La contracción muscular - Maestría en Salud Integral y Movimiento
Anuncio

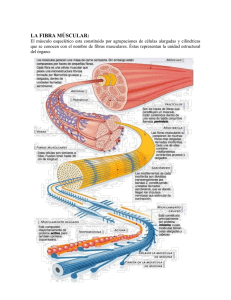
UNIVERSIDAD NACIONAL FACULTAD CIENCIAS DE LA SALUD ESCUELA CIENCIAS DEL DEPORTE MAESTRIA EN SALUD INTEGRAL Y MOVIMIENTO HUMANO Fisiología Avanzada del Movimiento Humano Tema: Mecánica de la contracción muscular Profesor: M Sc. Juan Carlos Gutiérrez Estudiantes: Alejandra Delgado Carolina Ruiz Karla Solis Marianela Obando Nahida Andrawus I Trimestre 2006 Introducción Los músculos son tejidos que permiten la movilización del cuerpo. Según Wilmore y Costill (2001), existen tres tipos de músculos: § El liso § El cardiaco § El esquelético Cada uno cumple funciones con diferentes directrices, permitiendo así el movimiento. Si una persona o animal sufre algún daño en el tejido de alguno de estos músculos es probable que se minimice su función. El músculo liso se denomina involuntario puesto que su funcionamiento no depende directamente de nuestra conciencia. Están ubicados en las paredes de la mayor parte de vasos sanguíneos lo cual les permite contraerse o dilatarse con el propósito de regular el flujo sanguíneo (Wilmore y Costill, 2001). Los músculos esqueléticos denominados voluntarios unen y mueven el esqueleto. El cuerpo humano contiene más de 215 parejas de este tipo de músculos. Por ejemplo, el dedo pulgar cuenta con 9 músculos independientes (Wilmore y Costill, 2001). El músculo cardiaco está ubicado en el corazón y abarca la mayor parte de la estructura. Se controla así mismo mediante los sistemas nervioso y endocrino (Wilmore y Costill, 2001). Como ya se sabe, el ejercicio requiere de los músculos para poder producir su movimiento, pese a las diferencias plasmadas anteriormente su accionar es similar y se requieren en conjunto para lograr esto y poder realizar actividad física. El presente trabajo se desarrolla con el objetivo de ahondar en la temática de la contracción muscular y su importancia radica en que los músculos que componen el cuerpo humano constantemente se están contrayendo para realizar cualquier tipo de movimiento. 2 Mecánica de la contracción muscular Resulta imprescindible abordar en primera instancia ciertas definiciones que faciliten una mejor comprensión de la mecánica de la contracción muscular, las cuales se detallan a continuación. Estructura muscular Los músculos están compuestos en su exterior por tejido conectivo que según Álvarez (2000), es un tejido denso que sirve como sostén. Sin embargo, en su interior, constan de una serie de estructuras. Primeramente se encuentra el epimisio que rodea al músculo, al cortar éste se ubica el perimisio que cubre fascículos que son pequeñas haces de fibras. Finalmente se hallan las fibras musculares individuales, a su vez, cada una de estas está cubierta por una vaina de tejido conectivo (Wilmore y Costill, 2001). Al observar detalladamente una fibra muscular compuesta por varias miofribrillas que abarcan su misma longitud, se ve que está rodeada por una membrana de plasma llamada sarcolema que se funde con el tendón insertándose en el hueso. Dentro del sarcolema se encuentra una sustancia similar a gelatina llamado sarcoplasma que contiene proteínas, minerales, glucógeno, grasas disueltas y organelas. Dentro del sarcoplasma también se encuentra una extensa estructura de túbulos transversales (llamados Túbulos T) que pasan por entre las miofibrillas y permiten que los impulsos nerviosos que recibe el sarcolema sean transmitidos a las miofibrillas individuales compuestas por subunidades más pequeñas llamadas sarcómeros. Cada sarcómero es la zona de la miofibrilla situada entre dos líneas Z. El sarcómero es considerado como la unidad funcional contráctil del músculo estriado (Luttgens y Wells, 1982). Las miofibrillas microscópicas, son elementos contráctiles, ordenadas de forma paralela dentro de la fibra y formando bandas alternas oscuras y claras que le dan el aspecto estriado a la fibra muscular. El microscopio electrónico ha mostrado que las estrías son un patrón de repetición de bandas y líneas debido a una interdigitación de dos grupos de filamentos. Se 3 ha postulado que estos filamentos de proteínas contráctiles, principalmente de actina y miosina al ser estimuladas se deslizan entre sí. (Luttgens y Wells, 1982). También se encuentra una red longitudinal de