Cap 2
Anuncio

&$3Ë78/2)81'$0(1726'(/$5('1(8521$/&0$&
,QWURGXFFLyQ
Las teorías de Albus y Marr se
desarrollan sobre la base de los avances en la
fisiología y funcionamiento del circuito
neuronal del cerebelo, entre éstos los trabajos
de Ito y Eccles [Ecc67] en la identificación de
las interconexiones funcionales en los
componentes del cortex cerebelar.
El conocimiento sobre el tipo y
fisiología de las células cerebelosas y su
sinápsis fue modelizada matemáticamente por
Albus [Alb81] . El objetivo de Albus era
desarrollar un sistema artificial capaz de emular
al cerebelo en el procesamiento de la
información característica de éste. Este sistema
artificial podría ser utilizado más tarde en el
control de sistemas articulados, como brazos
robots, ofreciendo una herramienta potente y
"!#%$ & (') * $
sencilla de utilizar.
+,(-+."/0 $1
234& $5& $) "067*3
El cerebelo recibe información desde
el cerebro, espina dorsal, sistema vestibular,
músculos, tendones, formación reticular y órganos sensores varios. A esta información se une una
entrada de realimentación inhibitoria, que reduce el número de células excitadas en la primera etapa de
sinápsis.
El cerebelo contribuye al control de los movimientos rápidos y precisos, control del desarrollo
temporal de movimientos balísticos rápidos, programa la duración de dichos movimientos rápidos, y
vigila y establece ajustes correctores de las actividades motoras desencadenadas por otras partes del
encéfalo. El mecanismo de realimentación provee de capacidades como la amortiguación de los
movimientos musculares, ayudando al sistema nervioso central en la predicción de la posición de las
partes móviles del cuerpo. Estas características son interesantes en el campo del control del movimiento
de robots, pudiendo servir como “cerebelo” artificial que controle la actividad motora en robots.
Un aspecto importante del cerebelo sería su aprendizaje, y capacidad de almacenamiento de
patrones dinámicos de entrada. El modelo CMAC “aprendería” los patrones de entrada según el modelo
de aprendizaje por condicionamiento instrumental. Este tipo de aprendizaje se basa en la asociación entre
una respuesta y un estímulo, permitiendo al organismo adaptar su conducta en función de las
consecuencias de la misma. La asociación se realiza por aproximación mediante estímulos reforzadores
(“respuesta correcta”) y punitivos (“repuesta no correcta”), reforzando y castigando los pesos de las
células neuronales que intervinieron en el procesamiento del estímulo.
El modelo Perceptron de Rosenblatt [Ros58] fue uno de los primeros en modelizar una neurona
básica, formulando una de las primeras redes neuronales. Se pensaba que estos trabajos influirían en
campos como la neurofisiología y la neuropsicología, ya que proporcionaban una herramienta de
simulación de los procesos neuronales, no obstante no fue así debido a que la complejidad de una neurona
real es mucho mayor que el modelo simple que proponía el Perceptron. Los experimentos de Eccles/Ito
durante los 60 resaltaron las semejanzas entre la estructura y funcionamiento del cerebelo y la de la Red
Neuronal Perceptron, a nivel no interno, sino simplemente funcional, sirviendo de base al modelo
neuropsicológico publicado por Albus en 1971 [Alb71] .
8
En los siguientes capítulos se profundiza en cómo la memoria almacena las experiencias, y como
estas influyen en nuestra conducta, la capacidad de las redes neuronales de modelizar el aprendizaje
neuronal, la Red Neuronal Perceptron, y el condicionamiento instrumental como forma de aprendizaje
básica por estímulo-respuesta. Se resumen también el control motor del cerebro, la estructura, fisiología,
y funcionamiento del circuito del cerebelo y corteza cerebelosa, y el control que realiza sobre la acción
motora.
$QDWRPtDGHODPHPRULD
El modo en que las experiencias son almacenadas por nuestro cerebro, y como éstas condicionan
nuestra conducta futura, está ligado con la naturaleza de su almacenamiento. La memoria no es algo
tangible, que se “ almacena” como una imagen en un ordenador y se “ evoca” según necesidad. Las
experiencias FDPELDQ HO PRGR HQ TXH SHUFLELPRV DFWXDPRV SHQVDPRV \ SODQLILFDPRV PRGLILFDQGR
ItVLFDPHQWH OD HVWUXFWXUD GHO VLVWHPD QHUYLRVR DOWHUDQGR ORV FLUFXLWRV QHXURQDOHV TXH SDUWLFLSDQ HQ OD
SHUFHSFLyQ HO SHQVDPLHQWR \ OD SODQLILFDFLyQ >&DUO@ . Una experiencia desagradable, perturbadora,
tenderá a ser evitada; en cambio, intentaremos repetir aquellas que nos dieron placer o satisfacción.
La principal función del cerebro HV PRYHU ORV P~VFXORV (para producir conductas útiles). La
función de la capacidad de aprendizaje es el desarrollo de conductas útiles adaptadas a un entorno
dinámico. El aprendizaje se pone de manifiesto de tres formas básicas: aprendizaje perceptivo,
aprendizaje estímulo-respuesta, y aprendizaje motor.
El aprendizaje perceptivo es la FDSDFLGDG GH DSUHQGHU D UHFRQRFHU HVWtPXORV YLVWRV FRQ
DQWHULRULGDG\GLVWLQJXLUORVGHRWURVVLPLODUHV. Su principal función es la identificación y categorización
de objetos y situaciones. Cada uno de nuestros sistemas sensoriales es capaz de tener aprendizaje
perceptivo, lográndose mediante cambios en la corteza de asociación sensorial asociada a cada tipo de
estímulo (visual, auditivo, olfativo...).
