obtenidas no son resistentes a la acidez y que las que se capturaron probablemente eran provenientes de una población que se encontraba en un microambiente donde las condiciones del suelo tales como pH, no eran tan ácidos, que le permitió infectar al huésped. La diferente nodulación en ambos suelos también podría deberse a la concentración de calcio (Ca), que según Malagón (1982) serían suelos muy pobres en este elemento (0 – 5%), ya que este elemento juega un papel importante en diversos procesos bacterianos como quimiotaxis, virulencia, transporte de azúcares y proteínas, reacciones de fosforilación, estabilización de los lipopolisacáridos (LPS) de la membrana externa y la activación entre enzimas entre otros, y en el caso de los rizobios es conocido que el calcio afecta su crecimiento, supervivencia, adsorción a los pelos radicales del hospedador, y su capacidad de nodulación (Caetano - Anollés et al. 1989; Munns, 1970). La adsorción de rizobios a los pelos radicales podría estar mediada por rhicadesina, una proteína cuyo anclaje a la superficie bacteriana es 50 dependiente de la presencia de Ca (Smit et al. 1991). Estas observaciones son consistentes con el uso de la práctica común del encalado de suelos ácidos que, además de corregir el pH, ejerce un efecto positivo sobre la supervivencia y desempeño simbiótico de los rizobios. De este modo, en el suelo cultivado con A. sativum L. (ajo), la posibilidad de capturar rizobios a través del cultivo trampa sería menor que en el suelo cultivado con S. tuberosum L. (papa), debido a que la cantidad de Ca hallada en ese suelo fue menor. Por otra parte se puede inferir que otras condiciones físico químicas del suelo, tales como las bajas temperaturas y contenido de materia orgánica que es medio (2 – 4%) según Gilabert et al. (1990) entre otras, no serían las más adecuados para permitir la sobrevivencia de estos microsimbiontes en el suelo, y podrían contribuir con la heterogeneidad de la distribución de la población rizobial en estos suelos. 1.2 Aislamiento y purificación en medio YMAmrc y medio glucosa peptona agar (GPA) (Ferrera Cerrato et al., 1993) Se obtuvieron 5 aislados rizobiales en principio: • Dos provenientes del suelo cultivado con Solanum tuberosum L., tratados con carbamatos, designados como ES1 y ES2. • Dos del suelo cultivado con Allium sativum L., tratados con carbamatos, indicados como EA1 y EA2. • Uno de la planta de Vicia faba L. de suelos no tratados con carbamatos: denominado EV1 (control). 51 1.2a.- Crecimiento en Medio YMAmrc Las colonias que originaron los aislados nodulares, presentaron abundante crecimiento en el medio YMAmrc (Anexo 5), siendo estas de color blanco o transparente, mucoides y de aspecto similar a las descritas para las colonias rizobiales (CIAT, 1988). 1.2b.-Crecimiento en medio glucosa- peptona, púrpura de bromocresol (GPA) Los aislados ES1 y EV1 presentaron un escaso crecimiento en el medio glucosa peptona, púrpura de bromocresol, manteniéndose el color morado, características que indican que se trata de bacterias rizobiales puras. En los demás aislados, ES2 del suelo de Solanum tuberosum L., EA1 y EA2 del suelo de Allium sativum L. se obtuvo un abundante crecimiento y el indicador viró de color morado a amarillo indicando la presencia de bacterias no rizobiales (Rodríguez y Ferrera– Cerrato, 1984). 2. Autenticación de los aislados en Macroptylium lathyroides L. y Phaseolus vulgaris L. Se mantuvieron 30 días las plantas en el umbráculo, un tiempo aceptable para permitir la nodulación y el reconocimiento de los rizobios, procediendo luego al desmontaje y la toma de los resultados (Tabla 13). 52 Tabla 13. Autenticación de los aislados de los suelos y nódulos en Macroptylium lathyroides L. y Phaseolus vulgaris L. Nodulación Posición color Posición Macroptylium Phaseolus Suelo lathyroides L. -Cultivado con S.tuberosum L.(papa) Tratado con carbamatos -Cultivado con A. sativum L. (ajo) Tratado con carbamatos -Cultivado con V. faba L.(haba) No tratado con carbamatos vulgaris L. color ES1: + ES2: - Blancos y marrones en todo el sistema radical ES1: + ES2: - Blancos y marrones en todo el sistema radical EA1: EA2: - _ EA 1: EA 2: - _ EV1: - _ EV1: + +: Presencia de nódulos. Blancos y en todo el sistema radical - : Ausencia de nódulos De los aislados del suelo cultivado con S. tuberosum L. tratados con carbamatos, resultó positiva la nodulación para el aislado ES1 en ambos hospedadores y ES2 negativa con ambas plantas. Los aislados del suelo cultivado con A. sativum L. tratados con carbamatos no formaron nódulos con ambos hospedadores al igual que el aislado ES2, por lo tanto, basándose en este resultado y por su crecimiento en el ensayo de purificación en medio GPA se descarta que sean rizobios y que la estructura similar a un nódulo obtenida en la captura de rizobios en el suelo cultivado con A. sativum L. se podría considerar como un nódulo incipiente, como se propuso anteriormente. El aislado de los nódulos de plantas de Vicia faba L. formó nódulos con las plantas de P. vulgaris L. mientras que con las plantas de M. lathyroides L. no presentó nodulación. A pesar de que ambos cultivos han sido designados como hospedadores de amplio rango hospedador (Bromfield y Barran 1990; Michiels, et al 1998) Odee et al., en 1996 sugirieron que P. vulgaris L. es nodulada por rizobios de rápido crecimiento (2-4 días para observar sus colonias) y Macropthylium atropurpureum por rizobios de 53 lento crecimiento (5-7 días para visualizar el crecimiento de sus colonias) (MatosCuzcano et al., 2004), por ende se sugiere que M. lathyroides L. no es una planta completamente no selectiva para la nodulación, como se obtuvo en los resultados de Marquina (2006), en donde en la prueba de infectividad, Crotalaria sp, superó los grados de infectividad presentados en el huésped M. lathyroides L. el cual no logró establecer simbiosis con todos los 12 aislados ensayados. Según estos resultados, se continuaron las demás pruebas solo con las bacterias que mostraron nodulación; el aislado (ES1) del suelo cultivado con S. tuberosum L. tratado con carbamatos y el aislado (EV1) proveniente de los nódulos de Vicia faba L., cultivada en suelos no tratados con carbamatos. En la Tabla 14 se describen las características de los rizobios que se obtuvieron de los aislados de nódulos formados en la prueba de autenticación en plantas de Phaseolus vulgaris L. y Macroptylium lathyroides L. Tabla 14. Características de las colonias aisladas del suelo tratado con carbamatos (Solanum tuberosum L.) y sin carbamatos (Vicia faba L.) Suelo Cultivado con Solanum tuberosum L. (papa), tratados con carbamatos Cultivado con Vicia faba L. (haba), no tratados con carbamatos Morfología de las colonias ‐ ES1: forma irregular, color blanco lechoso, aspecto opaco-brillante borde liso, textura muy mucoide, elevación convexa. Diámetro de la colonia a las 48h 3mm. EV1: forma irregular, color transparente, borde liso, aspecto brillante- traslúcida, textura muy mucoide, elevación convexa. Diámetro de la colonia a las 48h 4mm. Según las características macromorfológicas de las colonias de ambos rizobios se puede sugerir que en base a la clasificación de los géneros rizobiales expuestos por Marquina (2006), los dos aislados podrían pertenecer al género Rhizobium sp. Frank 1889. 54 3. Curvas de calibración del crecimiento de los aislados rizobiales ES1 y EV1. En la Figura 9 se observa que el aislado ES1 creciendo en un medio YMm líquido, alcanza el orden 108 UFC/ml a una Densidad Óptica de 0.5 aproximadamente, a comienzos de la fase exponencial, correspondiendo el mayor valor del título bacteriano a 7.6 x 108 obtenido a una DO=1.0, justo al final de la fase exponencial. Por lo tanto para los ensayos siguientes con el aislado ES1, no fue conveniente elegir una DO inferior a 0,5 y superior a 1.0, ya que no se encontrarían dentro de la UFC/ml 104 fase exponencial de crecimiento. 80000 70000 60000 50000 40000 30000 20000 10000 0 ES1 en YMm 0,02 0,09 0,39 0,58 0,75 0,95 1,1 1,4 D.O (600nm) Figura 9: Curva de calibración (UFC/ml Vs. D.O) en medio YMm para el aislado ES1. En la gráfica (Figura 10.) se observa que el aislado EV1 alcanza el orden 108 UFC/ml a una Densidad Óptica de 0,4 aproximadamente, a comienzos de la fase exponencial, correspondiendo el mayor valor del título bacteriano a 10x 108 obtenido a una DO= 0,85, justo al final de la fase exponencial. Por lo tanto en los ensayos posteriores realizados con el aislado EV1, no fue conveniente elegir una DO inferior a 0.4 y superior a 0.85. 55 120000 UFC/ml 104 100000 80000 60000 40000 EV1 en YMm 20000 1,1 0,85 0,75 0,58 0,5 0,21 0,14 0,04 0,02 0 D.O (600nm) Figura 10. Curva de calibración (UFC/ml Vs. D.O) en medio YMm para el aislado EV1. 4. Prueba cualitativa de viabilidad de los aislados rizobiales en presencia de carbamatos. Esta prueba fue cualitativa, de manera que los resultados de este ensayo fueron determinantes para los demás experimentos, en consecuencia como se muestra en las Tablas 15 y 17, los ensayos realizados con los aislados que no presentaron halo de inhibición, se consideran resistentes a ese carbamato (Ferrera et al., 1993) y a las que lo presentaron se les determinó el tamaño del halo de inhibición alrededor del papel de filtro (Tablas 16 y 18) (Anexo 6). 