Citoplasma-Citosol
Anuncio

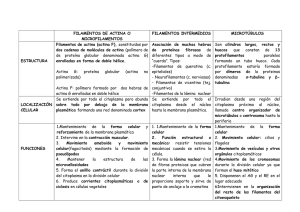
CITOPLASMA – CITOSOL – CITOESQUELETO VÍCTOR HUGO CASCO Citoplasma • El citoplasma comprende todo el volumen de la célula, salvo el núcleo • Está compuesto por el citosol, una solución acuosa concentrada que engloba numerosas estructuras especializadas y orgánulos, y el citoesqueleto, una red de filamentos proteicos que se extienden a través del citoplasma de todas las células eucariotas MATRIZ CITOPLASMÁTICA (CITOSOL) • El citosol es principalmente un medio acuoso (~85% de agua) en la que están disueltas pequeñas sustancias orgánicas: (aminoácidos, glucosa, ATP, etc.) e inorgánicas (iones, sales minerales, etc.) • El citosol tiene una organización muy alta a nivel molecular • Es un fluido de “naturaleza gelatinosa” que contiene ~ 20% de proteínas MATRIZ CITOPLASMÁTICA (CITOSOL) • En él están disueltas muchas de las moléculas que la célula necesita para su metabolismo, entre ellas moléculas orgánicas que son intermediarios del metabolismo. • El citosol está repleto de enzimas que dirigen (catalizan) y controlan el metabolismo celular (glicólisis, gluconeogénesis, vía de las pentosas fosfato, activación de aminoácidos, síntesis de ácidos grasos, síntesis de nucleótidos etc.),. • También contiene sistemas de transducción de señales internas (por ej., segundos mensajeros, AMPc, GMPc, IP3-inositol trifosfato-etc.), • Asimismo se encuentran receptores intracelulares de señalización (Por ej., factores de transcripción, quinasas proteasas, etc.) • En el citosol también se pueden encontrar inclusiones de material de reserva: de lípidos y de glucógeno. Se encuentran los ribosoma libres que realizan la síntesis de proteínas que serán distribuidas y destinadas a diferentes compartimentos celulares (mitocondrias, peroxisomas, núcleo) MATRIZ CITOPLASMÁTICA (CITOSOL) • Las propiedades coloidales de la célula, como las transformaciones básicas de SOL-GEL; de “Sol” (una Solución líquida del citosol más fluida) a “Gel” (un citosol más rígido, sólido y gelatinoso) son básicas para determinadas actividades básicas celulares – como las modificaciones de la viscosidad y el movimiento intracelular del citoplasma en forma de corrientes de fluido citosólico (en inglés: cyclosis o cellular streaming) o – locomoción celular de tipo ameboideo, la formación del huso mitótico y – el clivaje (escisión) de la célula madre para dar lugar dos células hijas durante la fase de citocinesis de la división celular (mitosis), – el crecimiento y formación sinápticas, y – liberación de vesículas de neurotransmisores. MATRIZ CITOPLASMÁTICA (CITOSOL) • Las transformaciones Sol-Gel que pueden ocurrir rápidamente (por ej. 40 ciclos sol-gel por segundo) depende fundamentalmente de los componentes del citosol, y es causado principalmente por las reacciones controladas de ensamblaje y desensamblaje (remodelamiento dinámico) de los elementos del citoesqueleto (embebidos en el citosol) principalmente microfilamentos de Actina y microtúbulos y de las asociaciones contráctiles Actina-Miosina. Inclusiones transitorias • Suspendidos en el seno del citosol existe un número variable de componentes citoplasmáticos que representan acúmulos de nutrientes o subproductos relativamente inertes del metabolismo celular. • Salvo contadas excepciones, no poseen membranas de recubrimiento. • Su concentración en las células depende de la edad de las células, del tipo celular y de su actividad fisiológica. Partículas Glucógeno • Cuando la disponibilidad de glucosa es elevada, la glucógenosintetasa elabora grandes polímeros ramificados que constituyen las moléculas de glucógeno, que es la forma de depósito intracelular de glucosa transitoriamente inmovilizada • Los glucosomas o partículas β son estructuras esferoidales de unos 30 nm, formadas por un centro del polisacárido rodeado por una capa de enzimas que intervienen en su formación y degradación • Los glucosomas pueden mantenerse como estructuras independientes o formar acúmulos (partículas α- o rosetas) • Cuando el contenido es especialmente alto se lo puede visualizar al MO mediante la técnica de PAS Inclusiones Lipídicas • Los acúmulos más comunes son las inclusiones de triglicéridos • Estas formaciones carecen de membrana y poseen tamaños muy diversos • Algunas son visibles sólo con el ME, mientras que otras alcanzan decenas de micrómetros como las gotas citoplasmáticas de los adipocitos que son las células especializadas en la acumulación de lípidos como reserva concentrada de nutrientes para todo el organismo • Pese a que las células utilizan ampliamente el colesterol para la síntesis de sus membranas no existen depósitos visibles de este esterol, excepto las glándulas endocrinas que sintetizan hormonas esteroides Inclusiones cristalinas • La mayoría de las inclusiones cristalinas son proteínas de función y estructura desconocidas, • En algunos casos la composición no es proteica, ejemplo de ello son los cristales