El aprendizaje estímulo-respuesta es la FDSDFLGDGGHDSUHQGHU D UHDOL]DU XQWLSRHVSHFtILFRGH
FRQGXFWDDQWHXQGHWHUPLQDGRHVWtPXOR>&DUO@. Ésta podría tratarse de un acto reflejo, o una secuencia
complicada de movimientos aprendidos con anterioridad. El aprendizaje estímulo-respuesta incluye dos
categorías principales de aprendizaje que han sido estudiadas por los psicólogos: condicionamiento
clásico y condicionamiento instrumental.
El aprendizaje motor, constituye de hecho una forma especial de aprendizaje estímulo-respuesta,
donde los cambios se dan en los sistemas motores. El aprendizaje PRWRU GLILHUH GH RWUDV IRUPDV
SULQFLSDOPHQWHHQHOJUDGRHQTXHVHDSUHQGHQQXHYDVFRQGXFWDVFXDQWRPiVQRYHGRVDVVHDQpVWDVPiV
FLUFXLWRVQHXURQDOHVGHOVLVWHPDPRWRUFHUHEUDOWHQGUiQTXHPRGLILFDUVH [Carl94] .
5 /0!+!(84 $ / 9:
49
;'%" <
=4')" '& >.
;'%" <
=5$& 2 ? $ '@$& )A
B'%"4 <
=523& !
CAPÍTULO 2. FUNDAMENTOS DE LA RED NEURONAL CMAC
9
&RQGLFLRQDPLHQWR,QVWUXPHQWDO
El psicólogo F.B. Skinner realizó durante la década de los 70 una labor fundamental, “ el análisis
de la conducta” , en términos de estímulos y respuestas [Carp96] [Carl94] . Considerando conducta como
“ la actividad observable del individuo” que consiste fundamentalmente en el movimiento, se trata de
establecer “ relaciones funcionales” , leyes que regulen la dependencia de elementos del comportamiento
con otros (principalmente los estímulos y las respuestas). El organismo se considera como “ una caja
negra” , del que sólo conocemos su LQSXW y su RXWSXW. La correlación funcional entre estímulos y
respuestas la denomina “ reflejo” . Aceptando el acto reflejo como “ instrumento” para la descripción de la
conducta (condicionamiento clásico), pero sólo como acto de comportamiento y no como concepto
neurológico.
El condicionamiento instrumental examina los
cambios en la conducta en función de los
resultados (es decir, la experiencia), centrando
pues su atención en el aprendizaje.
Si el condicionamiento clásico implica
respuestas automáticas, actos reflejos típicos de
la especie, el condicionamiento instrumental
implica conductas aprendidas. Mientras el
condicionamiento
clásico
involucra
la
asociación entre dos estímulos, el instrumental
requiere la DVRFLDFLyQ HQWUH XQD UHVSXHVWD \
XQHVWtPXOR, permitiendo al organismo adaptar
su conducta en función de las consecuencias de
la misma.
Es decir, cuando una conducta es seguida de
consecuencias
favorables
(HVWtPXORV
UHIRU]DQWHV), tiende a aparecer con más
frecuencia; cuando es seguida de consecuencias
desfavorables (HVWtPXORV SXQLWLYRV), tiende a
aparecer con menos frecuencia. Por ejemplo,
5 /0!4C9!DEF2 9 $ 2G' G 2H 4& una respuesta que permita a un sujeto encontrar
$& 234& 9 !I%
74
2
94> %')
4 .G J'% & AI& 3 / 7
4& 2H 9& %')$ & >!KE
L
& > L
comida será reforzada, y una respuesta que cause
$ $& 2
39@" 1 < 1& 4 3$ ;'$ $(M5!
dolor será castigada.
La mayor parte de la investigación fisiológica
sobre el condicionamiento instrumental ha estudiado el refuerzo y no el castigo. La recompensa o
refuerzos actúan estableciendo cierta conducta, mientras el castigo tiende a suprimirla. La conducta
castigada no desaparece, y además pueden presentarse reacciones perturbadoras. El refuerzo ocasiona
cambios en el sistema nervioso del un animal, que aumentan la probabilidad de que un determinado
estímulo provoque una determinada respuesta.
Por ejemplo, cuando se coloca una rata
hambrienta por primera vez en una jaula, no es
muy probable que apriete la palanca localizada
en una de sus paredes. Sin embargo, si la
aprieta y recibe un trozo de comida
inmediatamente después, la probabilidad de
que ejecute otra vez esa respuesta aumenta. El
refuerzo hace que la visión de la palanca
constituya el estímulo que provoca la respuesta
/0!N!#=20'@ 5 2H 4& L $& 29& 4 !
#59& 20'3@$
;')$@"954& 20')!
10
de apretarla. El proceso de refuerzo establece una conexión entre los circuitos neuronales involucrados
en la percepción (la visión de la palanca) y los involucrados en el movimiento (el acto de apretar la
palanca).
Podemos ver otro ejemplo en el tiempo que necesita un gato para salir de una caja preparada. La
visión de la palanca y el acto de apretarla se ven reforzados por la acción de poder salir de la caja. La
probabilidad de ejecutar la respuesta correcta aumenta, reduciéndose es el tiempo para escapar con el
número de intentos.
¿Cómo podría funcionar el condicionamiento instrumental? La figura 3, muestra un posible modelo. El
sistema visual de la rata contiene dos neuronas. Una descarga cuando la rata ve una palanca y la otra
cuando ve una botella de agua. El sistema motor tiene tres neuronas, cada una con una acción específica
(mover orejas, sostenerse con las patas traseras y apretar la
palanca).
Otra neurona forma parte del sistema de refuerzo. Esta última
se activa cuando la rata tiene hambre y recibe algo de comida.
Inicialmente, la sinapsis P (de “ palanca” ) es demasiado débil
para hacer que la moto-neurona se active.
Sin embargo, la rata va explorando el entorno y, por
casualidad, acaba apretando la palanca. La respuesta es la
obtención de comida a la rata, que rápidamente se la come.