56 Tabla 15. Prueba de viabilidad en presencia de carbamatos, del aislado rizobial (ES1) proveniente del suelo cultivado con Solanum tuberosum L. (tratado con agroquímicos) Pesticida Carbodan Dithane Previcur Vondozeb Zineb Control Crecimiento cualitativo Diluciones del pesticida carbamato Original 1/2 1/4 + ++ + + ++ + + + + ++ 1/6 + + + ++: Abundante crecimiento. + : Crecimiento ‐ : No hubo crecimiento Concentraciones de las diluciones Original; ½; ¼; 1/6 Carbodan (μl/100ml ): 165; 83; 41; 27.5. Dithane (g/100ml ): 0.25; 0.125; 0.063; 0.041 Previcur (μl/100ml ): 125; 62.5; 31.3; 20.8. Vondozeb (g/100ml ): 0.25; 0.125; 0.063; 0.041 Zineb (g/100ml ): 0.25; 0.125; 0.063; 0.041. Control (100ml ) Tabla 16. Tamaño del halo de inhibición (cm) de crecimiento celular alrededor del papel filtro para el aislado ES1. Tamaño del halo de inhibición ( cm) Pesticida Diluciones del pesticida carbamato Original 1/2 1/4 1/6 Carbodan Dithane 2,5 1,5 1 1 Previcur Vondozeb 2,2 1,2 1,1 0,8 Zineb Control ‐ : No hubo halo de inhibición Concentraciones de las diluciones Original; ½; ¼; 1/6 Carbodan (μl/100ml ): 165; 83; 41; 27.5. Dithane (g/100ml ): 0.25; 0.125; 0.063; 0.041 Previcur (μl/100ml ): 125; 62.5; 31.3; 20.8. Vondozeb (g/100ml ): 0.25; 0.125; 0.063; 0.041 Zineb (g/100ml ): 0.25; 0.125; 0.063; 0.041. Control (100ml ) 57 Tabla 17. Prueba de viabilidad en presencia de carbamatos, del aislado rizobial (EV1) proveniente de nódulos de Vicia faba L. cultivada en suelos sin tratamientos de agroquímicos. Pesticida Carbodan Dithane Previcur Vondozeb Zineb Control Crecimiento cualitativo Diluciones del pesticida carbamato Original 1/2 1/4 + ++ + + ++ + + + + ++ 1/6 + + + ++: Abundante crecimiento. +: Crecimiento. - : No hubo crecimiento Concentraciones de las diluciones Original; ½; ¼; 1/6 Carbodan (μl/100ml ): 165; 83; 41; 27.5. Dithane (g/100ml ): 0.25; 0.125; 0.063; 0.041 Previcur (μl/100ml ): 125; 62.5; 31.3; 20.8. Vondozeb (g/100ml ): 0.25; 0.125; 0.063; 0.041 Zineb (g/100ml ): 0.25; 0.125; 0.063; 0.041. Control (100ml ) El aislado EV1 presentó crecimiento en el tratamiento con Zineb en todas sus concentraciones, formando un pequeño halo de inhibición alrededor del papel filtro, cuyas dimensiones se señalan en la (Tabla 18), sin embargo, no fue tomado en cuenta este carbamato para elaborar la prueba cuantitativa de las curvas de crecimiento, ya que en términos comparativos, el resultado no era similar al del aislado ES1 (Tabla 16), pero si se consideró para la prueba de uso de fuentes carbonadas y nitrogenadas y para el ensayo de la germinación de semillas de Vicia faba L. De acuerdo a estos resultados, los tratamientos con carbamatos, en los cuales los aislados rizobiales presentaron abundante crecimiento fueron Carbodan ½ y Previcur ½ y el control. En vista, que en los dos aislados EV1 y ES1, la respuesta de crecimiento fue similar, esta apariencia fue discernida en las curvas de crecimiento y el conteo viable (plaqueo). 58 Tabla 18. Tamaño del halo de inhibición del crecimiento celular alrededor del papel filtro para el aislado EV1. Tamaño del halo de inhibición ( cm) Diluciones del pesticida carbamato Pesticida Carbodan Dithane Previcur Vondozeb Zineb Control Original 1/2 1/4 1/6 2,7 2,7 1 - 2,7 2,7 0,5 - 2 2,7 0,5 - 1,5 2,7 0,5 - - : No hubo halo de inhibición Concentraciones de las diluciones Original; ½; ¼; 1/6 Carbodan (μl/100ml ): 165; 83; 41; 27.5. Dithane (g/100ml ): 0.25; 0.125; 0.063; 0.041 Previcur (μl/100ml ): 125; 62.5; 31.3; 20.8. Vondozeb (g/100ml ): 0.25; 0.125; 0.063; 0.041 Zineb (g/100ml ): 0.25; 0.125; 0.063; 0.041. Control (100ml ) 5. Prueba de utilización de carbamatos como única fuente carbonada y nitrogenada. Basándose en la resistencia que tienen los rizobios ES1 y EV1 a los pesticidas usados en la prueba de viabilidad cualitativa, se seleccionaron los carbamatos Previcur, bacteriales. Carbodan y Zineb, por permitir el crecimiento de estos aislados Por lo tanto, se realizó un ensayo para corroborar si los aislados rizobiales eran capaces de emplear los 3 carbamatos como única fuente carbonada o nitrogenada, demostrándose que los microsimbiontes ES1 y EV1 son incapaces de metabolizarlos y utilizarlos como fuente de carbono y nitrógeno, ya que solo hubo crecimiento en el control positivo de ambos aislados que contenía manitol y NH4NO3 como fuente de carbono y nitrógeno respectivamente. Los resultados se muestran en las Tablas 19, 20, 21 y 22. 59 Tabla 19. Utilización de carbamatos como única fuente carbonada para el aislado ES1 proveniente del suelo cultivado con Solanum tuberosum L. Fuente de carbono Carbodan Zineb Previcur Control positivo (manitol) Control negativo + Crecimiento. Crecimiento D.O (600nm) 0.03 0.02 0.02 + 1.0 0.02 - No hubo crecimiento Tabla 20. Utilización de carbamatos como única fuente nitrogenada para el aislado ES1 proveniente del suelo cultivado con Solanum tuberosum L. Fuente de nitrógeno Crecimiento D.O (600nm) Carbodan 0.02 Zineb 0.02 Previcur 0.05 Control positivo (manitol) + 1.0 Control negativo 0.02 + Crecimiento. - No hubo crecimiento Tabla 21. Utilización de carbamatos como única fuente carbonada para el aislado EV1 del suelo cultivado con Vicia faba L. Fuente de carbono Carbodan Zineb Previcur Control positivo (manitol) Control negativo + Crecimiento. Crecimiento D.O (600nm) 0.02 0.02 0.02 + 1.0 0.02 - No hubo crecimiento Tabla 22. Utilización de carbamatos como única fuente nitrogenada para el aislado EV1 del suelo cultivado con Vicia faba L. Fuente de nitrógeno Carbodan Zineb Previcur Control positivo (manitol) Control negativo + Crecimiento. Crecimiento D.O (600nm) 0.02 0.02 0.02 + 1.0 0.02 - No hubo crecimiento 60 Con estos resultados se rechaza una de las hipótesis planteadas, ya que se comprobó que estos aislados no son capaces de metabolizar estos carbamatos, al no utilizarlos como fuentes de carbono ni de nitrógeno. Sin embargo, sí se conoce que los utilizan varios géneros bacteriales, tales como: algunas cepas de Pseudomonas sp. aisladas de suelos de jardín que son capaces de metabolizar el carbaryl como fuente carbonada, demostrado por la medición fluorométrica de la producción de 1- naphtol como resultado de la hidrólisis del Carbaryl (Larkin et al., 1985, Chapalamadugu et al., 1991), Arthrobacter sp. cepa RC100 por Hayatsu et al., 1999 y Rhodococcus sp. (NCIB 12038) reportada por Larkin et al., 1985., aisladas de suelos tratados con carbaryl que degradan ese pesticida carbamato y los usan como fuente carbonada, y en el caso de la NCIB también como fuente nitrogenada. De igual manera Feng et al. (1997), quienes aislaron bacterias del género Sphingomonas sp. cepa CF06 de suelos tratados con carbofurano y con la habilidad de usar este carbamato como fuente de carbono y nitrógeno en la cual están implicados los plásmidos pCF01–pCF05 de esa bacteria, y específicamente en el género Rhizobium sp. la cepa AC100 sp aislada de suelos tratados con carbaryl estudiada por Hashimoto et al., en el 2001, la cual es capaz de degradar este carbamato, y usarlo como fuente de carbono. Resultados similares fueron obtenidos por Castro (2008). Este estudio permitió suponer la presencia de un cometabolismo en los aislados de suelos sometidos a tratamiento agroquímico que no fueron capaces de utilizar el Acarin (la forma comercial del acaricida Dicofol) como fuente carbonada, pero sí lo degradaron al estar presente el glicerol como fuente de carbono más el acaricida. 61 6. Prueba cuantitativa de viabilidad de los aislados rizobiales en presencia de carbamatos. En la Figura 11 se observa el crecimiento (aumento de DO) del aislado ES1 proveniente de suelos tratados con carbamatos, en función del tiempo, 2 cultivos rizobiales, uno tratado con Carbodan y el otro con Previcur, cada uno diluido a ½ de la concentración original empleada en el campo, comparando con el control sin agroquímico. Se observa una curva sigmoide típica del crecimiento celular bacteriano, con la fase lag, que transcurre durante 6 horas aproximadamente para todos los tratamientos. La fase exponencial de los cultivos que crecen en YMm + Previcur ½ y YMm + Carbodan ½, presentaron mayor pendiente en relación al control sin carbamatos en YMm, significando un crecimiento más apresurado, el cual es más pronunciado en los cultivos que contiene Previcur, que alcanza la fase estacionaria más rápidamente a las 29 h aproximadamente, seguido por el tratamiento con Carbodan (31 h) y el cultivo sin carbamato (39 h).En todos los tratamientos el aislado obtuvo un máximo de DO. = 1,45. Densidad óptica (600nm) 1,6 1,4 1,2 1 ES1 en YMm 0,8 ES1 en YMm + Carbodan 1/2 0,6 0,4 ES1 en YMm + Previcur 1/2 0,2 0 0 6 11 14,5 18 23 29 35,5 45 Tiempo (h) Figura 11. Curva de crecimiento del aislado ES1, proveniente del suelo tratados con carbamatos, cultivado con Solanum tuberosum L. (papa), creciendo en medio YMm en presencia de carbamatos. Las concentraciones ½ para Carbodan y Previcur corresponden a 12,45μl/15ml YMm y 9,3 μl/15ml YMm respectivamente. 62 Densidad óptica (600nm) 1,2 1 0,8 EV1 en YMm 0,6 0,4 EV1 en YMm + Carbodan 1/2 0,2 EV1 en YMm + Previcur 1/2 0 0 4 8 12 15 17 20 22 45 53 Tiempo (h) Figura 12. Curva de crecimiento del aislado EV1, proveniente del suelo no tratado con carbamatos cultivado con Vicia faba L. (haba), creciendo en medio YMm en presencia de carbamatos. Las concentraciones ½ para Carbodan y Previcur corresponden a 12, 45μl/15ml YMm y 9,3 μl/15ml YMm respectivamente. En la Figura 12 se observa que el crecimiento del aislado EV1, proveniente del suelo no tratado con carbamatos, presenta de igual manera que el aislado ES1 procedente del suelo tratado con carbamatos, un comportamiento sigmoidal con la fase lac, más breve aproximadamente 4 h en todos los tratamientos. La fase exponencial de los cultivos que crecen en YMm + Previcur ½ y YMm + Carbodan ½, exhibieron la misma pendiente, indicando que el crecimiento ocurre a la misma velocidad en ambos tratamientos, pero son más acelerados en relación al control sin carbamatos en YMm, alcanzando rápidamente entre las 9 y 10 h la fase estacionaria con una DO máxima = a 0.52, que se mantiene durante el transcurso del experimento (60 h). Por el contrario en el tratamiento sin carbamatos, la fase exponencial transcurre a menor velocidad prolongándose hasta las 53 h, cuando alcanza la fase estacionaria con una DO máxima de 1.1. En la Figura 11 y 12 se observa una promoción del crecimiento de los dos aislados ES1 y EV1, cuando son cultivados en un medio que les proporciona las fuentes de carbono (manitol y sacarosa) y además tiene el carbamato, superando al 63 crecimiento rizobial del tratamiento control, que crece en presencia de las mismas fuentes carbonadas sin carbamatos. Es importante referirse a los resultados negativos que se obtuvieron en el ensayo de los carbamatos como únicas fuentes carbonadas y nitrogenadas (Tablas 19, 20, 21 y 22), en donde no hubo crecimiento bacteriano, permitiendo sugerir que los aislados EV1 y ES1, en presencia de las fuentes carbonadas (manitol y sacarosa) son capaces de biodegradar el Previcur (9,3μl/15ml YMm) y el Carbodan (12,45μl/15ml YMm) en diferente medida, bajo un proceso denominado cometabolismo, definido como la transformación de un compuesto llamado cosustrato, en presencia obligada de un sustrato durante el crecimiento o por células en reposo en ausencia del sustrato de crecimiento (Perry, 