Embriogénesis Temprana de los Vertebrados
Anuncio

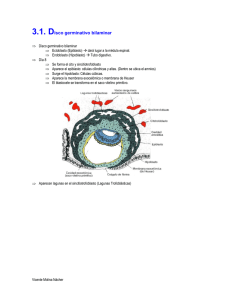
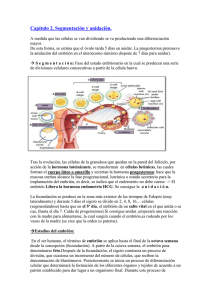
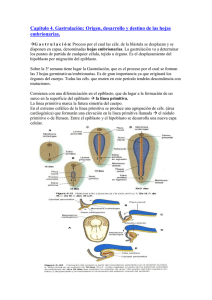
Embriogénesis Temprana de los Vertebrados Alexander Vargas Laboratorio de Ontogenia y Filogenia Depto Biología, Facultad de Ciencias, U. de Chile PDF en hEp://vargaslab.wordpress.com Divisiones celulares tempranas dan lugar a la blástula o embrión temprano La canLdad de vitelo (“yema”) en el ovocito limita la división de las celulas (iso, meso o telolecíLco) La blástula genera (más tarde o más temprano) una cavidad interna (Blastocele) que presenta un epiblasto dorsal (“techo” del blastocele) y un hipoblasto ventral (“piso” del blastocele) En muchos animales, incluyendo vertebrados basales, tanto el epiblasto como el hipoblasto contribuyen al cuerpo del embrion, con el epiblasto dando origen a epidermis y nervios, y el hipoblasto dando origen a estomago‐visceras. En amniotos de huevo teloleciLco, hay un espacio entre el embrión y el vitelo que no corresponde al blastocele (espacio subgerminal). En estas formas, y en mamíferos ocurre la formación de un hipoblasto secundario Formación del “hipoblasto secundario” en embrión discoidal de aves (huevo telolecíLco) El hipoblasto se desarrolla por celulas que ingresan por delaminacion del epiblasto que se integran con celulas de la zona marginal (futura zona posterior) que migran hacia anterior. El hipoblasto en amniotos con huevo teloleciLco comunica la yema (vitelo) con el estómago del embrión. La zona pellucida es una matriz extracelular protectora que parLcipa en el proceso de fecundación y rodea al embrión en los estadios más tempranos del desarrollo. Contrario a lo que dicen muchos textos, no es propia de mamíferos, sino de todos los vertebrados (con disLntos nombres: corion en peces, “membrana perivitelina” en aves). Posee proteínas altamente conservadas (ZP glicoprotreins), presentes en todos los vertebrados Ojo: en marsupiales no hay cavitación de una mórula ni concentración de una masa celular interna Lo indicado en la figura como Blastocele en monotremas es en realidad lo mismo que la cavidad subgerminal entre el embrión y el vitelo. Luego hay una blastulacion secundaria. Lo de placentados tampoco es blastocele: el trofoblasto no es un hipoblasto, el cual tambien se establece despues, de manera secundaria. El trofoblasto permite la implantacion del ovocito en el utero, y no contribuye al embrion. En marsupiales, el embrion siempre es una esfera hueca (protodermo). Una mitad del protodermo da origen al trofoblasto, la otra al embrion. El hipoblasto se forma despues, de manera secundaria. Formación secundaria del hipoblasto en mamiferos. Celulas que delaminan y migran a parLr del epiblasto dan origen a un hipoblasto (tambien llamado exoendodermo) que da lugar a un saco vitelino y no contribuye al embrion. No hay vitelo (yema) y el saco vitelino mas bien conforma parte del cordon umbilical y placenta Hay una desafortunada nomenclatura paralela en mamíferos donde la formación de un hipoblasto secundario ha sido confundido con la gastrulación (proceso que revisaremos mas adelante). El ‘endodermo” que aparece ahi no contribuye al embrion y el “gastrocoele” tampoc es tal (ver más adelante). Sólo el disco superior contribuye al embrión. Placozoa (adulto) Origen de las blástulas: animales basales En todos los animales se observa un proceso de ingreso de células hacia el blastocele: la gastrulación. Larva plánula de Cnidarios Amphiblastula (larva) de Porifera (Esponja) Embrión temprano de Amphioxus Tipos de “movimientos celulares” de gastrulación Mediante el proceso de gastrulación, las células que ingresan conforman un intesLno primiLvo (arquenteron) en forma de tubo interno. El espacio al interior del arquenteron se denomina gastrocele. Durante la gastrulación, el blastocele que exis]a entre epiblasto e hipoblasto es obliterado y desaparece. La capa de células que queda hacia el exterior se llama ectodermo; la capa de células del tubo interno (que da al gastrocele) se le llama endodermo, que da lugar a todas las visceras (excepto urogenitalia) ect ectodermo endodermo endodermo gastrocele Invaginacion en amphioxus Ingresion en un cnidario gastrocele El epiblasto da origen al ectodermo, la capa externa de células que dan lugar a la piel y el sistema nervioso El hipoblasto da origen a estructuras asociadas a la