Este “ hecho positivo” , es detectado por el sistema de refuerzo,
que sinapta con todas las moto-neuronas. Cuando la sinapsis
R (de “ refuerzo” ) está activa, fortalece todas las sinapsis que
acaban de activarse. En este caso, es la sinapsis P de la
neurona la que hace que el hecho de apretar la palanca se vea
fortalecido.
Cuando esta última sinápsis ha sido
/0!O!#=20'@ 5 2H 4& H $& 2& 4
suficientemente fortalecida, la visión de la palanca
9$"
: (')%PQ !
provocará la conducta de apretar la palanca.
/DPRGHOL]DFLyQGHODFDSDFLGDGGHDSUHQGL]DMHGHO
FHUHEUR5HGHV1HXURQDOHV
La experiencia cambia las conexiones neuronales de nuestro cerebro, y estos cambios
representan lo que se ha aprendido. Pero ¿por qué un nuevo estímulo sensorial altera el modo en que las
neuronas responden a los estímulos ya aprendidos? Una posible respuesta se puede deducir a partir de los
modelos de funcionamiento de los circuitos neuronales, las denominadas UHGHV QHXURQDOHV A cada
elemento se le otorgan propiedades similares a las de las neuronas, y se les conecta entre sí por medio de
uniones similares a las sinapsis. Como las sinapsis, estas uniones pueden ejercer efectos excitatorios o
inhibitorios.
Cuando uno de los elementos recibe una cantidad crítica de excitación, envía un mensaje a
aquellos otros con los que está conectado, etc. Algunos de los elementos de una red tienen líneas de
entrada que pueden recibir señales del “ exterior” , lo cual podría representar un órgano sensorial o la
información recibida desde otra red, actuando como realimentación de la red. Otros elementos tienen
líneas de salida, que comunican con otras redes o controlan músculos, produciendo conductas. Por tanto,
los patrones particulares de entradas pueden representar estímulos determinados y los patrones
particulares de salidas pueden representar respuestas.
Es posible enseñar a las redes neuronales a reconocer estímulos concretos. Las redes reciben
sólo un patrón de entrada, siendo los elementos inhibitorios y reforzadores del circuito los que refinan la
respuesta de la red ante una determinada entrada. La primera vez que se presenta un estímulo
determinado, los elementos de salida responden de manera débil e inespecífica; pero después de varias
CAPÍTULO 2. FUNDAMENTOS DE LA RED NEURONAL CMAC
11
presentaciones, surge un patrón de salida fuerte y fiable. Pueden mostrase cada vez más estímulos a la
red, y generarse así patrones de salida únicos para cada uno de ellos.Las características funcionales de las
redes neuronales son similares a algunas de las exhibidas por los sistemas nerviosos reales, y es eso lo que
hace que resulten fascinantes para los científicos interesados en las bases neuronales del aprendizaje. Por
ejemplo, las redes neuronales muestran generalización, discriminación y degradación suave.
La JHQHUDOL]DFLyQ en este contexto debe entenderse como la capacidad para reconocer similitudes entre
estímulos.
Por ejemplo, supongamos que una red ha aprendido a reconocer unos cuantos estímulos
diferentes. Si luego le mostráramos un estímulo parecido a alguno de los conocidos, su patrón de
respuesta se parecerá mucho al patrón que produce cuando se le muestra el estímulo ya conocido. La
GLVFULPLQDFLyQse refiere a la capacidad para reconocer diferencias entre estímulos. Si mostramos a una
red varios estímulos similares, aprenderá a distinguirlos, y producirá patrones de salida muy diferentes
ante cada uno de ellos. Finalmente, la GHJUDGDFLyQ VXDYH hace referencia al hecho de que la lesión
aleatoria de los elementos de una red o de sus conexiones (“ sinapsis” ) no hace que la red deje de
funcionar totalmente. En lugar de ello, su ejecución se deteriora, siendo el nivel de este deterioro
proporcional a la magnitud de la lesión. Estos tres fenómenos, generalización, discriminación y
degradación suave, caracterizan también el funcionamiento de nuestro cerebro.
Como el cerebro está formado por redes neuronales, el aprendizaje tiene lugar en redes
neuronales. La cuestión es si las redes neuronales funcionan del mismo modo que el cerebro, y el único
modo de responder a esta cuestión es conociendo la anatomía detallada de los circuitos neuronales reales
y las características fisiológicas de las neuronas individuales y de sus conexiones sinápticas.
Cuanto más aprendamos sobre estos temas, más realistas serán los modelos de redes neuronales que
podrán construirse.
Los investigadores que estudian las propiedades de las redes neuronales hacen hincapié en que
están estudiando la PLFURHVWUXFWXUD del cerebro (las funciones llevadas a cabo por módulos individuales).
El cerebro contiene un elevado número de redes (probablemente, muchos millares), cada una de las cuales
está destinada a realizar funciones individuales. Es probable que esas redes existan en una especie de
jerarquía, en la que algunas controlen las funciones de otras y regulen el intercambio de información entre
12
ellas. Por consiguiente, la comprensión de las operaciones de redes neuronales individuales nunca
revelará todo lo que necesitamos saber acerca de las funciones del cerebro. También necesitaremos
conocer la organización del cerebro (las relaciones entre las redes individuales que lo forman).
Necesitamos conocer la PDFURHVWUXFWXUD cerebral. Así pues el desarrollo y perfeccionamiento de las
redes neuronales está ligado a la investigación sobre micro y macroestructura cerebral, que proporcione
una base fisiológica sobre la que asentar el desarrollo de las redes neuronales artificiales.
[Carl94]
/D5HG1HXURQDO3HUFHSWURQ
Desarrollada por el neurobiólogo Frank, en 1958, la red Perceptron fue el primer detector de
patrones regulares implementado intentando imitar las conexiones neuronales del cerebro. Rosenblatt
estaba intrigado con el funcionamiento del ojo de la mosca, el cual realiza la mayoría del proceso que
induce a la mosca a volar. El Perceptron, resultado de esta investigación, fue construido en KDUGZDUH,
siendo la más antigua de las redes neuronales todavía en uso.