túbulos llamados retículos sarcoplasmásticos que sirven como depósito para el calcio (Wilmore y Costill, 2001). Figura N. 1. Estructura del músculo 4 Propiedades del tejido muscular Las propiedades del músculo estriado son: extensibilidad, elasticidad y contractibilidad. Las dos primeras capacitan al músculo para estirarse como una banda elástica y volver de nuevo a su longitud normal en reposo, cuando la fuerza de extensión se interrumpe (Luttgens y Wells, 1982). La contractibilidad es la característica que permite el acercamiento de las fibras entre sí. De esta manera hay una diferencia entre la longitud máxima y mínima de una fibra muscular conocida como amplitud de su acción (Luttgens y Wells, 1982). Miofibrilla El componente de la célula muscular es la miofibrilla. Esta contiene dos filamentos proteicos fundamentales, uno más espeso denominado miosina y otro más delgado que recibe el nombre de actina (Bowers y Fox, 1998) Estas proteínas presentan una distribución geométrica en el músculo y otorga su apariencia en bandas o estriadas y forman los componentes activos en el proceso contráctil y representan el lugar de utilización de la energía, ósea facilitan la contracción muscular (Bowers y Fox, 1998). Sarcómero Se lo define como la distancia entre dos “lineas Z” (Bowers y Fox, 1998). El músculo se caracteriza por la presencia de zonas claras y oscuras que se alternan. La banda I de un sarcómero está constituida sólo por filamentos de actina que se extienden desde las líneas Z hacia el centro del sarcómero. La banda A está constituida por filamentos de actina y de miosina (Bowers y Fox, 1998). Las diminutas proyecciones que se extienden desde los filamentos de miosina hacia los filamentos de actina reciben el nombre de puentes de miosina. Estas proyecciones son el instrumento que hacen posible el acortamiento del músculo durante la contracción. Una 5 zona en el centro de la banda A que está constituida principalmente por filamentos de miosina, se denomina zona H. Durante la contracción, la zona H se volverá más pequeña o desaparecerá a medida que la actina y la miosina se deslicen entre sí (Bowers y Fox, 1998). Figura N. 2 Sarcómero La contracción muscular Según Jean Hanson y Hugo Huxley en la época de 1950, la contracción muscular debía ser un proceso de plegamiento. Sin embargo, Hanson y Huxley propusieron que el músculo esquelético se acorta durante la contracción debido a que sus filamentos gruesos y finos se deslizan unos sobre otros. Su modelo es conocido como el mecanismo de deslizamientos de los filamentos de la contracción muscular (Tórtora y Gabowski, 1998). Durante la contracción muscular los puentes transversales de la miosina tiran de los filamentos finos, haciendo que se deslicen hacia dentro en dirección a la zona H. Cuando los puentes transversales tiran de los filamentos finos, éstos acaban por encontrarse en el centro de la sarcómero (Tórtora y Gabowski, 1998). A medida que los filamentos finos van deslizándose hacia dentro, los discos Z van aproximándose entre ellos y la sarcómera se acorta, pero la longitud de los filamentos gruesos y finos no cambia. El deslizamiento de los filamentos y el acortamiento de las 6 sarcómeras determinan el acortamiento de la totalidad de la fibra muscular y, en último termino, de todo el músculo (Tórtora y Gabowski, 1998). Figura N.3. Deslizamiento de filamentos del sar cómero El inicio del deslizamiento se debe a un aumento de la concentración de Ca 2+ en el sarcoplasma, mientras que un descenso de dicha concentración interrumpe el proceso del deslizamiento. Esto sucede cuando la fibra muscular está relajada la concentración de Ca en el sarcoplasma es bajo. Ello se debe a que la membrana del retículo sarcoplamático contiene bombas para el transporte activo del Ca 2+ , que eliminan el calcio del sarcoplasma (Tortora y Gabowski, 1998). Los iones de calcio liberados del retículo sarcoplasmático se combinan con troponina, haciendo que cambie de forma, lo que hace que el complejo troponina­tropomiosina se separe de los lugares de unión a la miosina que posee la actina (Tórtora y Gabowski, 1998). La contracción muscular requiere de Ca 2+ y energía en forma de ATP (Adenosin Trifostato). El ATP llega a los lugares de unión del ATP existentes en los puentes transversales de la miosina. Una porción de