alimentación, el arquenteron en animales basales, y saco vitelino en vertebrados amniotos. Ambos se consideran derivados endodérmicos Las células que ingresan al embrión, además de dar lugar al endodermo dan lugar a una tercera capa de células que se ubica entre el endodermo y el ectodermo: El mesodermo, que da lugar a tejidos conecLvos y musculatura El mesodermo genera cavidades corporales internas, “celomas” conformando una suerte de endoesqueleto hidrostáLco. Este es un rasgo propio de los animales bilaterales (protostomos + deuterostomos) o celomados, ausente en cnidarios y poríferos. En los bilaterales basales, el mesodermo de las cavidades celómicas se origina a parLr de evaginaciones del endodermo, hacia la región dorsal del arquenteron. Esto se denomina como formación enterocélica del celoma. La cavidad del celoma es originalmente conLnua con la del blastocele. Formacion enterocelica del celoma en un erizo En la gastrulación, las células del mesodermo pueden ingresar como un mesénquima laxo de celulas ameboideas altamente móviles. Estas pueden luego condensarse en bloques inicialmente macizos pero que luego desarrollan una cavidad interna, en lo que se conoce como desarrollo esquizocélico de la cavidad del celoma El celoma da origen a la pared muscular del cuerpo y a los revesLmientos musculares de las visceras En algunos bilaterales la organización de cavidades internas se simplifica o pierde parcialmente: Acelomados, Pseudocelomados. Alguna vez se pensó que algunas formas como estas eran basales, hoy sabemos que descienden de verdaderos celomados. Gastrulación en Amphioxus (Chordata) En la región anterior, de manera enterocélica, el techo del arquenteron da origen a los somitos, una serie de bloques mesodermicos pareados (dan lugar a la musculatura axial segmentada), y al notocordo, una barra longitudinal axial de tejido conecLvo y muscular en amphioxus (solo conecLvo en vertebrados). Lo mismo ocurre para los somitos anteriores en ciclostomos (peces sin mandibula) y en peces carLlaginosos (Lburones, rayas y quimeras) La cavidad de los somitos de la región más posterior se forma de manera esquizocelica. En vertebrados óseos, todos los somitos se forman de manera esquizocélica. En la región anterior puede decirse que el endodermo da lugar al mesodermo, y por tanto puede hablarse de un mesendodermo. En cnidarios, hay zonas de endodermo (“estómago” cnidario) que expresan genes de mesodermo y que pueden desarrollarse como musculatura, por lo que se dice que el origen evoluLvo del mesodermo está en el endodermo. Corte transversal Vista superior (dorsal) anterior posterior En el ectodermo dorsal hay una “placa neural” de ectodermo engrosado que por invaginación entra al embrión, conformando un tubo neural, que da lugar al sistema nervioso central. Este proceso se llama “neurulación”. Amphioxus se caracteriza porque ademas dos extensiones epidermicas (no neurales) se encuentran por sobre la placa neural que desciende En el huevo mesoleciLco de la lamprea, no hay una invaginacion completa del hipoblasto como la de amphioxus, hay un labio dorsal del blastoporo en el que ocurre involución e ingresión de celulas. Las celulas que ingresan por el blastoporo convergen hacia el eje medio y en el mismo senLdo, tal que las primeras en ingresar conformará la parte mas anterior (cabeza) y las úlLmas, la parte más posterior. El blastoporo termina siendo el ano. En este proceso el embrión va de forma “esférica” a alargada. La neurulación en lampreas y teleosteos ocurre de manera disLnta a la invaginación del tubo en los demás vertebrados. La placa neural es más angosta y se denomina” quilla” neural. Esta ingresa en el embrión como un cordon macizo y luego genera una cavidad interna (neurocele) por un proceso de cavitacion interna Aspectos morfogené8cos compar8dos por todos los vertebrados En vertebrados en general, el mesodermo puede dividirse en tres zonas: 1) El epímero dorsal, que da origen a los somitos 2) El mesómero, que da origen a riñones y gónadas. 