Hasta la fecha, los investigadores en redes neuronales suponían que las conexiones neuronales
en el cerebro eran en su mayor parte aleatorias. Rosenblatt inició una nueva fase en la investigación
neuronal abandonando la idea de conexiones celulares autoformadas, proponiendo que la memoria era
simplemente un cambio en las relaciones entre entradas y salidas, las cuales eran modificadas en su uso
repetitivo, lo que implicaba que patrones regulares y consistentes serían aprendidos. Consecuentemente,
la aleatoriedad en las conexiones neuronales no es desechada, pero sólo ocurre entre distintas capas de
células neuronales. Este tipo de redes de aprendizaje no-supervisado se denominan redes de autoaprendizaje o detectores de patrones regulares. La principal asunción del modelo Perceptron para
Rosenblatt es que “ las conexiones físicas del sistema nervioso que están involucradas en el
reconocimiento y aprendizaje no son idénticas entre organismos. En el nacimiento, la construcción de las
redes más importantes es en su mayor parte aleatoria, sujeta a mínimos cambios delimitados
genéticamente” . [Ros58]
5 /0!W!@9 3:
$ 1 5XL
"? K% 2H &
/0!9S!T@5M@ '& 7U$ 9!CV4& :
$5+5$"
4 9!
El primer modelo Perceptron pretende responder a tres cuestiones fundamentales sobre el
cerebro: como el sistema biológico interpreta o detecta la información del entorno; de qué modo se
almacena o recuerda la información y cómo influencia la información almacenada al reconocimiento y a
la conducta. La única neurona de salida del Perceptron realiza la suma ponderada de las entradas, resta el
umbral y pasa el resultado a una función de transferencia de tipo escalón. La regla de decisión es
responder +1 si el patrón presentado pertenece a la clase A, o –1 si el patrón pertenece a la clase B, la
salida depende de la entrada neta (a= suma de las entradas SR ponderadas).
La red tipo Perceptron emplea principalmente dos funciones de transferencia, KDUGOLP con
salidas 1, 0 o KDUGOLPV con salidas 1, -1; su uso depende del valor de salida que se espera para la red, es
decir si la salida de la red es unipolar o bipolar.
CAPÍTULO 2. FUNDAMENTOS DE LA RED NEURONAL CMAC
13
Una técnica utilizada para analizar el comportamiento de redes como el Perceptrón es presentar
en un mapa las regiones de decisión creadas en el espacio multidimensional de entradas de la red, en estas
regiones se visualiza qué patrones pertenecen a una clase y cuáles a otra, el Perceptrón separa las regiones
por un hiperplano cuya ecuación queda determinada por los pesos de las conexiones y el valor umbral de
la función de activación de la neurona, en este caso los valores de los pesos pueden fijarse o adaptarse
empleando diferentes algoritmos de entrenamiento.
El test más importante para redes neuronales clasificadoras
de patrones consiste en un conjunto de tests en el que la red
neuronales debe ser capaz de discriminar patrones más
pequeños y completamente contenidos en otros patrones más
extensos. Una variación es la conocida como “ or exclusiva” ,
donde dos pequeños patrones diferentes están contenidos
dentro de uno mayor, y los tres deben ser discriminados.
El Perceptron original no siempre es capaz de pasar el test si
sólo utiliza LQSXWV positivos. El uso de LQSXWV QHJDWLYRV
permite solucionar el problema, discriminando los
subconjuntos de patrones, de forma similar al algoritmo de
convolución.
En cualquier caso, siempre se posible
/0!G-!]F > $ $ '%
9 3:/( 9$)
encontrar
un
problema
sencillo que el Perceptron no puede
9 $ 3"
4 <
E')M5" '@& 9!
resolver; ello es debido a su limitación intrínseca: las
fronteras de decisión que establece entre las regiones son
siempre lineales, mientras que en la mayoría de los problemas reales interesantes no lo son. Por otra parte,
el uso de conexiones aleatorias malgasta recursos. La mayoría de las conexiones y relaciones nunca serán
usadas.
Además, el uso de conexiones aleatorias lleva a una explosión combinatoria a medida que el
número y complejidad de los patrones se incrementa. La mayor desventaja de este tipo de redes es su
incapacidad para solucionar problemas que no sean linealmente separables.
En su libro “ Perceptrons” Marvin Minsky y Seymour Papert (MIT, 1969) [Min69], demostraron que el
elemento de proceso introducido por Rosenblatt tenía problemas a la hora de resolver ciertos problemas
específicos.
Minsky y Seymour mostraron que una red neuronal perceptron simple (una capa) no podía:
aprender a simular la función XOR y en distinguir la conectividad existente en ciertas figuras compuestas
(Fig.7).
Minsk y Papert se apartaron de la aproximación
probabilística de Rosenblatt y volvieron a las ideas de cálculo de
predicados en el análisis del Perceptron.
El libro de Minsky y Papert supuso un freno en la
financiación de la investigación en redes neuronales durante más
/0!Y!(D(
3"/0 H$ '5 9 Z $ /4 $ [.
de 10 años. El libro fue ampliamente interpretado como
demostración de que las redes neuronales estaban limitadas en 1"4& @
39$)""/( $$ '@ :
4 $
$ '
:
$Z / $$5\7 ";[
sus conceptos básicos, lo cual fue una conclusión errónea: lo
único que probaron fue las limitaciones del Perceptron
(superadas por el Perceptron Multicapa según ellos mismos
reconocieron).
14
&RQWUROFHUHEUDOGHOPRYLPLHQWR)LVLRORJtDGHO&HUHEHOR
,QWURGXFFLyQ(OVLVWHPDPRWRUFHUHEUDO
El sistema motor cerebral es complejo. Un buen modo de darse cuenta de esto puede ser
mediante un ejemplo. Supongamos que vemos por el rabillo del ojo algo que se está moviendo.