1979), el cual requiere agregar un factor iniciador del crecimiento bacteriano que en este caso será las fuentes carbonadas, que induzcan la producción de enzimas que puedan transformar estos compuestos carbamatos. Resultados similares se han encontrado en bacterias del género Brevibacterium sp., en una investigación realizada por Luo et al. (2008), quienes detectaron el cometabolismo del Bensulfuron-methyl (BSM) en presencia de varias fuentes de carbono nitrógeno y fósforo, determinándose la mejor respuesta, usando el lactato de sodio como fuente de carbono con un 79,5% de degradación del BSM. En este trabajo se observó que el aislado ES1 presenta una resistencia a los agroquímicos Previcur y Carbodan en una dilución correspondiente a ½ de la concentración empleada en el campo (9,3 μl/15ml YMm y 12,45μl/15ml YMm respectivamente). El aislado EV1, se considera como un aislado más sensible a las mismas concentraciones de estos carbamatos, pese a tener un crecimiento inicial más acelerado en presencia de ellos, ya que hay un punto a una DO = 0.52, en donde se está tornando tóxica la presencia del compuesto transformado o cometabolizado, tal como se podrá evidenciar más adelante en el conteo viable. Debido al número de repeticiones en esta prueba, se utilizó el análisis de comparación de medidas repetidas para la prueba estadística, que permitiera 64 establecer, si había diferencias significativas entre los tratamientos dentro del mismo aislado rizobial. Dicha prueba se hizo usando el programa computacional SPSS, el cual arrojó en la prueba de Post hoc diferencias significativas a nivel 0,05 para el aislado ES1 entre todos los tratamientos, y para el aislado EV1 diferencias significativas entre el control y los dos tratamientos con carbamatos (Tabla 23 y 24 respectivamente). Tabla 23. Comparaciones múltiples. Pruebas de post hoc (Curvas D.O/ Tiempo) para ES1. Tratamiento Tratamiento (i) (j) Carbodan Control Previcur Control Previcur Carbodan Previcur Carbodan Control Diferencia entre medias (i‐j) Error típ. Significación Intervalo de confianza al 95% Límite inferior Límite superior ,0998(*) ‐,1291(*) ,02306 ,02306 ,023 ,011 ,0264 ‐,2025 ,1732 ‐,0557 ,1647(*) ‐,2289(*) ,02306 ,02306 ,023 ,002 ‐,1732 ‐,3023 ‐,0264 ‐,1555 ,1291(*) ,2289(*) ,02306 ,02306 ,011 ,002 ,0557 ,1555 ,2025 ,3023 Basado en las medias observadas. * La diferencia de medias es significativa al nivel ,05. Tabla 24. Comparaciones múltiples. Pruebas de post hoc (Curvas D.O/ Tiempo) para EV1. Tratamiento (i) Carbodan Control Previcur Tratamiento (j) Diferencia entre medias (i‐j) Error típ. Significación Límite inferior Límite superior Control Previcur Carbodan Previcur Carbodan Control ‐,1647(*) ,0103 ,01375 ,01375 ,001 ,510 ‐,2085 ‐,0335 ‐,1210 ,0540 ,1647(*) ,1750(*) ,01375 ,01375 ,001 ,001 ,1210 ,1312 ,2085 ,2188 ‐,0103 ‐,1750(*) ,01375 ,01375 ,510 ,001 ‐,0540 ‐,2188 ,0335 ‐,1312 Basado en las medias observadas. Intervalo de confianza al 95% * La diferencia de medias es significativa al nivel ,05. 65 En las siguientes gráficas (Figuras 13 y 14) se muestra el conteo viable para ambos aislados en medio YMm y con la adición a este medio de los carbamatos Carbodan y Previcur en la dilución ½. 160000 140000 UFC/ml 104 120000 100000 ES1 en YMm 80000 ES1 en YMm + Carbodan 1/2 60000 40000 ES1 en YMm + Previcur 1/2 20000 0 0 4 11 13,5 15,5 18 23 29 33,5 Tiempo (h) Figura 13. Unidades formadoras de colonias del aislado ES1, proveniente del suelo tratados con carbamatos y cultivado con Solanum tuberosum L. (papa), creciendo en medio YMm en presencia de carbamatos. Las concentraciones ½ para Carbodan y Previcur corresponden a 12,45μl/15ml YMm y 9,3 μl/15ml YMm respectivamente. Acorde con el comportamiento del aislado ES1 en la curva de crecimiento de la Figura 11, y su correspondiente conteo viable, representado en la Figura 13, se observa una promoción del crecimiento celular mayor en los tratamientos con carbamatos que en el control, siendo el que contiene Previcur el que produjo un mayor número de unidades formadoras de colonia por mililitro (UFC/ml). Estas cantidades máximas fueron 14 x 108 a las 16,5 h en el tratamiento con Previcur, 11 x 108 a las 27 h con Carbodan y 7,6 x 108 a las 29 h con el control (YM). Así como el Previcur promovió el incremento de la población bacteriana hasta aumentar incluso de orden (109), en un corto tiempo a las 13,5 h aproximadamente, también disminuyó bruscamente registrando valores más bajos (2,5 x 108) que el control a las 29 h, justamente cuando este tratamiento obtuvo su máxima cantidad. Las fases de la curva de crecimiento observables en la Figura 13 y 14 se corresponden a la fase lag, exponencial y fase de muerte, en donde es importante destacar que la fase estacionaria no es observable posiblemente debido a que el medio de crecimiento 66 usado (YMm) no es un medio completamente rico, en donde no hay tantos sustitutos de las fuentes de carbono, que al finalizar la fase estacionaria disminuyen en el medio y se produce la muerte celular por agotamiento de nutrientes. 120000 UFC/ml 104 100000 80000 EV1 en YMm 60000 40000 EV1 en YMm + Carbodan 1/2 20000 EV1 en YMm + Previcur 1/2 0 0 2 4 6 8 10 14 15 19 21 45 Tiempo (h) Figura 14. Unidades formadoras de colonias del aislado EV1, proveniente del suelo no tratado con carbamatos, cultivado con Vicia faba L. (haba), creciendo en medio YMm en presencia de carbamatos. Las concentraciones ½ para Carbodan y Previcur corresponden a 12,45μl/15ml YM y 9,3 μl/15ml YM respectivamente. En la Figura 14 se muestra la curva de crecimiento (UFC/ml Vs. Tiempo) para el aislado EV1. Observándose al final de la fase exponencial de los tratamientos con carbamatos (correspondiente a la zona media de la fase exponencial del tratamiento control) que el crecimiento celular se detiene y comienza a disminuir rápidamente (a las 10 h), tanto para los tratamientos con Carbodan ½ como con Previcur ½, mientras que en el medio YMm sin carbamato este aislado continúa creciendo hasta las 21h y luego si comienza a decrecer su división celular. Los mayores valores de UFC/ml obtenidos fueron 3,3 x 108 para Previcur, 4,5 x 108 para Carbodan y 10 x 108 para el control, donde prácticamente se duplica el título bacteriano. De tal manera que se observa un decaimiento del conteo viable al final de la fase exponencial para los tratamientos con carbamatos, acorde con el resultado de la curva D.O Vs. Tiempo (Figura 12), quedando evidenciado primeramente que el aislado EV1 es capaz de 67 cometabolizar ambos carbamatos , debido al crecimiento acelerado que presentan los cultivos rizobiales en medio YMm en presencia de los carbamatos y en segundo lugar se puede sugerir que el compuesto que se origina por trasformación a partir del carbamato (cometabolismo), origina toxicidad en el medio, causando la muerte celular de este aislado rizobial. De igual manera debido al número de repeticiones en esta prueba, se utilizó el análisis de comparación de medidas repetidas para la prueba estadística que lograra establecer si había diferencias significativas entre los tratamientos dentro del mismo aislado rizobial, el cual arrojó en la prueba de Post hoc diferencias significativas a nivel 0,05 para el aislado ES1 y EV1 entre todos los tratamientos (Tabla 25 y 26). Tabla 25. Comparaciones múltiples. Pruebas de post hoc (Curvas UFC/ml/ Tiempo) para ES1. Tratamiento (i) Tratamiento (j) Diferencia entre medias (i-j) Error típ. Significación Carbodan Control Previcur Carbodan Previcur Carbodan Control 17916,3861(*) ‐20138,2929(*) 1920,44878 1920,44878 ,000 ,000 13217,2172 ‐24837,4618 22615,5550 ‐15439,1240 ‐17916,3861(*) ‐38054,6790(*) 1920,44878 1920,44878 ,000 ,000 ‐22615,5550 ‐42753,8479 ‐13217,2172 ‐33355,5101 20138,2929(*) 38054,6790(*) 1920,44878 1920,44878 ,000 ,000 15439,1240 33355,5101 24837,4618 42753,8479 Control Previcur Basado en las medias observadas. Intervalo de confianza al 95% Límite Límite inferior superior * La diferencia de medias es significativa al nivel ,05 68 Tabla 26. Comparaciones múltiples. Pruebas de post hoc (Curvas UFC/ml/ Tiempo) para EV1. Tratamiento (i) Tratamiento (j) Diferencia entre medias (i-j) Error típ. Significación Carbodan Control Previcur Carbodan Previcur Carbodan Control -10399,8782(*) 7142,0673(*) 10399,8782(*) 17541,9455(*) -7142,0673(*) -17541,9455(*) 710,02371 710,02371 710,02371 710,02371 710,02371 710,02371 ,000 ,000 ,000 ,000 ,000 ,000 Control Previcur Basado en las medias observadas. Intervalo de confianza al 95% Límite Límite inferior superior -12137,2436 -8662,5128 5404,7019 8879,4327 8662,5128 12137,2436 15804,5800 19279,3109 -8879,4327 -5404,7019 -19279,3109 -15804,5800 * La diferencia de medias es significativa al nivel ,05 7. Dosis Letal media (DL50) Como antepenúltimo ensayo se determinó la Dosis Letal media (DL50) de los carbamatos usados en la prueba de viabilidad de rizobios en presencia de ellos. En base al ejemplo mostrado en Miranda (2006) para calcular la DL50 se realizó una regresión para conocer la recta que más se ajustara y de ahí hacer los despejes de las ecuaciones resultantes de esas regresiones. Para esto fue necesario usar los programas de computación Maple 11 y Matlab R2007a, y la tendencia polinómica, arrojó ecuaciones de 3er grado para hallar las dosis de ES1 con Carbodan y EV1 con Carbodan y Previcur y 4to grado para ES1 con Previcur ya que en esta fue necesario usar 5 concentraciones para hallar la DL50. Se tomó el mayor número de UFC/ml como el 100% de la población y la mitad de este valor como el 50% de la población, para la cual había que hallar la dosis. Los resultados se presentan en las Figuras 15, 16, 17 y 18. 