cada cabeza de miosina actúa como una ATPasa, enzima que divide el ATP en ADP + fósforo (P) mediante una reacción de hidrólisis. Esta reacción transfiere energía desde el ATP a la cabeza de la miosina, incluso antes de que inicie la contracción muscular. Los puentes transversales de la miosina se encuentran en un estado activado. Cuando el nivel del Ca 2+ se eleva y la tropomiosina se desliza y abandona su posición de bloqueo, estas cabezas de miosina activadazas se unen espontáneamente a los lugares de unión de la miosina existentes en la actina. El cambio de 7 forma que se produce cuando la miosina se une a la actina genera el golpe de potencia de la contracción (Tórtora y Gabowski, 1998). Durante el golpe de potencia de los puentes transversales de la miosina, giran hacia el centro del sarcómero como los remos de un bote. Esta acción arrastra a los filamentos finos sobre los filamentos gruesos hacia la zona H. Las cabezas de la miosina giran a medida que van liberando el ADP (Adenosin Difosfato), (Tortora y Gabowski, 1998). Una vez completado el golpe de potencia, el ATP se combina de nuevo con los lugares de unión del ATP que poseen los puentes transversales de la miosina. Cuando esta unión se produce, las cabezas de la miosina se separan de la actina. De nuevo se produce la degradación del ATP, lo que proporciona energía a la cabeza de miosina, que recupera su posición recta original, momento en el que vuelve a estar dispuesta para combinarse con otro lugar de unión de la miosina del filamento fino que se encuentre en una posición más alejada (Tórtora y Gabowski, 1998). El ciclo se repite una y otra vez. Los puentes transversales de miosina se mantienen en movimiento, hacia atrás y adelante con cada golpe de potencia, desplazando a los filamentos finos hacia la zona H. (Tórtora y Gabowski, 1998). Los golpes de potencia se repiten mientras exista ATP disponible y el nivel de Ca 2+ cerca del filamento fino se mantenga alto (Tórtora y Gabowski, 1998). Después de la contracción, dos cambios permiten que la fibra muscular vuelva a relajarse: · La acelticolina (Ach) es rápidamente degradada por una enzima llamada acelticolinesterasa (AchnE), ésta se encuentra en la hendidura sináptica. Cuando los potenciales de acción cesan en la neurona motora, no se libera más Ach y la AchnE degrada con rapidez la Ach ya existente en la hendidura sináptica. Con ello se detiene la generación de potenciales de acción muscular y los canales de liberación del Ca 2+ del retículo sarcoplásmatico se cierran. (Tortora y Gabowski, 1998). 8 · En segundo lugar, las bombas de transporte activo del Ca 2+ eliminan con rapidez el Ca 2+ existentes en el sarcoplasma pasándolo al interior del retículo sarcoplásmatico (Tórtora y Gabowski, 1998). Cuando el nivel de Ca 2+ cae en el sarcoplasma, el complejo tropomisina­troponina vuelve a deslizarse sobre los lugares de unión de la miosina existentes en la actina, lo que impide que los puentes transversales de la miosina se unan a la actina, de forma que los filamentos finos recuperan su posición relajada (Tórtora y Gabowski, 1998). La teoría del filamento deslizante en la contracción muscular según Bowers y Fox, no se conoce plenamente el mecanismo implicado en el desplazamiento. Sin embargo, existe un consenso bastante amplio sobre el hecho de que al producirse la estimulación de un músculo los puentes de miosina forman un tipo de unión con sitios seleccionados en los filamentos de actina (Bowers y Fox, 1998). Este proceso de acoplamiento algunas veces denominado formación de actomiosina, dependen de la presencia de iones de calcio. Una vez fijados, los puentes giran de tal manera que los filamentos de actina son atraídos sobre la miosina y hacia el centro del sarcomero. Durante este proceso el ATP es degradado a ADP y a P, el músculo se acorta y se desarrolla tensión. Cuando la estimulación se detiene el músculo se relaja y regresa a su estado de reposo (Bowers y Fox, 1998). 