Rápidamente giramos la cabeza y dirigimos la mirada hacia la fuente del movimiento, y descubrimos que
alguien ha dado un golpe a un jarrón de flores y que éste está a punto de caerse de la mesa. Rápidamente
extendemos el brazo, agarramos el jarrón e intentamos volver a situarlo en una posición estable para que
no se caiga.
El movimiento rápido de la cabeza y los ojos está controlado por mecanismos que implican a los
colículos superiores y a algunos núcleos próximos. El movimiento de la cabeza y el movimiento
correspondiente del tronco están mediados por el fascículo tectoespinal. Percibimos cómo el jarrón se
inclina gracias a la actividad de las neuronas de la corteza visual de asociación. Esta misma corteza
proporciona también información sobre la profundidad al lóbulo parietal derecho, cuya corteza de
asociación determina a su vez la localización espacial bastante aproximada del jarrón. Nuestro lóbulo
parietal izquierdo utiliza la información espacial, junto con su propio registro de la localización de la
mano, para determinar el recorrido que ésta ha de realizar para interceptar el jarrón. La información se
envía al lóbulo frontal izquierdo, donde la corteza motora asociativa inicia el movimiento. Ya que éste
tendrá que ser balístico, el FHUHEHOR controlará su pauta temporal, basándose en la información que recibe
de la corteza asociativa de los lóbulos frontal y parietal. Nuestra mano se para en el instante que toca el
jarrón, y las conexiones entre la corteza somatosensorial y la corteza motora primaria inician el reflejo de
cerrar la mano para cogerlo.
El movimiento de la mano se controla mediante la cooperación entre las vías corticoespinales,
rubroespinales y ventromediales. Incluso antes de que nuestra mano se mueva, el fascículo ventral
corticoespinal y las vías ventromediales empezarán a ajustar nuestra postura para evitar que caigamos
cuando vayamos a cogerlo. Dependiendo de lo alejado que se encuentre el jarrón, el fascículo
reticuloespinal puede incluso hacernos dar un paso para no perder el equilibrio. El fascículo rubroespinal
controla los músculos del brazo, y el fascículo corticoespinal lateral controla el movimiento de los dedos
y de la mano. Puede que digamos triunfalmente “ lo conseguí” . La vía corticobulbar, bajo el control de
los mecanismos del habla del hemisferio izquierdo, es la responsable de que los músculos de nuestro
aparato vocal puedan emitir estas palabras [Carl94] .
(O&HUHEHOR,QWURGXFFLyQ
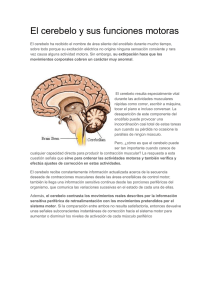
El cerebelo es una parte importante del sistema motor. Consta de dos hemisferios que contienen
varios núcleos profundos situados por debajo de su plegada y arrugada corteza. De este modo, el cerebelo
se parece a un cerebro en miniatura.
/0!,!T@$& N]^"")A
La corteza del cerebelo recibe LQSXWV de la corteza
cerebral, contribuyendo DO FRQWURO GH ORV PRYLPLHQWRV
UiSLGRV \ SUHFLVRV; control del desarrollo temporal de
PRYLPLHQWRV EDOtVWLFRV UiSLGRV, movimientos demasiado
rápidos como para poder ser modificados por circuitos de
realimentación, por que lo que la secuencia de los
movimientos musculares debe programarse con
anterioridad, y los músculos individuales deben activarse
en el momento adecuado, estimando la distancia entre
nuestra mano y el objetivo, nuestro cerebelo calcula la
cantidad de tiempo que los músculos deben estar activos,
CAPÍTULO 2. FUNDAMENTOS DE LA RED NEURONAL CMAC
15
transcurrido ese tiempo, el cerebelo activa brevemente los músculos antagonistas para detener el
movimiento; otra de las funciones principales del cerebelo consiste en SURJUDPDU OD GXUDFLyQ GH ORV
PRYLPLHQWRVUiSLGRV.
El cerebelo YLJLOD \ HVWDEOHFH DMXVWHV FRUUHFWRUHV GH ODV DFWLYLGDGHV PRWRUDV GHVHQFDGHQDGDV
SRU RWUDV SDUWHV GHO HQFpIDOR. Recibe continuamente información actual de las partes periféricas del
cuerpo, para determinar el estado instantáneo de cada uno de sus áreas - su posición, su ritmo de
movimiento, las fuerzas que actúan sobre él, etc. El cerebelo compara el estado físico actual de cada parte
del cuerpo según indica la información sensorial, con el estado que intenta producir el sistema motor. Si
los dos no se comparan favorablemente, de manera instantánea se trasmiten señales correctoras adecuadas
hacia el sistema motor, para aumentar o disminuir la actividad de músculos específicos.
El cerebelo se comunica con el cerebro a través de unos cordones de fibras llamadas pedúnculos
superiores, con la protuberancia anular por los pedúnculos medios y con el bulbo raquídeo por los
pedúnculos inferiores.
La sustancia gris contiene células en las cuales se originan fibras que van a formar sinapsis con
los que provienen de otras partes del encéfalo y que penetran al cerebelo. Los impulsos de los centros
motores del cerebro, de los conductos semicirculares del oído interno y de los músculos estriados llegan
al cerebelo por los pedúnculos. Los impulsos motores del cerebelo son transmitidos hacia los centros
motores del cerebro y de la medula con destino a los músculos.
[Carl94]
(O6LVWHPDGH(QWUDGDGHO&HUHEHOR
El cerebelo se divide en tres lóbulos: lóbulo floculonodular, lóbulo anterior, y lóbulo posterior.
En el lóbulo anterior y posterior se encuentra la vermis. En ésta terminan la mayor parte de las señales
nerviosas que provienen de las áreas somáticas del cuerpo. La vermis tiene un papel en la integración de
los mecanismos posturales subconscientes. En los hemisferios cerebelosos terminan la mayor parte de las
señales que llegan de los niveles más altos del cerebro, en especial de las áreas motoras de la corteza
cerebral.