69 10000 UFC/ml 104 8000 6000 ES1 YM + Carbodan 4000 Polinómica (ES1 YM + Carbodan) 2000 0 83 165 206 248 DL50=200.81µl/100ml Concentraciones (µl/100ml) Figura 15. Dosis letal media de Carbodan para el aislado rizobial ES1del suelo tratado con carbamatos y cultivado con Solanum tuberosum L. En la Figura 15, se observa que el 50% de la población corresponde a 4350x104 UFC/ml y la DL50 hallada para ocasionar la muerte de esta población fue 200.81 μl/ UFC/ml 104 100ml. 14000 12000 10000 8000 6000 4000 2000 0 ES1 YM + Previcur Polinómica (ES1 YM + Previcur) 62,5 125 156,3 187,5 250 Concentraciones (µl/100ml) DL50=214.14µl/100ml Figura 16. Dosis letal media de Previcur para el aislado rizobial ES1 del suelo tratado con carbamatos y cultivado con Solanum tuberosum L. En la Figura 16, se observa que el 50% de la población corresponde a 6550x104 UFC/ml y la DL50 hallada para ocasionar la muerte de esta población fue 214.14 μl/ 100ml. 70 16 14 12 10 8 6 4 2 0 UFC/ml 104 EV1 YM + Carbodan Polinómica (EV1 YM + Carbodan) 27,5 41 83 165 Concentraciones (µl/100ml) . DL50=123.35µl/100ml Figura 17. Dosis letal media de Carbodan para el aislado rizobial EV1 del suelo no tratado con carbamatos y cultivado con Vicia faba L. En la Figura 17, se observa que el 50% de la población corresponde a 7.3 x 104 UFC/ml y la DL50 hallada para ocasionar la muerte de esta población fue 123.35 μl/ UFC/ml 104 100ml. 20 18 16 14 12 10 8 6 4 2 0 20,8 31,3 62,5 125 DL50=108.06µl/100ml Concentraciones (μl/100ml ) Figura 18. Dosis letal media de Previcur para el aislado rizobial EV1 del suelo no tratado con carbamatos cultivado con Vicia faba L. En la Figura 18, se observa que el 50% de la población corresponde a 8.55 x 104 UFC/ml y la DL50 hallada para ocasionar la muerte de esta población fue 108.06 μl/ 100ml. 71 En estos resultados se puede observar que las DL50 halladas para el aislado ES1 tienen un valor mayor en ambos carbamatos que las DL50 calculadas para el aislado EV1 creciendo en presencia de los mismos carbamatos, demostrando que efectivamente la bacteria ES1, que estuvo bajo presiones de selectividad por el tratamiento químico de ese suelo, ha desarrollado una mayor resistencia ante estos compuestos que el aislado EV1 que no ha estado sometido a tratamiento químico, al menos durante los 12 años precedentes al aislamiento. 8. Prueba de germinación de Vicia faba L. en presencia de carbamatos. Los resultados mostrados en la Figura 19, indican que el mayor porcentaje de germinación fue con el tratamiento Previcur en su concentración original correspondiente a un 77,5% de germinación, incluso por encima del porcentaje del tratamiento control que fue de 72,5%. Sin embargo, entre ellos no hubo diferencias significativas, como se denota en la Figura 19 con las letras a y ab, y en la Tabla 27. Por lo tanto, no se puede sugerir que el tratamiento con Previcur en su concentración original estimula la germinación de la semilla, sino que simplemente no la afecta. No obstante la respuesta no inhibitoria que presentaron las semillas con este tratamiento podría deberse a que este pesticida es un fungicida de acción sistémico-estimulante (Indice agropecuario, 2001). 72 % total de germinación 90 9 80 8 70 7 60 6 50 5 40 4 30 3 20 2 10 1 0 a ab T Tratamientos Figura a 19. Porce entaje total de d germinacción de las semillas s de Vicia faba L. L a los 6 día as de 1 acuerd do a los trattamientos. Tratamientos T s µl ó g/400ml de Sol. Hoagland H /4: Carbodan n orig: 1 660 µl; Carbodan ½: 330µl; Carbodan C /4: 165µl; Ca arbodan 1/6: 110µl. Pre evicur orig: 500µl; 5 Previccur ½: 250µ µl; Previcur ¼: 125µl; Previcur P 1/6 6: 83,3µl. Zin neb orig: 1g g; Zineb ½: 0,5g; Zineb ¼: 0,25g; Zineb 1/6: 0,1 16g. Control: 400ml Sol. Hoagland ¼. ¼ Mientras tanto la mayoría de los otrros tratamie entos sí affectaron ne egativamen nte la germinación com mo se muesstra en la Figura F 19 y en la Tablla 27 en do onde se obsserva que el e análisis estadísticco mediantte el ANO OVA establece que hay h diferen ncias signifiicativas enttre los trata amientos, id dentificándo ose en el contraste c múltiple de rango r con letras diferrentes y un u desplazzamiento a otra colu umna formando parte e de difere entes grupo os homogé éneos. C Con estos resultadoss se acep pta la hipó ótesis propu uesta de que altas con ncentraciones de carba amatos (po or lo menoss de Carbod dan y Zineb b) pueden in nhibir la germinación de d las semillas de Viccia faba L., debido a que el meno or porcentajje de germ minación fue precisam mente en lo os tratamie entos Carbodan original (25%) y Zineb original (30%) y son estadísticamente diferente es con resp pecto ontrol. El tratamiento o estadísticco para estta prueba consistió en e un al trattamiento co ANOV VA simple realizado r co on el progra ama Statgra aphics. 73 Tabla 27. ANOVA simple para resultados de germinación de V. faba L. con p < 0,05 y contraste múltiple de rango con un nivel de confianza del 95,0% según los tratamientos. Análisis de la Varianza -----------------------------------------------------------------------------Fuente Sumas de cuad. Gl Cuadrado Medio Cociente-F P-Valor -----------------------------------------------------------------------------Entre grupos 12026,9 12 1002,24 2,88 0,0061 Intra grupos 13550,0 39 347,436 -----------------------------------------------------------------------------Total (Corr.) 25576,9 51 Contraste Múltiple de Rango para germinacion según tratamiento -------------------------------------------------------------------------------Método: 95,0 porcentaje LSD tratamiento Frec. Media Grupos homogéneos -------------------------------------------------------------------------------Carbodan Origin4 25,0 e zineb original-4 30,0 ed zineb 1/2-6 4 32,5 edc zineb 1/6 -6 4 35,0 edc Carbodan 1/2-6 4 37,5 edc Carbodan 1/6-6 4 42,5 edc zineb 1/4 -6 4 45,0 edc Previcur 1/6-6 4 45,0 edc Previcur 1/4-6 4 50,0 edcb Carbodan 1/4-6 4 55,0 dcba Previcur 1/2-6 4 57,5 cba Control-6 4 72,5 ba Previcur origin4 77,5 a -------------------------------------------------------------------------------- 9. Pueba de nodulación y crecimiento de Vicia faba L. en presencia de carbamatos. Por último se hizo esta prueba a nivel de umbráculo, y luego de 50 días de crecimiento de las plantas de V. faba L. después de la inoculación con los dos aislados rizobiales ES1 y EV1 y el carbamato Previcur, se desmontaron y se midieron los parámetros mostrados en las siguientes Figuras. 74 18 Número de nódulos/planta 16 14 12 10 8 6 4 2 0 Control ES1 EV1 P + EV1 Tratamientos P + ES1 Previcur Figura 20. Número de nódulos formados en plantas de Vicia faba L. Control (sin aislado y sin Previcur), ES1 (aislado ES1 sin Previcur), EV1 (aislado EV1 sin Previcur), P+EV1 (Previcur + EV1), P+ES1 (Previcur + ES1), Previcur (Previcur sin aislado). En la Figura 20, se observa el número de nódulos formados en las plantas de Vicia faba L. de acuerdo a los tratamientos. En otras palabras se puede ver la infectividad de los aislados rizobiales. El aislado ES1 resultó no ser infectivo con este hospedador, ya que no formó nódulos ni en el tratamiento ES1 (inoculado con la bacteria, sin carbamato) ni en el tratamiento P+ES1 (inoculado con el carbamato + aislado ES1). Este resultado no es de extrañar si se compara con los resultados obtenidos en otras investigaciones como la de Mutch y Young (2004), en donde observan la especificidad de Rhizobium leguminosarum biovar viciae, con cultivos de varias leguminosas entre las cuales está la de nuestro interés que es Vicia faba L. y obtienen que ésta es una planta bastante específica ya que fue nodulada en gran cantidad por bacterias aisladas precisamente de ese mismo cultivo (10 nódulos por planta) mientras que fue escasamente nodulada por bacterias aisladas a partir de otras plantas (entre 1 y 4 nódulos por planta dependiendo del hospedador original) en total fue infectada por solo un 32,5% de los 625 aislados rizobiales. Un ejemplo de la deficiente nodulación de Rhizobium leguminosarum biovar viciae con este cultivo fue estudiado por Esser-Mönning et al. (1995), con dos líneas puras de Vicia faba L. l 40 y l 25, en donde asociaron la falta de infectividad al control que ejerce un 75 gen dominante (Sym-2) para la línea l 40, y un gen recesivo (sym-3) para la línea l 25, siendo el gen Sym-2 epistático del gen sym-3. Por otra parte, el aislado EV1 sí noduló a Vicia faba L. como era de esperarse, ya que esta bacteria fue aislada a partir de nódulos eficientes de estas plantas (Anexo 11). En las plantas del tratamiento EV1 (inoculadas con el aislado EV1 sin carbamato), se obtuvo un total de 92 nódulos de color blanco con un promedio por planta de 15 nódulos, a un total de 9 nódulos de color verde con un promedio de 2 nódulos por planta, en su mayoría situados en la parte central del sistema radical. Mientras que para el tratamiento P+EV1 (inoculadas con el carbamato + aislado EV1), se obtuvo un total de 29 nódulos de color blanco con un promedio por planta de 5 nódulos, a un total de 12 nódulos de color marrón con un promedio de 2 nódulos por planta, en su mayoría ubicados en la zona terminal del sistema radical. Por estas características, se puede sugerir que el aislado no fue efectivo, ya que no se encontraron nódulos de color rojizo en ambos tratamientos, lo que indica que no hubo actividad fijadora de N2 debido posiblemente a la ausencia de leghemoglobina ya que ésta es de color rojo, y que se observó en la formación de nódulos de color blanco característica de nódulos inefectivos (Lodeiro et al., 2003). Mientras tanto, la coloración verde obtenida en algunos nódulos del tratamiento EV1 (inoculadas con el aislado EV1 sin carbamato) es característica de los nódulos que están comenzando a desintegrarse (Amadeo, 2006). Referente a la coloración marrón de algunos de los nódulos obtenidos en el tratamiento P+EV1 (inoculadas con el carbamato + aislado EV1), y a su ubicación terminal en el sistema radical, se puede inferir que la aplicación del carbamato Previcur en su concentración original (aplicada en campo) afectó desfavorable la nodulación, ya que el color marrón de los nódulos es característico de los nódulos senescentes o ya degradados (Lodeiro et al., 2003; Amadeo, 2006), y la ubicación en la zona terminal de la raíz indican que la nodulación se produjo cuando ya el carbamato Previcur estaba más diluído por los riegos efectuados durante la realización de la prueba de nodulación en el umbráculo, y permitió que el rizobio EV1 lograra infectar las raíces nuevas de las plantas de Vicia faba L. 76 En este sentido, con el análisis estadístico ANOVA simple se encontraron diferencias significativas entre ambos tratamientos (Tabla 28), lográndose aceptar la segunda hipótesis planteada, en donde se supuso que la aplicación de altas concentraciones de carbamatos podría afectar desfavorablemente la nodulación de esta planta, acorde con la información de Alexander (1977), quien señala que la nodulación por Rhizobium sp. en las leguminosas se anula o