3)El hipómero, que da origen a las paredes musculares del cuerpo y de las vísceras. El hipómero posee un aspecto lateral, la somatopleura, y un aspecto ventral, la esplacnopleura. El epimero y mesomero son segmentados, el hipomero es conLnuo El somito se divide en tres: dermatomo (da origen a la dermis de tejido conecLvo laxo que existe debajo de la epidermis), miotomo (da origen a musculatura de vertebras y extremidades) y esclerotomo (da origen a las vertebras) Ectodermo Mesodermo Endodermo En todos los vertebrados, existen pares de placodas ectodermicas hacia la zona de la cabeza, “islas” de celulas engrosadas que dan origen a neuronas sensoriales de la nariz (placodas olfactorias), del oido (placoda oLca), de la linea lateral (organo mecanoreceptor de peces) y epibranquiales, que contribuyen neuronas sensoriales a los pares de nervios craneales VII, IX, y X En todos los vertebrados se desarrolla ademas una cresta neural. Esta se origina a parLr de bordes laterales engrosados de la placa neural, que acaban en la posicion dorsal del tubo neural. Desde ahi migran hacia el interior del embrion, dando lugar en todo el cuerpo a melanocitos (celulas de pigmentacion) y a los ganglios de los nervios sensoriales del sistema nervioso periférico. En la región cefálica, la cresta neural da origen a la mayoría de los huesos del cráneo. Se ha postulado que la cresta neural y las placodas ectodermicas fueron clave en el origen de los vertebrados, especialmente en relación a la especialización de la cabeza en sensorialidad/alimentación Peces teleósteos presentan un sincicio endodérmico sobre el vitelo (periblasto) que no contribuye al embrión, y un blastodisco en el cual se observa una capa ectodérmica y una capa mesodérmica. En la gastrulación ingresa un “mesendodermo” que da origen a mesodermo y endodermo. A la derecha, la designación del mesendodermo como hipoblasto me parece discuLble, el periblasto me parece análogo al hipoblasto de amniotos, asociado al vitelo, que no contribuye al embrión). Los teleosteos ademas se caracterizan por los extensos movimientos de epibolia por medio del cual la epidermis se exLende desde el disco embrionario por sobre el vitelo de todo el huevo Gastrulación en peces óseos basales (Holosteos) y Anfibios (huevos mesolecíLcos) Gastrulación en Aves y RepLles Hay un surco primiLvo por el cual ingresan celulas que se exLenden hacia anterior y hacia los lados. Las primeras celulas en ingresar hacia anterior conforman la cabeza del embrión. Las células endodérmicas que ingresan desplazan las del hipoblasto hacia los lados. Junto a las celulas del hipoblasto, conforman el saco vitelino. El hipoblasto no contribuye al arquenteron o intesLno primiLvo (primiLve gut) El extremo anterior del surco primiLvo, conforma el “nodo de hensen”, analogo al blastoporo. Este nodo va ‘retrocediendo” hacia posterior “cerrando” el surco promiLvo (recuerda a una cremallera). Las ulLmas celulas en ingresar por el nodo de Hensen son las mas posteriores. E l nodo, como el blastoporo, termina siendo el ano. En mamíferos, debido a sus orígenes a parLr de amniotos ovíparos, y pese a la ausencia de vitelo, hay un embrión completamente discoidal y la gastrulación ocurre igual a un repLl o ave (como sobre una “yema fantasma”) Lo que aparece acá como “gastrulación” es en realidad el establecimiento del hipoblasto secundario. El “gastrocele” no es tal, no se trata de la cavidad del intesLno primiLvo (como en otros vertebrados) sino que del saco vitelino En mamíferos, como en los demas amniotos, las células del hipoblasto secundario pasan al saco vitelino (yolk sac), desplazadas en la gastrulación por las precursoras del intesLno primiLvo. En mamiferos, a diferencia de otros amniotos, se forma desde temprano una nueva capa de celulas y cavidad por sobre el epiblasto del disco embrionario: la cavidad amnioLca. En los demas amniotos, el amnios se forma mas tarde a parLr de la somatopleura del hipomero. Anexos embrionarios: Corion (recubrimiento, respiracion), Amnios (“charca” que rodea al embrion) , Alantoides (desechos, respiracion) , Saco vitelino (alimentacion) Los amniotos presentan reproducción en Lerra por fecundación interna El huevo cleidoico es semipermeable y permite la ovoposición fuera del agua. Existe desarrollo directo sin etapa larval libre. El huevo provee su propio “charco” gracias al amnios (“Asimilación del medio”) Una membrana coriónica de origen ectodérmico, que simplemente recubre el embrión, está presente en todos los vertebrados. NO es exclusiva de amniotos Pez teleósteo En aves y repLles, y en algunos mamíferos el mesodermo somáLco (de la somatopleura) se exLende por encima del embrión generando la cavidad amnióLca. La esplacnopleura genera un diver]culo que junto al endodermo forma el alantoides FIN