La vía corticocerebelosa nace en la corteza motora enlazando directamente con la corteza del
cerebelo. Además, vías aferentes importantes nacen del tallo cerebral; uniendo la oliva inferior, corteza
motora, ganglios basales, zonas dispersas de la formación reticular y la médula espinal, a todas las zonas
del cerebelo.
5 /0!9+_!M) '%
9 $%$`
4 $)59& "
4" El cerebelo también recibe señales sensoriales importantes directamente de la periferia del
cuerpo. Las señales transmitidas por estos haces nacen de los husos musculares, los órganos tendinosos de
Golgi, y los grandes receptores táctiles de piel y articulaciones, que informan al cerebelo del estado actual
de la contracción muscular, el grado de tensión de los tendones, las posiciones de las partes corporales, y
las fuerzas que actúan sobre las superficies del cuerpo. Toda esta información es conservada en el
cerebelo, constantemente informado del estado físico instantáneo del cuerpo.
Las vías espinocerebelosas pueden trasmitir impulsos a velocidades mayores de 100m por
segundo, que es la conducción más rápida en cualquier vía de todo el sistema nervioso central. Esta
conducción extraordinariamente rápida permite que instantáneamente el cerebelo conozca los cambios
que se están produciendo en el estado de los músculos. El cerebelo recibe información continua de todas
las partes del cuerpo, aunque estén operando a nivel subconsciente.
La principal entrada al cerebelo es portada por fibras musgosas. Las fibras musgosas llevan
información de fuentes que controlan el equilibrio (sistema vestibular), la alerta (formación reticular), la
actividad motora (cortex cerebral), órganos sensores y de medida de cantidad en la posición de tendones,
velocidad de contracción de músculos, y presión de la piel.
Las ILEUDVPXVJRVDV pueden ser clasificadas por el punto base de origen en dos clases: aquellas
que llevan información que podrían incluir órdenes de niveles jerárquicos más altos, y las que transportan
información de realimentación sobre resultados de la salida del control motor. Una vez en el cerebelo, los
dos clases se mezclan, resultando indistinguibles.
[Alb81] [DSE94]
/RFDOL]DFLyQWRSRJUiILFDGHOLQJUHVR
VHQVRULDODODFRUWH]DFHUHEHORVD
No ha sido posible localizar las diferentes partes del cuerpo en la
mayoría de las áreas del cerebelo. Sólo se ha encontrado una
representación general de las estructuras de la línea media.
Muchos neurofisiólogos creen probable que haya relaciones
espaciales, punto a punto, entre hemisferios y áreas
estimuladoras específicas de la corteza cerebral, dado
que
cuantas veces se transforma una señal motora a la periferia al
mismo tiempo se encuentra una señal hacia los hemisferios
cerebrales.
6HxDOHVGHVDOLGDGHOFHUHEHOR
5 /0!a++!FP9J') 19 ^a& ;bc" d$:/Gef
'@9& g^ / hi"
4 29& 9 hj $19 $
2$B/ $
$EAa$& Fe & 23L9H 7$
4 L $3k $
9EM"Q =!
Localizados profundamente en la masa cerebelosa hay cuatro núcleos cerebelosos profundos.
Estos núcleos reciben señales de dos fuentes diferentes: la corteza cerebelosa, y todas las vías sensoriales
aferentes hacia el cerebelo. La entrada en el cerebelo es portada por las fibras musgosas del cerebro, hacia
donde salen tres importantes vías eferentes:
-
Una que se inicia en la corteza de los dos hemisferios cerebelosos, que se dirige a la corteza
motora.
Una vía que se origina en las estructuras de la línea media del cerebelo (vermis) hacia las
regiones bulbares y pontinas del tallo encefálico. Este circuito funciona en íntima relación con
el aparato del equilibrio y las actitudes postulares del cuerpo.
CAPÍTULO 2. FUNDAMENTOS DE LA RED NEURONAL CMAC
-
17
Una vía que se origina en las áreas intermedias a cada lado del cerebelo, entre el vermis y los
hemisferios cerebelosos, a la corteza motora, a los ganglios basales, al núcleo rojo y la
formación reticular de la porción superior del tallo encefálico. Este circuito funciona para
coordinar las actividades entre las dos primera vías cerebelosas de salida comentadas - es decir,
para ayudar a coordinar las interrelaciones entre el control de la postura subconsciente del
cuerpo y el control consciente voluntario de la corteza motora. La formación reticular tiene
funciones relacionadas con la alerta.
(OFLUFXLWRQHXURQDOGHOFHUHEHOR
El cerebelo humano es una lámina grande fruncida, de unos 17 cm de anchura por 120 cm de
largo; los pliegues se cruzan. Cada pliegue se llama lámina. El cerebelo tiene unos 30 millones de
unidades funcionales casi idénticas. Esta unidad funcional se centra alrededor de la célula de 3XUNLQMH;
hay también 30 millones de estas células en la corteza cerebelosa.
Las tres capas principales del cerebelo son la capa molecular, la capa de FpOXODVGH3XUNLQMH y la
capa de FpOXODVJUDQXODUHV. Además de estas capas, los núcleos profundos están localizados en el interior
de la masa cerebelosa. La salida de la unidad funcional proviene de una célula de núcleo profundo.
Esta célula se halla continuamente bajo la influencia de estimulaciones de ambos tipos,
H[FLWDGRUDV e LQKLELGRUDV. Las influencias inhibidoras provienen totalmente de las células de 3XUNLQMH de
la corteza cerebelosa.
Las HQWUDGDV en el cerebelo son de dos tipos, uno llamado de tipo de ILEUDWUHSDGRUD y el otro de
tipo de ILEUD PXVJRVD. Hay una fibra trepadora por cada 10 células de Purkinje, aproximadamente.