reduce por pesticidas, ya que en principio éstos afectan la viabilidad de la bacteria además de la capacidad para nodular raíces, y en consecuencia la posibilidad de la Fijación Biológica de Nitrógeno. Esto en contraste a los resultados obtenidos por Aamil et al. (2004), quienes mostraron que las semillas de Cicer arietinum tratadas con bajas concentraciones de pesticidas producían plantas en las que la nodulación se incrementaba significativamente, y las que eran tratadas con altas concentraciones de pesticidas tenían una reducción en la nodulación con Mesorhizobium sp.. 77 Tabla 28. ANOVA simple para resultados de Nº de nódulos de V. faba L. con p < 0,05 y contraste múltiple de rango con un nivel de confianza del 95,0%, según los tratamientos. Análisis de la Varianza -----------------------------------------------------------------------------Fuente Sumas de cuad. Gl Cuadrado Medio Cociente-F P-Valor -----------------------------------------------------------------------------Entre grupos 1420,22 5 284,044 8,63 0,0000 Intra grupos 987,667 30 32,9222 -----------------------------------------------------------------------------Total (Corr.) 2407,89 35 Contraste Múltiple de Rango para Nº de Nódulos según Tratamientos -------------------------------------------------------------------------------Método: 95,0 porcentaje LSD Tratamientos Frec. Media Grupos homogéneos -------------------------------------------------------------------------------PREVICUR 6 0,0 c CONTROL 6 0,0 c ES1 6 0,0 c P+ES1 6 0,0 c P+EV1 6 6,83333 b EV1 6 16,8333 a -------------------------------------------------------------------------------- En la Figura 21 se muestran los pesos frescos y secos de los nódulos obtenidos para cada tratamiento, específicamente para los tratamientos EV1 y P + EV1 que fue en donde hubo nodulación. Observándose que ambos pesos resultaron ser mayores en el tratamiento EV1 que en el P + EV1, significando que el primer tratamiento presentó mayor biomasa producto de la mayor incorporación de materia orgánica como elementos estructurales, por lo tanto mayor tejido nodular que es capaz de hidratarse y registrar peso fresco mayor respecto al tratamiento P + EV1. 78 0,035 Peso promedio (g) 0,03 0,025 0,02 0,015 Peso fresco nódulos 0,01 Peso seco nódulos 0,005 0 Tratamientos Figura 21. Pesos fresco y seco promedio de nódulos por tratamiento en Vicia faba L.. Control (sin aislado y sin Previcur), ES1 (aislado ES1 sin Previcur), EV1 (aislado EV1 sin Previcur), P+EV1 (Previcur + EV1), P+ES1 (Previcur + ES1), Previcur (Previcur sin aislado). En las Tablas 29 y 30, se demuestra mediante el análisis estadístico del ANOVA simple que diferencias estadísticamente significativas entre ambos tratamientos vistas en el contraste múltiple de rango, y se puede corroborar que la nodulación se vé afectada negativamente por la aplicación de altas concentraciones de carbamatos, como sucedió con la variable número de nódulos. 79 Tabla 29. ANOVA simple para resultados de peso fresco de nódulos de V. faba L. con p < 0,05 y contraste múltiple de rango con un nivel de confianza del 95,0%, según los tratamientos. Análisis de la Varianza -----------------------------------------------------------------------------Fuente Sumas de cuad. Gl Cuadrado Medio Cociente-F P-Valor -----------------------------------------------------------------------------Entre grupos 0,00455522 5 0,000911044 11,17 0,0000 Intra grupos 0,00244667 30 0,0000815556 -----------------------------------------------------------------------------Total (Corr.) 0,00700189 35 Contraste Múltiple de Rango para -------------------------------------------------------------------------------Método: 95,0 porcentaje LSD Tratamientos Frec. Media Grupos homogéneos -------------------------------------------------------------------------------PREVICUR 6 0,0 c CONTROL 6 0,0 c ES1 6 0,0 c P+ES1 6 0,0 c P+EV1 6 0,0113333 b EV1 6 0,0303333 a -------------------------------------------------------------------------------- Tabla 30. ANOVA simple para resultados de peso seco de nódulos de V. faba L. con p < 0,05 y contraste múltiple de rango con un nivel de confianza del 95,0%, según los tratamientos. Análisis de la Varianza -----------------------------------------------------------------------------Fuente Sumas de cuad. Gl Cuadrado Medio Cociente-F P-Valor -----------------------------------------------------------------------------Entre grupos 0,000422806 5 0,0000845611 16,46 0,0000 Intra grupos 0,000154167 300,00000513889 -----------------------------------------------------------------------------Total (Corr.) 0,000576972 35 Contraste Múltiple de Rango -------------------------------------------------------------------------------Método: 95,0 porcentaje LSD Tratamientos Frec. Media Grupos homogéneos -------------------------------------------------------------------------------PREVICUR 6 0,0 c CONTROL 6 0,0 c ES1 6 0,0 c P+ES1 6 0,0 c P+EV1 6 0,00283333 b EV1 6 0,00933333 a -------------------------------------------------------------------------------- En la Figura 22 se observan las longitudes de vástago y raíz de acuerdo a cada tratamiento. La mayor longitud de vástago fue la del tratamiento P + ES1, sin embargo, es notorio en la Tabla 31, que no existen diferencias significativas entre todos los tratamientos. La longitud de los sistemas radicales en los diferentes tratamientos son prácticamente del mismo tamaño, sin diferencias significativas. Por 80 lo tanto, no puede sugerirse un efecto promotor de crecimiento de parte de la bacteria ni siquiera debido al fungicida (Tabla 32). Longitud promedio (cm) 60 50 40 30 Long Vástago 20 Long Raíz 10 0 Control ES1 EV1 P + EV1 P + ES1 Previcur Tratamientos Figura 22. Longitud promedio de vástago y raíz de plantas de Vicia faba L. Control (sin aislado y sin Previcur), ES1 (aislado ES1 sin Previcur), EV1 (aislado EV1 sin Previcur), P+EV1 (Previcur + EV1), P+ES1 (Previcur + ES1), Previcur (Previcur sin aislado). Tabla 31. ANOVA simple para longitud de vástago de V. faba L. con p < 0,05 y contraste múltiple de rango con un nivel de confianza del 95,0%, según los tratamientos. Análisis de la Varianza -----------------------------------------------------------------------------Fuente Sumas de cuad. Gl Cuadrado Medio Cociente-F P-Valor -----------------------------------------------------------------------------Entre grupos 329,733 5 65,9467 0,76 0,5831 Intra grupos 2590,52 30 86,3506 -----------------------------------------------------------------------------Total (Corr.) 2920,25 35 Contraste Múltiple de Rango -------------------------------------------------------------------------------Método: 95,0 porcentaje LSD Tratamientos Frec. Media Grupos homogéneos -------------------------------------------------------------------------------ES1 6 38,1667 X EV1 6 41,0 X PREVICUR 6 42,5 X P+EV1 6 42,5 X CONTROL 6 43,25 X P+ES1 6 48,2833 X -------------------------------------------------------------------------------- 81 Tabla 32. ANOVA simple para resultados de longitud de raíz de V. faba L. con p < 0,05 y contraste múltiple de rango con un nivel de confianza del 95,0%, según los tratamientos. Análisis de la Varianza -----------------------------------------------------------------------------Fuente Sumas de cuad. Gl Cuadrado Medio Cociente-F P-Valor -----------------------------------------------------------------------------Entre grupos 21,1014 5 4,22028 0,43 0,8240 Intra grupos 294,408 30 9,81361 -----------------------------------------------------------------------------Total (Corr.) 315,51 35 Contraste Múltiple de Rango -------------------------------------------------------------------------------Método: 95,0 porcentaje LSD Tratamientos Frec. Media Grupos homogéneos -------------------------------------------------------------------------------P+ES1 6 23,0833 X CONTROL 6 23,0833 X EV1 6 23,25 X P+EV1 6 23,25 X ES1 6 24,6333 X PREVICUR 6 24,9167 X -------------------------------------------------------------------------------- En la Figura 23 se muestran los pesos frescos y secos del vástago de las plantas de V. faba L. con diferentes tratamientos. El tratamiento Previcur, presentó el peso más alto, tanto fresco como seco, en comparación con los demás. Es importante destacar que las plantas de este tratamiento presentaron necrosis en la yema apical que se mantuvo durante todo el ensayo, posiblemente debida al efecto tóxico ocasionado por la concentración del carbamato usado, ya que era igual a la utilizada en campo pero en un área mucho más pequeña, lo que pudo haber ocasionado un exceso de nitrógeno (haciendo referencia al análisis de nitrógeno foliar de la Tabla 37) y haber producido la quema de este tejido vegetal. 82 8 Peso promedio (g) 7 6 5 4 3 Peso Fresco Vástago 2 Peso Seco Vástago 1 0 Tratamientos Figura 23. Pesos fresco y seco promedio de vástago de Vicia faba L.. Control (sin aislado y sin Previcur), ES1 (aislado ES1 sin Previcur), EV1 (aislado EV1 sin Previcur), P+EV1 (Previcur + EV1), P+ES1 (Previcur + ES1), Previcur (Previcur sin aislado). En la Tabla 33, el ANOVA mostró diferencias significativas entre los pesos fresco del vástago, siendo los grupos Previcur y P + ES1 iguales en su respuesta en el contraste múltiple de rango, pero muestran diferencias con el resto de tratamientos. El tratamiento P + EV1 es distinto de los dos anteriores así como de los demás, y los tratamientos Control, ES1 y EV1 iguales entre sí, y diferentes a los demás tratamientos. Según esto, la aplicación del carbamato favorecería la toma de agua por parte de las plantas, ya que son los valores más altos en cuanto al peso. 83 Tabla 33. ANOVA simple para resultados de peso fresco del vástago de V. faba L. con p < 0,05 y contraste múltiple de rango con un nivel de confianza del 95,0% según los tratamientos. Análisis de la Varianza -----------------------------------------------------------------------------Fuente Sumas de cuad. Gl Cuadrado Medio Cociente-F P-Valor -----------------------------------------------------------------------------Entre grupos 44,241 5 8,8482 4,19 0,0052 Intra grupos 63,3191 30 2,11064 -----------------------------------------------------------------------------Total (Corr.) 107,56 35 Contraste Múltiple de Rango -------------------------------------------------------------------------------Método: 95,0 porcentaje LSD Tratamientos Frec. Media Grupos homogéneos -------------------------------------------------------------------------------EV1 6 4,49833 c ES1 6 4,69333 c CONTROL 6 4,90667 c P+EV1 6 5,205 cb P+ES1 6 6,64333 ba PREVICUR 6 7,50167 a -------------------------------------------------------------------------------- En la Tabla 34 el ANOVA para el peso seco del vástago arrojó diferencias estadísticamente significativas entre los tratamientos, siendo en el contraste múltiple de rango los tratamientos Previcur, P + ES1 y Control iguales entre sí, presentando los valores más altos de peso seco, pudiéndose deber esto al riego que tuvieron 1 vez a la semana con la solución Hoagland a ¼ de la concentración de nitrógeno, lo que permitió la toma de este elemento por parte de las plantas y la traslocación al vástago, además de la aplicación del carbamato Previcur, ya que las plantas de Vicia faba L. acumulan este carbamato en la parte aérea (según el análisis de la tabla 37) específicamente a los tratamientos que Previcur y P + ES1. Por otra parte, los tratamientos EV1 y P + EV1 a pesar de haber sido los tratamientos que nodularon, hace inferir que esos nódulos fueron inefectivos ya que los pesos secos del vástago de este tratamiento fueron los más bajos, incluso similares al del tratamiento ES1 (tabla 34) que no presentó nodulación lo que indica que no hubo incorporación de nitrógeno a la planta. 