9 Figura N.4. Mecánica de la contracción Ley del todo o nada Para que un músculo se contraiga es necesario que se genere un potencial de acción, es decir que se “acumule ”la suficiente energía para que se dé da contracción. Esto sucede a partir de los ­70 miliVoltios, estos es porque el interior de la membrana celular está cargada con iones negativos. Conforme se va acercando al umbral propio de la célula muscular (­70 mV) las probabilidades que se de la contracción son mayores. Si llega a este, se contrae, y si el estímulo se mantiene, la contracción se mantiene; pero por el contrario, si no se da, este no se contrae. Es por eso que se dice que responde a la Ley del todo o nada (Ganong, 1998). Fibras muscular es de contracción lenta y de contracción rápida En el cuerpo humano, las fibras musculares no son iguales; un mismo músculo puede tener dos tipos diferentes, a saber fibras de contracción lenta (ST, en inglés Show­twich) y de contracción rápida (FT, en inglés fast­twich). 10 Las fibras de contracción lenta necesitan 110 ms para alcanzar su tensión cuando son estimuladas, mientras que las fibras de contracción rápida alcanzan su máxima tensión en aproximadamente 50 ms. Las fibras FT se clasifican en FTa (Contracción rápido de tipo a), las FTb de contracción rápida de tipo b y las FTc. Las diferencias no se comprenden del todo, pero se cree que las más utilizadas son las FTa y las que se usan con menos frecuencia son las FTc. Características de las fibras ST y FT Las fibras de contracción lenta y rápida se denominan de esta forma por su velocidad de acción que el resultado de diferentes formas de miosina ATPasa que es la enzima que libera la energía para producir la contracción, por lo que las fibras FT disponen de energía más rápido que las ST. Las FT tienen un retículo sarcoplamático más desarrollado que las otras y son las neuronas las que parecen determinar la diferencia entre estas. Tabla N. 1. Tipos de fibras muscular es Clasificación de las fibras Sistema 1 Contracción lenta Contracción rápida a Contracción rápida b Sistema 2 Tipo I Tipo IIa Tipo IIb Sistema 3 OL GOR G Capacidad oxidativa Alta Moderadamente alta Baja Capacidad glucolítica Baja Alta La más alta Velocidad contráctil Lenta Rápida Rápida Resistencia a fatiga Alta Moderada Baja Fuerza de unidad motora Baja Alta Alta Característica Fuente: Wilmore y Costill, 2001; pag 35. Es importante rescatar que las fibras de contracción lenta se caracteriza por tener una elevada resistencia aeróbica por lo que se movilizan con más frecuencia durante las pruebas de resistencia de baja intensidad, como por ejemplo una maratón. 11 Las fibras de contracción rápida sirven para rendir anaeróbicamente por lo que se fatigan más fácilmente. Por ejemplo: las carreras de una milla. Cuando una persona envejece, los músculos pierden fibras rápidas y se incrementa el porcentaje de fibras de contracción lenta (Wilmore y Costill, 2001). Figura N.5 Uso de las diferentes fibras musculares según tensión Conclusión Para poder llevar a cabo la mecánica de la contracción muscular se requieren una serie de estructuras que conjuntamente unifican sus funciones y características para realizar el movimiento. A si mismo, el cuerpo va a necesitar sustancias que provean energía o que participen en el proceso para obtenerla, como el fósforo para producir el ATP (Adenosin Trifosfato) que será la forma de energía a utilizar. 12 A la vez se requieren de otros nutrientes, en esto mencionamos minerales como el calcio considerado como indispensable para lograr la contracción muscular. Además el organismo tiene en sus músculos diferentes tipos de fibras que se complementan para realizar la actividad física, y cada una se utilizará de acuerdo al tipo de actividad física, ya sea aeróbica o anaeróbica. Como se mencionó en un principio, una lesión en algún componente estructural que conforma la mecánica de la contracción muscular es posible que limite de alguna manera la capacidad del movimiento. Bibliografía Alvarez, Juan. (2000). Diccionario Mosby. Medicina, Enfermería y Ciencias de la Salud. Quinta Edición. Harcourt España. Madrid. Bowers Richard, Fox Edgard. (1998). Fisiología del Deporte. Tercera Edición. Editorial Panamericana. México. Ganong, William. (1998). Fisiología Médica. Décimo sexta Edición. Editorial El Manual Moderno. México. Luttgens Kathryn, Wells Katharine. (1982) Kinesiología Bases Científicas del Movimiento Humano. Sétima Edición. España. Tortora Gerard, Grabowski Sandra. (1998). Principios de Anatomía y Fisiología. Sétima Edición. España. Wilmore R y Costill, D. (2001). Fisiología del Esfuerzo y del Deporte. Cuarta Edición. Editorial Paidotribo. Barcelona. 13