Después de mandar señales colaterales a las diversas células nucleares profundas, la ILEUDWUHSDGRUD se
proyecta a todo lo largo hasta la capa molecular de la corteza cerebelosa, donde establece unas 300
sinapsis con las dendritas de cada célula de Purkinje.
Las ILEUDV PXVJRVDV también mandan señales colaterales para excitar células nucleares
profundas. Luego estas fibras proceden siguiendo hacia la capa granular de la corteza, donde se
establecen sinapsis con centenares de FpOXODVJUDQXODUHV.
5 /0!G+C9!#%$"l92
L i& B<
F !Gm@:9$7 1"2
9 793"
4 24& 9 $a& :
$ ')& 9$G'3 $919 :
$2$;/0$
$LZ 2$:$ A)19 :$ [.
n
oI o"
4 <
n4& & oBb9 & 9& dZ p[.n>
$i 4& $iIk $7/4"
9 "$qZ /4"
4 I $ [!^KE
$ak $Fsr5 /()"9
23 4& s $)19 :
$'%
9 $I59e2nqk $/4"
9 "$i
9& >
$!hKE
$Ik $aFM5:Q =q"
4 <
n@$2
4& n5 1& 7')$ & >
')";') 4 $19 :
$9'%
"
9 $)l45" !7
29 t$2
H 1& 9:/ & >7Z ? [95k $H
$& Z $"Q&9 [H 9>:$
$!@KE
3$2
$& $'$$& 2 L 01 39& :
$ 1 !
18
Estas células, a su vez, mandan sus axones muy cortos, hasta la superficie externa de la corteza
cerebelosa para penetrar en la capa molecular más superficial. Aquí los axones se dividen en dos ramas.
Es en esta capa molecular donde las dendritas de las células de Purkinje se proyectan, y a cada célula de
Purkinje hace sinapsis con de 80.000 a 200.000 de estas ILEUDV SDUDOHODV. La estimulación de una sola
fibra musgosa nunca desencadenará un potencial de acción en la FpOXODGH3XUNLQMH; por el contrario, hay
que estimular gran número de células musgosas simultáneamente para activar la FpOXODGH3XUNLQMH.
5 /0!+N!T)"
HHN]^5k $ AE1 "
$)
" Las FpOXODVGH*ROJL son también contactadas por las fibras paralelas. Estas células poseen un
denso árbol dentrítico. Cada célula de *ROJL tiene un axón que se ramifica extensamente, realizando
contacto inhibitorio (-) con alrededor de 100.000 células granulares en su inmediata vecindad, incluyendo
algunas de las células granulares (ramificadas en fibras paralelas), que las excitaron. Las FpOXODVGH*ROJL
suprimen las salidas de todas las FpOXODV JUDQXODUHV que no están máximamente estimuladas. Como
resultado, cada patrón o vector de entrada, es transformado por la capa granular en un pequeño y
relativamente fijo porcentaje, o subconjunto, de fibras paralelas activas.
Cada FpOXOD GH 3XUNLQMH realiza un sumatorio sobre sus entradas (sinapsis), produciendo una
única salida, siendo ésta la salida del cortex cereberal.
Las FpOXODVFDQDVWR y las HVWUHOODGDV, son esencialmente inversores, que proveen a la FpOXODGH
3XUNLQMH de pesos negativos, que son sumados a los pesos positivos de las fibras paralelas.
Las FpOXODV WUHSDGRUDV son el segundo conjunto de fibras entrantes del córtex cerebelar.
Típicamente existe una célula trepadora por célula de Purkinje, ajustando el peso de las conexiones
sinápticas con las fibras paralelas, alterando la salida de la célula. Las células trepadoras podrían tener
alguna relación proveyendo la información necesaria para el aprendizaje.
[Alb81] [Alb71] [DSE94]
&RQWUROFHUHEHORVRSRUUHWURDOLPHQWDFLyQGHODIXQFLyQPRWRUD
FRUWLFDO
La corteza motora trasmite señales a la periferia para causar una función motora, pero al mismo
tiempo transmite esta información al cerebelo. La corteza motora manda muchos más impulsos que los
que se necesitan para realizar cada movimiento, y el cerebelo debe inhibir la corteza motora en el
momento apropiado cuando el músculo ha empezado a moverse. El cerebelo aprecia automáticamente la
velocidad del movimiento y calcula el tiempo que se necesitara para alcanzar el punto deseado.
CAPÍTULO 2. FUNDAMENTOS DE LA RED NEURONAL CMAC
19
Luego se trasmiten a la corteza motora los impulsos correspondientes, que inhiben los músculos
agonistas y activan los antagonistas. En esta forma, se dispone de un "freno" adecuado para detener el
movimiento en el punto preciso.
Aprender la inercia del sistema es función importante del mecanismo cerebeloso de
retroalimentación, aunque es probable que este aprendizaje tenga lugar en la corteza cerebral más bien
que en el cerebelo.
Cuando se realiza un movimiento rápido hacia cierto punto, los músculos agonistas se contraen
violentamente durante la primera parte del movimiento, luego, de repente, poco tiempo antes que se
alcance el punto en cuestión, los músculos agonistas se inhiben completamente, mientras se excitan
considerablemente los antagonistas. La activación de los músculos antagonistas cerca del final de un
movimiento es función enteramente automática y subconsciente, y de ninguna manera constituyente una
contracción "voluntaria" similar a la contracción inicial del músculo agonista.
Un efecto secundario del mecanismo cerebeloso de retroalimentación es su capacidad de
"DPRUWLJXDU" los movimientos musculares.
Debido a la inercia, todos los movimientos pendulares, la mayoría de los del cuerpo, tienen
tendencia a sobrepasar la posición del propósito inicial. Si el cerebelo está intacto, señales subconscientes
apropiadas detienen el movimiento exactamente en el sitio requerido, evitando así que se pase de él y
suprimiendo el temblor de amortiguación, característica básica de un sistema de amortiguación.