84 Tabla 34. ANOVA simple para resultados de peso seco del vástago de V. faba L. con p < 0,05 y contraste múltiple de rango con un nivel de confianza del 95,0%, según los tratamientos. Análisis de la Varianza -----------------------------------------------------------------------------Fuente Sumas de cuad. Gl Cuadrado Medio Cociente-F P-Valor -----------------------------------------------------------------------------Entre grupos 0,854791 5 0,170958 3,47 0,0136 Intra grupos 1,47815 30 0,0492717 -----------------------------------------------------------------------------Total (Corr.) 2,33294 35 Contraste Múltiple de Rango -------------------------------------------------------------------------------Método: 95,0 porcentaje LSD Tratamientos Frec. Media Grupos homogéneos -------------------------------------------------------------------------------EV1 6 0,6845 c ES1 6 0,689167 c P+EV1 6 0,803 cb CONTROL 6 0,9 cba P+ES1 6 1,02 ba PREVICUR 6 1,08833 a -------------------------------------------------------------------------------- En la Figura 24 se observan los pesos frescos y secos de la raíz. El tratamiento P + EV1 presentó los valores más altos en comparación a los demás tratamientos, pero a pesar de esto no hay diferencias significativas entre los tratamientos respecto al control según el ANOVA para peso fresco de raíz (Tabla 35) y el ANOVA para el peso seco de raíz (Tabla 36). Sin embargo se observa en el contraste múltiple de rango para ambos pesos (Tabla 35 y 36) que el tratamiento ES1 tiende a ser diferente estadísticamente de los demás, y el tratamiento EV1 tiende a ser igual estadísticamente al ES1 en cuanto al peso seco, y diferente a los demás (Tabla 36). Con estos resultados se ratifica que la presencia del aislado rizobial EV1 en el sistema radical no es efectiva, y que tanto este aislado como el ES1, no pueden sugerirse tampoco como rizobacterias promotoras de crecimiento. 85 10 9 Peso promedio (g) 8 7 6 5 4 Peso Fresco Raíz 3 Peso Seco Raíz 2 1 0 Control ES1 EV1 P + EV1 P + ES1 Previcur Tratamientos Figura 24. Pesos fresco y seco promedio de raíz de Vicia faba L.. Control (sin aislado y sin Previcur), ES1 (aislado ES1 sin Previcur), EV1 (aislado EV1 sin Previcur), P+EV1 (Previcur + EV1), P+ES1 (Previcur + ES1), Previcur (Previcur sin aislado). Tabla 35. ANOVA simple para resultados de peso fresco de la raíz de V. faba L. con p < 0,05 y contraste múltiple de rango con un nivel de confianza del 95,0%, según los tratamientos. Análisis de la Varianza -----------------------------------------------------------------------------Fuente Sumas de cuad. Gl Cuadrado Medio Cociente-F P-Valor -----------------------------------------------------------------------------Entre grupos 74,197 5 14,8394 1,66 0,1753 Intra grupos 268,525 30 8,95084 -----------------------------------------------------------------------------Total (Corr.) 342,722 35 Contraste Múltiple de Rango -------------------------------------------------------------------------------Método: 95,0 porcentaje LSD Tratamientos Frec. Media Grupos homogéneos -------------------------------------------------------------------------------ES1 6 5,00333 b EV1 6 5,975 ba CONTROL 6 7,62 ba P+ES1 6 8,24833 ba PREVICUR 6 8,76167 a P+EV1 6 8,80333 a -------------------------------------------------------------------------------- 86 Tabla 36. ANOVA simple para resultados de peso seco de la raíz de V. faba L. con p < 0,05 y contraste múltiple de rango con un nivel de confianza del 95,0%, según los tratamientos. Análisis de la Varianza -----------------------------------------------------------------------------Fuente Sumas de cuad. Gl Cuadrado Medio Cociente-F P-Valor -----------------------------------------------------------------------------Entre grupos 0,254114 5 0,0508228 1,75 0,1526 Intra grupos 0,868799 30 0,02896 -----------------------------------------------------------------------------Total (Corr.) 1,12291 35 Contraste Múltiple de Rango ------------------------------------------------------------------------------Método: 95,0 porcentaje LSD Tratamientos Frec. Media Grupos homogéneos ------------------------------------------------------------------------------ES1 6 0,474333 b EV1 6 0,484 b CONTROL 6 0,602667 ba P+ES1 6 0,616167 ba PREVICUR 6 0,65 ba P+EV1 6 0,705333 a ------------------------------------------------------------------------------- En la Figura 25 se muestra el contenido de nitrógeno de la parte aérea de Vicia faba L. para cada tratamiento. Como se observa, el tratamiento con Previcur fue el que aportó mayor cantidad de nitrógeno a las plantas, posiblemente debido a la aplicación de este fungicida, lo que implica una fuente de nitrógeno artificial, aunado al riego que se hizo con solución Hoagland a ¼ de la concentración de nitrógeno, por tanto, este tratamiento tuvo doble fuente de nitrógeno y por ello la diferencia en las cantidades asimiladas con respecto a los otros tratamientos. 87 Porcentaje de Nitrógeno foliar (%) 3 2,5 2 1,5 1 0,5 0 Control ES1 EV1 P + EV1 P + ES1 Previcur Tratamientos Figura 25. Análisis del contenido de nitrógeno de la parte aérea de Vicia faba L. (%). Control (sin aislado y sin Previcur), ES1 (aislado ES1 sin Previcur), EV1 (aislado EV1 sin Previcur), P+EV1 (Previcur + EV1), P+ES1 (Previcur + ES1), Previcur (Previcur sin aislado). En la Tabla 37 se muestran los resultados del ANOVA simple para el análisis de nitrógeno de la parte aérea de Vicia faba L. según los tratamientos, el cual no arroja diferencias significativas entre los tratamientos. En el contraste múltiple de rango no se observan diferencias significativas entre los tratamientos que contienen ambos aislados como lo son ES1, P + ES1, EV1 y P + EV1, ni entre ellos con respecto al control, por lo que el aislado EV1 quien fue el que noduló a Vicia faba L. no es una bacteria efectiva en el aporte de nitrógeno debido a que no realizó la fijación biológica de N. Sin embargo, el tratamiento Previcur se muestra como estadísticamente diferente con respecto a los tratamientos Control, ES1 y P + ES1. La fitorremediación es una reciente técnica efectiva, estéticamente agradable y de bajo costo tecnológico que consiste en utilizar plantas para descontaminar los suelos o el agua, en donde se utiliza la habilidad de las plantas para concentrar elementos y compuestos del medio ambiente y metabolizar varias moléculas en sus tejidos. Las plantas pueden transportar y concentrar metales del suelo, inclusive compuestos orgánicos (Garbisu, et al., 2002). 88 En este sentido, Garcinuño et al., 2003, evaluaron el efecto de los pesticidas Carbaryl, Fenamiphos y Permethrin en semillas de Lupinus angustifolius, mostrando que éstas, luego de 17 días, tienen una alta capacidad de retención de los pesticidas mencionados y para extraerlos de sus semillas utilizaron solventes orgánicos como methanol, acetonitrilo, ethyl acetato y n-hexano, y no detectaron residuos de estos pesticidas, de manera que atribuyeron la capacidad de estas semillas para degradar los pesticidas utilizados. De igual manera Garcinuño et al. (2006) observaron por cromatografía líquida que plantas de Lupinus angustifolius bajo condiciones hidropónicas luego de 16 días de crecimiento, lograron degradar el 57, 53 y 55% de los pesticidas Carbaryl, Linuron y Permetrhrin respectivamente, sugiriendo que L. angustifolius podría ser usada como remediadora de aguas contaminadas por pesticidas. En el caso de Vicia faba L., se puede sugerir que esta especie podría actuar probablemente como una planta fitoacumuladora o fitodestoxificante en base a los resultados del ANOVA simple del contenido de nitrógeno de la parte aérea (Tabla 37), en donde las plantas de V. faba L. con el tratamiento Previcur (Previcur sin aislado) resultaron diferentes estadísticamente a las del control, lo que indica que la planta está tomando el Previcur, y hace inferir que esta especie podría actuar como una planta fitoacumuladora de este fungicida. 89 Tabla 37. ANOVA simple para resultados de análisis de contenido de nitrógeno de la parte aérea de V. faba L. con p < 0,05 y contraste múltiple de rango con un nivel de confianza del 95,0%, según los tratamientos. -----------------------------------------------------------------------Fuente Sumas de cuad. Gl Cuadrado Medio Cociente-F P-Valor -----------------------------------------------------------------------------Entre grupos 1,01395 5 0,202789 2,23 0,0772 Intra grupos 2,72842 30 0,0909472 -----------------------------------------------------------------------------Total (Corr.) 3,74236 35 Contraste Múltiple de Rango -----------------------------------------------------------------------------Método: 95,0 porcentaje LSD Tratamiento Frec. Media Grupos homogéneos -----------------------------------------------------------------------------ES1 6 2,00833 b P+ES1 6 2,12 b CONTROL 6 2,145 b P+EV1 6 2,245 ba EV1 6 2,26833 ba PREVICUR 6 2,545 a ------------------------------------------------------------------------------ Los resultados de este trabajo sugieren que el aislado nodulante EV1 es infectivo más no es efectivo para la especie Vicia faba L. debido a que no es una bacteria potencialmente fijadora de nitrógeno, y que el aislado ES1 no es infectivo para esta especie de planta y debe ser probado en otra especie de leguminosa para determinar su infectividad y efectividad en la fijación biológica de nitrógeno. En cuanto al efecto sobre la planta, los resultados indican que la presencia del carbamato Previcur en su concentración original (aplicada en campo), incrementó significativamente el contenido de nitrógeno en la parte aérea en relación al tratamiento control, indicando una fitoacumulación de este carbamato en esta parte de la planta. 