Otro efecto colateral importante del mecanismo cerebeloso de retroalimentación es que ayuda al
sistema nervioso central a SUHGHFLU las posiciones futuras de todas las partes móviles del cuerpo. Sin el
cerebelo esta función pronosticadora es tan deficiente que las partes del cuerpo en movimiento rápido se
desplazarían mucho más allá del punto de intención.
5HVXPHQ
La memoria se almacena en los sistemas biológicos neuronales mediante refuerzo y creación de
relaciones entre las neuronas. Los patrones de entrada del sistema nervioso, nuestro entorno y
experiencias, modifican la estructura neuronal. La conducta pues es resultado del procesamiento de estos
patrones, creando a su vez cambios que afectan al entorno.
Los psicólogos conductistas, consideran al sistema de proceso como una caja negra, con entradas
(estímulos) y una salida (conducta), con capacidad de aprendizaje basada en estímulos y respuestas. El
condicionamiento instrumental explica el aprendizaje básico, como el motor, en estímulos reforzantes de
conductas, mediante recompensa y castigo. Los patrones de entrada que realizan una conducta “ deseada”
son reforzados, aumentando la probabilidad de conductas “ deseadas” en los patrones aprendidos.
Las redes neuronales realizan una modelización del funcionamiento de los circuitos neuronales.
A cada elemento de la red se le otorgan propiedades similares a las de las neuronas, y se les conecta entre
sí por medio de uniones similares a la sinapsis. Estas uniones ejercen efectos excitatorios o inhibitorios,
enseñando a las redes neuronales a reconocer estímulos concretos. Las redes neuronales muestran
generalización, discriminación y degradación suave, los cuales caracterizan también el funcionamiento de
nuestro cerebro.
La comprensión de la micro y macroestructura cerebral proporciona una base fisiológica sobre la
que asentar las redes neuronales artificiales.
El Perceptron fue el primer detector de patrones regulares implementado imitando las
conexiones neuronales del cerebro. El primer modelo Perceptron pretendía responder como el sistema
biológico interpreta o detecta la información del entorno; de qué modo se almacena o recuerda la
información; y cómo influencia la información almacenada al reconocimiento y a la conducta.
El comportamiento de redes como el Perceptron puede analizarse como un mapa de regiones de
decisión creadas en el espacio multidimensional de entradas de la red. Un hiperplano separa qué patrones
pertenecen a una clase y cuáles a otra. La ecuación del hiperplano está determinada por los pesos de las
20
conexiones y el valor umbral de la función de activación de la neurona. Los valores de los pesos pueden
fijarse o adaptarse empleando diferentes algoritmos de entrenamiento.
La red Perceptron básica presentaba problemas en el reconocimiento de patrones contenidos en
otros patrones mayores y resolución de problemas no-lineales.
El sistema motor cerebral es complejo, implica multitud de órganos sensoriales, de decisión y
actuadores del cuerpo humano, entre ellos el cerebelo. El cerebelo es una parte importante del sistema
motor. La corteza del cerebelo recibe inputs de la corteza cerebral, contribuyendo al control de los
movimientos rápidos y precisos; control del desarrollo temporal de movimientos balísticos rápidos; y de
movimientos demasiado rápidos como para poder ser modificados por circuitos de realimentación, por
que lo que la secuencia de los movimientos musculares debe programarse con anterioridad.
Los impulsos motores del cerebelo, constantemente informado del estado físico instantáneo del
cuerpo, son transmitidos hacia los centros motores del cerebro y de la medula con destino a los músculos.
La principal entrada al cerebelo es portada por fibras musgosas. Las fibras musgosas llevan información
de fuentes que controlan el equilibrio (sistema vestibular), la alerta (formación reticular), la actividad
motora (cortex cerebral), órganos sensores y de medida de cantidad en la posición de tendones, velocidad
de contracción de músculos, y presión de la piel.
Las fibras musgosas podrían transmitir información como órdenes de niveles jerárquicos más
altos e información de realimentación sobre resultados de la salida del control motor. El cerebelo tiene
unos 30 millones de unidades funcionales casi idénticas. Esta unidad funcional se centra alrededor de la
célula de Purkinje. Esta célula se halla continuamente bajo la influencia de estimulaciones excitadoras e
inhibidoras.
Las entradas en el cerebelo son de dos tipos, uno llamado de tipo de fibra trepadora y el otro de
tipo de fibra musgosa. Las fibras musgosas mandan señales colaterales hacia la capa granular de la
corteza, donde se establecen sinapsis con centenares de células granulares. Estas células dividen sus
axones en dos ramas de fibras paralelas. Las fibras paralelas sinaptan con las células de Purkinje. Es
necesario estimular gran número de células musgosas simultáneamente para activar la célula de Purkinje.
Las células de Golgi son también contactadas por las fibras paralelas. Las células de Golgi
suprimen las salidas de todas las células granulares que no están máximamente estimuladas. Como
resultado, cada patrón de entrada, es transformado en un pequeño y relativamente fijo porcentaje, o
subconjunto, de fibras paralelas activas.
Cada célula de Purkinje realiza un sumatorio sobre sus entradas (sinapsis), produciendo una
única salida, siendo ésta la salida del cortex cereberal. Las células canasto y las estrelladas, suman sus
pesos negativos al sumatorio de la célula de Purkinje.
Las células trepadoras son el segundo conjunto de fibras entrantes del cortex cerebelar.
Típicamente existe una célula trepadora por célula de Purkinje, ajustando el peso de las conexiones
sinápticas con las fibras paralelas, alterando la salida de la célula. Las células trepadoras podrían tener
alguna relación proveyendo la información de realimentación necesaria para el aprendizaje.
Un efecto secundario del mecanismo cerebelar de retroalimentación es su capacidad de
"amortiguar" los movimientos musculares, y ayudar al sistema nervioso central a predecir las posiciones
futuras de todas las partes móviles del cuerpo.