90 CONCLUSIONES 1. Se lograron capturar, aislar, purificar y autenticar bacterias del suelo tratado con carbamatos y del suelo sin tratamiento agroquímico, y fueron designadas como ES1 y EV1 respectivamente. 2. ES1 y EV1 posiblemente sean del género Rhizobium sp. 3. Los rizobios mostraron ser más resistentes a los pesticidas Carbodan y Previcur en la dilución ½, y fueron sensibles a Dithane, Vondozeb y Zineb. 4. Los aislados no metabolizaron los carbamatos Carbodan, Previcur y Zineb, es decir, no los utilizaron ni como fuente carbonada ni como fuente nitrogenada. 5. Existe una promoción del crecimiento bacteriano en presencia del carbamato Previcur o Carbodan y fuentes de carbono, sugiriendo un proceso de cometabolismo para ambas bacterias, que favorece a ES1 y es tóxico para EV1. 6. ES1 cometabolizó Previcur y Carbodan, en donde el producto biotransformado aceleró el metabolismo y promovió su crecimiento celular. 7. EV1 cometabolizó Previcur y Carbodan, originando un producto tóxico que acelera la disminución de su población bacteriana. 8. Se determinaron las DL50 para ambos aislados con los carbamatos Carbodan y Previcur, sugiriendo una mayor resistencia del aislado ES1 a dichos carbamatos, indicando que de alguna manera esta bacteria tiene la propiedad de ser más resistente a pesticidas por encontrarse en un suelo sometido a tratamientos agroquímicos, bajo una presión selectiva. 9. La germinación de semillas de Vicia faba L. resultó ser inhibida por los carbamatos Carbodan y Zineb en sus concentraciones originales (aplicadas en campo), mientras que no fue afectada por la concentración original del carbamato Previcur. 91 10. En la prueba de nodulación de Vicia faba L. el aislado ES1 resultó no ser infectivo, señalando que esta leguminosa es muy especifica como hospedador. El aislado EV1 fue infectivo en esta prueba, pero en condiciones de invernadero no fue potencialmente fijador de nitrógeno con Vicia faba L., ya que no hubo diferencias significativas entre los tratamientos en el análisis de nitrógeno foliar, ni se obtuvieron nódulos de coloración rojiza. 11. La nodulación fue afectada desfavorablemente ante la presencia de altas concentraciones del carbamato Previcur (concentración original aplicada en campo) debido a la disminución del número de nódulos del tratamiento EV1 al tratamiento P + EV1, ya que esta diferencia fue estadísticamente significativa. 12. Es importante destacar que el Previcur incrementó los niveles nitrogenados de la parte aérea de las plantas de las plantas de Vicia faba L., permitiendo sugerir a esta especie de leguminosa como una posible planta fitoacumuladora o fitodestoxificante para este carbamato. 92 RECOMENDACIONES • Se recomienda hacer un ensayo de nodulación y crecimiento en otra especie de leguminosa con el aislado ES1 para observar su infectividad y efectividad. • El aislado ES1 podría ser objeto de estudios posteriores en el tema de la biorremediación, ya que resultó ser una bacteria que cometaboliza el carbamato Previcur y Carbodan y el producto biotransformado puede ser utilizado por él mismo promoviendo su crecimiento, y posiblemente este producto pueda ser degradado por otros microorganismos en la naturaleza, y disminuir los niveles de estos carbamatos en el suelo. • Realizar estudios utilizando Vicia faba L., como una posible planta fitoacumuladora o fitodestoxificante del carbamato Previcur, en condiciones de campo. 93 REFERENCIAS BIBLIOGRÁFICAS Aamil, M., Zaidi, A., Saghir, Md. 2004. Fungicidal impact on chickpea-Mesorhizobium symbiosis. Journal of Environmental Science and Health. 2004; 39(5-6): 779-790. Aguilar, O. M., López, M. V. and Riccilio, P. M. 2001. The Diversity of Rhizobia Nodulating Beans in Northwest Argentina as a Source of More Efficient Inoculant Strain. Biotechnol. 91: 181 – 188. Allen, O.N y Allen, E. 1981. The Leguminosae. A Source Book of Characteristics, Uses and Nodulation. Macmillan Publishers LTD. United states of America. p.p 677, 678, 680. Alexander, M. 1977. Introduction to Soil Microbiology. Second edition. John Wiley and Sons. New York. p.p 442, 447. Alexander, M. 1999. Biodegradation and bioremediation. Academic Press. U.S.A. Amadeo, C. 2006. Disponible: Inoculación. La fijación del nitrógeno por simbiosis. http://www.elsitioagricola.com/articulos/amadeo/Inoculacion%20- %20la%20Fijacion%20del%20Nitrogeno%20por%20Simbiosis.asp. Visitada en Marzo de 2009. Beringer, J. E. 1974. R. Factor Transfer in Rhizobium leguminosarum. J. Gen. Microbiol. 84: 188 - 198. Bromfield, E. S. y Barran, L. R. 1990. Promiscuous Nodulation of Phaseolus vulgaris, Macroptilium atropurpureum and Leucaena leucocephala by Indigenous Rhizobium meliloti. Can. J. Microbiol, 36: 369 – 372. 94 Caetano – Anollés, G., Lagares, A., Favelukes, G. 1989. Adsorption of Rhizobium meliloti to alfalfa roots: Dependence on divalent cations and pH. Plant and Soil 117: 67 – 74. Castro, D. 2008. Caracterización de cepas bacterianas resistentes al dicloro difenil tricloroetano (DDT) y Paration aisladas de suelos sometidos a manejo agrícola intensivo de diferentes localidades del estado Mérida. Trabajo especial de grado para optar al título de Licenciado en Biología. Facultad de Ciencias. Universidad de Los Andes, Venezuela. p.p 12, 20, 39. Chapalamadugu, S., Chaudhry, GR. 1991. Hydrolisis of carbaryl by a Pseudomonas sp. and construction of a microbial consortium that completely metabolizes carbaryl. Applied and Environmental Microbiology. Mar. 1991; 57(3):744-50. CIAT (Centro Internacional de Agricultura Tropical). 1988. Simbiosis LeguminosaRizobio. Manual de Métodos de Evaluación, Selección y Manejo Agronómico. Centro Internacional de Agricultura Tropical. Cali. Colombia. p.p. 3-1, 3-2. Criddle, C. S. 1993. The kinetics of cometabolism. Biotechnology and Bioengineering. 41: 1048 - 1056 Cronquist, A. 1981. An Integrated System of Classification of flowering Plants. Columbia University Press. New York. United States of America. p.p xiii, xv. Díaz, C. y De la Sovera, V. 2009. Carbamatos. Disponible en http://mail.fq.edu.uy/~planta/Cursos/neonicarbam.pdf. Visitada en Febrero de 2009. 95 Dich, J.: Zahm, S.: Hanberg, A.: Adami, H. O. (1997). Pesticides and cancer. Cancer Causes Control 8: 420-443. Esser-Mönning, K., Roskothen, P., Röbbelen, G. 1995. Two host genes in Vicia faba for nodulation deficiency with strain specificity for Rhizobium leguminosarum. Plant Breeding. 1995; 114 (4): 363-365. Evans, L. S., Lewin, K. F., Vella, F. A. 1980. Effect on nutrient medium pH on symbiotic nitrogen fixation by Rhizobium leguminosarum and Pisum sativum. Plant soil 56: 71 – 80. Feng, X., Ou, L. y Ogram, A. 1997. Plasmid-Mediated Mineralization of Carbofuran by Sphingomonas sp. Strain CF06. Applied and Environmental Microbiology. Apr. 1997; 63 (4): 1332–1337. Ferrer, A. (2003). Intoxicación por Plaguicidas; Pesticida Poisoning. Anales del Sistema Sanitario de Navarra 26: 155-171. Ferrera, R., González, M., Rodríguez, M. 1993. Manual de Agromicrobiología. Editorial Trillas. México. p.p. 45-46. Garbisu, C., Hernández-Allica, J., Barrutia, O., Alkorta, I., Becerril, J. M. 2002. Phytoremediation: a technology using green plants to remove contaminants from polluted areas. Rev Environ Health. Jul-Sep; 17 (3): 173 - 88. García-Rivero, M. y Peralta-Pérez, M. R. 2008. Cometabolismo en la Biodegradación de Hidrocarburos. Revista Mexicana de Ingeniería Química. Abr 2008; 7(1): 1-12. Garcinuño, R. M., Fernández-Hernando, P., Cámara, C. 2003. Water Res. Aug; 37 (14): 3481 – 9. 96 Garcinuño, R. M., Fernández-Hernando, P., Cámara, C. 2006. Removal of carbaryl, linuron, and permethrin bye Lupinus angustifolius under hydroponic conditions. J Agric Food Chem. Jul 12; 54 (14): 5034 – 9. Gil Martínez, F. 1994. Fijación de Nitrógeno. En: Elementos de Fisiología vegetal. Ediciones Mundi – Prensa, Madrid. p.p 945. Gilabert, J., López, I., Pérez, R. 1990. Análisis de Suelos para Diagnóstico de Fertilidad. Manual de Métodos y Procedimientos de Referencia. MAC, FONAIAP, CNIA, Escuela de Agronomía – UCLA. Maracay, Venezuela. ISBN 980 – 6090 – 89 – 6. Glenn, A. R. y Dilworth, M. J. 1994. The life of root nodule bacteria in the acidic underground. FEMS Microbiol. Lett 123: 1 – 10. Graham, P. H., Draeger, K. J., Ferrey, M. L., Conroy, M. J., Hammer, B. E., Martínez, E., Aarons, S. R., Quinto, C. 1994. Acid pH tolerance in strains of Rhizobium and Bradhyrhizobium, and initial studies on the basis for acid tolerance of Rhizobium tropici UMR 1899. Can. J. Microbiol 40: 198 – 207. Hashimoto, M., Fukui, M., Hayano, K., Hayatsu, M. 2001. Nucleotide sequence and genetic structure of a novel carbaryl hydrolase gene (cehA) from Rhizobium sp. strain AC100. Applied and Environmental Microbiology. Mar.2002; 68(3): 12201227. Hayatsu, M., Hirano, M., Nagata, T. 1999. Involvenment of two plasmids in the degradation of carbaryl by Arthrobacter sp. strain RC100. Applied and Environmental Microbiology. Mar.1999; 65(3): 1015-9. Hotchkiss, J. (1992). Pesticide Residue Controls to Ensure Food Safety. Critical Rev. Food Sci. Nutrition 31:191-203. 97 Jordan, D. C. 1984. Rhizobiaceae. In Bergey’s Manual of Systematic Bacteriology. Vol I. Edit N. R. Krieg.. Williams and Wilkings, Baltimore, Usa. P.p 234-256. Indice Agropecuario. 2001. Disponible en http://danac.org.ve/indice/detalle_productos.php?ps=478554. Visitada en Enero de 2009. Janke, D. y Fristche. W. 1985. Nature and significance of microbial cometabolism of xenobiotics. Journal of Basic Microbiology Lal, R y Saxena, D. (1982). Accumulation, Metabolism and Effects of Organochlorine Insecticides on Microorganisms. Microbiological Reviews 46 (1): 95- 127. Larkin, M. J., Day, M. J. 1984. The effect of pH on the selection of carbaryl-degrading bacteria from garden soil. Applied Bacteriology. 1985: 58: 175-185. Larkin, M.J., Day, M. J. 1985. The metabolismo f carbaryl by three bacterial isolates, Pseudomonas spp. (NCIB 12042 & 12043) and Rhodococcus sp. (NCIB 12038) from garden soil. Applied Bacteriology. Mar. 1986; 60(3): 233-42. Leadbetter, E.R. y Foster, J. W. 1959. Oxidation products formed from gaseus alkanes by the bacterium Pseudomonas methanica. Archives of Biochemistry and Biophysics 82: 491 – 492. Lodeiro, A.R., López García, S., Mongiardini, E., Quelas, J. I. y Perticari, A. 2003. Los Rizobios y la Inoculación de las Leguminosas para la Fijación Simbiótica de Nitrógeno. En: Microbiología Agrícola. Un aporte de la investigación argentina. Albanesi, A., Anriquez, A., Luna, S., Kunst, C., Ledesma, R. (eds.). Universidad Nacional de Santiago del Estero. ISBN: 987-99083-5-X. pp. 159-193. 98 Longnecker, MP., Rogan, WJ., Lucier, G. (1997). The human effects of DDT (dichloro diphenyl trichloroethane) and PCBs (polychlorinated bisphenyls) and an overview of organochlorines un public health. Annu Rev Public Health 18: 211-244. Luo, W., Zhao, Y., Ding, H., Lin, X., Zheng, H. 2008. Co-metabolic degradation of bensulfuron-methyl in laboratory conditions. Journal of Hazardous Materials 158 (2008) 208–214. Macbride, J. F. 1943. Flora of Peru. Botanical Series Field Museum of Natural History, Volume XIII, Part III, Number 1. United States of America. October, 1943 publicación 531. p.p 356. Machado, D. 2005. Un Enfoque Agroecosistémico para el Manejo Eficiente de Nitrógeno en el Cultivo de Papa en los Andes Venezolanos. Tesis Doctoral. Universidad de Los Andes, Instituto de Ciencias Ambientales y Ecológicas, Postgrado en Ecología Tropical. Mérida, Venezuela. 233p. Madigan, M. T., Martinko, J. M., Parker, J. 2000. Brock Biology of Microorganisms. (Bergey’s Manual of Systematic Bacteriology 2da Ed. A-8). Prentice Hall 9ed. United States of America. Malagón, D. 1982. Caracterización de suelos (Interpretación integral en función de su evolución). Serie Suelo y Clima SC – 55 CIDIAT – Mérida. Mápula-Larraeta, M., López-Upton, J., Vargas-Hernández, J. y Hernández-Livera, A. 2008. Germinación y Vigor de semillas de Pseudotsuga menziesii de México. Ra Ximhai Vol. 4. Número 1, enero.abril 2008, p.p 123. Maroni, M., Fait, A. (1993). Health Effects in Men from Long-term Exposure to Pesticides. Toxicology 78:1-180. 99 Marquina, M. 2006. Caracterización Fenotípica y Genotípica de Rizobios Autóctonos, Provenientes de Diversas Regiones de Venezuela. Tesis de Maestría para optar al Título de Magister Scientiae en Biotecnología de Microorganismos. Departamento de Biología, Facultad de Ciencias, Universidad de Los Andes. Mérida- Venezuela. p.p 1-2, 9-10, 62, 82, 130. Martinez-Romero, E. 2003. Diversity of Rhizobium Taxonomy. Plant and Soil 161: 11-20. Matos – Cuzcano, G., Ormeño – Orillo, E., Zúñiga – Dávila, D. 2004. Diversidad de los rizobios que nodulan el cultivo de Pallar (Phaseolus lunatus L.) En la Costa Central del Perú. Disponible en http://www.lamolina.edu.pe/ciencias/ecologia/revistas5.htm. Michiels, J., Dombrecht, B., Vermeiren, N., Xi, Ch., Luyten, E. y Vanderleyden, J. 1998. Phaseolus vulgaris is non-selective host for nodulación. Microbiology Ecology 26: 193-205. Miller, J.H. 1972. Experiments in molecular genetics. Cold Spring Harbor Laboratory, Cold Spring Harbor, N. Y. p.p 431-435. Miranda, I. (2006). “La estadística del agropecuario. 10 preguntas 10 respuestas”. Disponible: http://www.monografias.com/trabajos19/estadistica- agropecuario/estadistica-agropecuario.shtml (visitada en octubre de 2008). Munns, D. N. 1970. Nodulation of Medicago sativa in solution culture. V. Calcium and pH requirements during infection. Plant and Soil 32: 90 – 102. Mutch, L. and Young, J. P. 2004. Diversity and specificity of Rhizobium leguminosarum biovar viciae on wild and cultivated legumes. Molecular Ecology. 2004; 13: 2435-2444. 100 Nan, Z., Ge, G., Li, C. 2002. Effect of pesticides on field-controlling root rot of Vicia faba. Ying Yong Sheng Tai Xue Bao.. Aug; 13(8): 943-7. Noel, T. C., Sheng, C. K., Yost, R. P., Pharis and Hynes, M. F. 1996. Rhizobium leguminosarum as a Plant Growh – Promoting Rhizobacterium. Direct Growth Promotion of Canola and Lettuce. Can. J. Microbiol. 42: 279-282. Odee, D.W., Sutherland, E.T., McInroy, S.G. y Sprent, J.I. 1996. Phenotypic characteistics and composition of rhizobia associated with woody legumes growing in diverse Kenyan conditions. Plant and Soil 188: 65-75. Olea, N. y Fernández, M. (2001). Plaguicidas persistentes. Congreso Implementación del Convenio de Contaminantes Orgánicos Persistentes. Madrid, 26-27 de noviembre. Pacovsky, interaction R.S., Bayne, between H.G strain of and Bethlenfalvay, Rhizobium G.J. phaseoli 1984. and Symbiotic cultivars of Phaseolus vulgaris L. Crop. Sci 24: 105-107. Perry, J.J. 1979. Microbial cooxidations involving hydrocarbons. Microbiological Reviews, Mar. 1979; 43(1): 59 – 72. Primo, E. y Carrasco, J.M. 1980. Química Agrícola II Plaguicidas y Fitorreguladores. Editorial Alhambra, S.A. Madrid – España. p.p 1, 224 - 225, 228 – 229, 400 – 401. Ricardi, M. 1992. Familias de Dicotiledóneas venezolanas II. Universidad de Los Andes. Departamento de Biología. Facultad de Ciencias. Mérida, Venezuela. p.p 26. 101 Rodríguez - Mendoza M.N. y Ferrera – Cerrato, R. 1984. Estudio y Caracterización de la Relación Simbiótica de Rhizobium phaseoli frijol Phaseolus vulgaris de Distintos Hábitos de Crecimiento. Memorias XII Reunión Latino Americana sobre Rhizobium, Campinas, Sao Paulo - Brasil. p.p 22. Sánchez, J., Rodríguez, Biorremedación. J. Fundamentos Universidad de y aspectos Oviedo. Microbiológicos, Disponible en http://ingenierosdeminas.org/docu/documentos/fundamentos_%20biorremediacion.pd f Visitada en Febrero de 2009. Skwierinski, R. y Marquina, M. 2006. Captura y nodulación de bacterias rizobiales a partir de muestras de suelos. Prácticas de Simbiosis. Curso de Simbiosis. Departamento de Biología. Facultad de Ciencias. Universidad de Los Andes. Mérida. Venezuela. Smit, G., Tubbing, D. M. J., Jijne, J. W., Lugtenberg, B. J. J. 1991. Role of Ca2+ in the activity of ricadhesin from Rhizobium leguminosarum biovar viciae, which mediates the first step in attachment of Rhizobiaceae cells to plant root hair tips. Arch. Microbiol. 155: 278 – 283. Smith, R. J. y Gallon, J. R. 1993. Nitrogen Fixation. En: Plant Biochemistry and Molecular Biology. Edited by P- J. Lea and R. C. Leegood. John Wiley & Sons Ltd. p.p 131. Taiz , L., Zeiger, E. 2002. Plant Physiology. 3ed. Ed. Sunderland (USA): Sinauer Associates Inc. p.p 71. Vandana P., S., y Prashant S., P. 2005. Metabolism of Carbaryl via 1,2- dihydroxynaphthalene by Soil Isolates Pseudomonas sp. Strain C4, C5 and C6. Applied and Environmental Microbiology, Oct. 2005; 71(10): 5951-5956. 102 Vincent, JM. 1975. Manual Práctico de Rizobiología. Editorial Hemisferio Sur. Buenos Aires. Argentina. p.p. 4. Wang, T., Romero-Martínez, J., López-Lara, I. 2001. Rhizobium y su Destacada Simbiosis con Plantas. En Microbios en Línea (Editrs. Martínez-Romero, E. y Martínez-Romero, J). cap 8. Disponible en: (http://biblioweb.dgsca.unam.mx/libros/microbios/index.html). Werner, D. 1992. Symbiosis of Plants and Microbes. Chapman & Hall. Great Britain. p.p 82. Young, J. M. 2003. The Genus Ensifer Casida 1982. Take Priority Over Sinorhizobium Chen et a. 1988, and Sinorhizobium morelense Wang et al., 2002 is a Later Synonym of Ensifer adhaerens (Casida 1982). Is the Combination ‘Sinorhizobium adhaerens’ (Casida 1982) Willems et al. 2003 Legitimate? Request For an Opinion. Int. J. Sust. Evolu. Microbiol. 53: 2107-2110. 103 ANEXOS a) b) c) Anexo 1. Parcelas de donde fueron tomadas las muestras de suelo. a) Parcela cultivada con Solanum tuberosum L. (papa) tratada con agroquímicos; b) Parcela cultivada con Allium sativum L. (ajo) tratada con agroquímicos; c) Parcela natural o no cultivada sin tratamiento agroquímico. Anexo 2. Parcela cultivada con Vicia faba L. (haba) sin tratamiento agroquímico y que fue tomada como control. 104 a) b) Anexo 3. Captura de rizobios con cultivos trampa. a) Plantas de Phaseolus vulgaris L. (caraota); b) Plantas de Macroptilium lathyroides L. (siratro). a) b) Anexo 4. Nódulos de donde fueron aislados los rizobios. a) Nódulos formados en plantas de Phaseolus vulgaris L.; b) Nódulos de las plantas de Vicia faba L. 105 a) b) Anexo 5. Bacterias rizobiales aisladas creciendo en medio YMAmrc. a) Aislado ES1 proveniente del suelo cultivado con Solanum tuberosum L. tratado con carbamatos; b) Aislado EV1 proveniente del suelo cultivado con Vicia faba L. no tratado con carbamatos (control). a) b) c) Anexo 6. Prueba cualitativa de discos de papel impregnados con las distintas concentraciones de los carbamatos para observar la viabilidad de los rizobios en presencia de estos compuestos. a) Aislado ES1 en YMA + Carbodan ½ (hubo crecimiento); b) Aislado ES1 en YMA + Vondozeb 1/6 (mostrando halo de inhibición); c) Aislado EV1 en YMA + Vondozeb 1/6 (no hubo crecimiento). 106 Anexo 7. Trabajo experimental en el laboratorio realizando las curvas de crecimiento de rizobios en presencia de carbamatos. Anexo 8. Prueba de germinación de semillas de Vicia faba L. en presencia de carbamatos. a) b) c) d) Anexo 9. Preparación de la prueba de nodulación y crecimiento de Vicia faba L.; a) Lavado de la arena nueva; b) Secado de la arena en estufa a 120ºC; c) Llenado y preparación de los potes; d) Esterilización de los potes 1,5 Psi (1Atm)x1h/3 días. 107 Anexo 10. Plantas de Vicia faba L. en el invernadero en la prueba de nodulación y crecimiento de esta especie. ES Control Anexo 11. Plantas de Vicia faba L. nodulada (ES1) y no nodulada (control) en el desmontaje de la prueba de nodulación y crecimiento de esta especie. 108 a) b) c) Anexo 12. Medición de algunas de las variables en la prueba de nodulación y crecimiento de Vicia faba L. a) Longitud de raíz (cm); b) Peso fresco del vástago (g); c) Peso fresco de nódulos (g). Anexo 13. Preparación de las muestras para el análisis de nitrógeno foliar. 109