intranucleares de hepatocitos caninos que están compuestos de uratos, lo mismo que en las células de la cornea humana de individuos que sufren de gota. • En otros casos la composición proteica de los cristales es bien conocida, por ejemplo, el complejo de apoferritina y hierro (ferritina), que es la forma intracitosólica de depósito del hierro. • En otros casos, la presencia de cristales constituye una adaptación funcional importante como en el tapetum lucidum de animales de hábitos nocturnos. Estos cristales regularmente espaciados, actúan reforzando la sensibilidad de la visión nocturna. Pigmentos • Muchas células poseen granulaciones citoplasmáticas de color pardo, descriptas por los citólogos clásicos como inclusiones de lipofusina o lipocromo. • Estas inclusiones no se presentan en células de vida corta o en organismos juveniles, • Conforme los individuos envejecen su cantidad se incrementa paulatinamente. Ejemplo de ello se da en las células que como las neuronas y los cardiocitos se encuentran terminalmente diferenciados y que han perdido su capacidad de dividirse. • Se tiñen positivamente con la tinción de PAS y débilmente con los colorantes para lípidos. Estos pigmentos representan los estadios finales de la desintegración en vacuolas autofágicas que los años se acumulan en forma de residuos no digeribles, son metabólicamente inertes no son desdoblados por las enzimas lisosómicas. • Otro pigmento que puede observarse en las células es la melanina. Es un pigmento derivado del aminoácido tirosina que se sintetiza en células especializadas llamadas melanocitos. Red de Actina Filamentos intermedios Microtúbulos Célula en movimiento Componentes del Citoesqueleto eucariota El citoesqueleto es exclusivo de Eucariotas? Filamentos helicoidales de la actina bacteriana de Bacillus subtilis (imagen obtenida con microscopio de fluorescencia) R. Carballido-López Citoesqueleto Filamentos de Actina • 7 nm • Actina G Filamentos Intermedios • 10 nm • ~ 50 tipos de Proteínas Microtúbulos • 25 nm • Dímeros de Tubulinas α y β Citoesqueleto: Microfilamentos ¿Actinas? bacterianas Superposición de las estructuras de Actina libre (ADP) y MreB. A pesar Protofilamento del bajo porcentaje de identidad en de actina la secuencia de aminoácidos (~15%) los plegamientos son prácticamente Idénticos. Protofilamento de MreB Filamentos de Actina (~ 7nm) Ensamble y estructura del filamento de actina Extremo (-) 166o G-actina Dímero Trímero Extremo (–) Extremo (+) F-actina Extremo (+) Monómero de Actina 375 aác. 43kd Treadmilling Extremo (-) Extremo (+) Polimerización de la actina Efectos de las proteínas de unión a actina en el recambio del filamento Profilina Cofilina Organización de los filamentos de actina Manojos Redes Filamentos de actina Proteína de entrecruzamiento Filamentos de actina Proteína de entrecruzamiento Formas de organización de la actina y proteínas asociadas Filamentos de actina Filamentos de actina Dominios de unión a Ca2+ Dominio espaciador -helicoidal Fimbrina Dominios de unión a Ca2+ -Actinina Organización de los microfilamentos en las microvellosidades Extremo (-) Membrana Plasmática Filamentos de actina Brazo lateral Miosina l-calmodulina Vilina Fimbrina red terminal Redes de actina y filamina Dominios espaciadores Hoja plegada- Dominio de dimerización Dominio de unión a actina Filamentos de actina Estructura de la espectrina Cadena beta Ca2+ Cadena alfa Dominio de hoja plegada beta Unión de fibras de estrés a la MP en las adhesiones focales Unión de fibras de estrés a la MP en las adhesiones focales Anclaje de FA a uniones adherentes A MP cadherinas Estructura de las células musculares Haz de fibras musculares una fibra muscular (célula muscular) Músculo Núcleo MP Miofibrilla Banda A Disco Z Disco Z Sarcómero Estructura del sarcómero 26.000X Banda A Banda I Zona H actina miosina Disco Z Línea M Sarcómero Esquema de un sarcómero y sus componentes 26.000X Banda A Banda I (Oscura) (Clara) Zona H Actina (filamento delgado) Miosina (filamento grueso) Línea Z Línea M Sarcómero Línea Z Titina y Nebulina Disco Z actina nebulina Miosina Titina (3000kd) Deslizamiento de filamentos Modelo de contracción muscular Disco Z miosina actina Línea M Deslizamiento de los filamentos contracción Miosina II Región de Cabeza globular Cadena liviana esencial Cadena pesada Cadena liviana regulatoria Colas hélices - coiled coil Modelo de acción de miosina La cabeza de miosina se une a una nueva posición de A Disociación del complejo A-M La cabeza de miosina retorna a la posición orginal Hidrólisis de ATP cambio conformacional Desplazamiento de la cabeza de M Desplazamiento de actina Distribución y relaciones de la troponina y tropomiosina musculares Asociación de Tropomiosina y troponina con Filamentos de actina Tropomiosina Troponina Actina Cabeza de miosina Sitio de unión de miosina Tropomiosina Sitio de unión de miosina expuestos Citoesqueleto: Filamentos Intermedios Crescentina ¿FI bacterianos? Filamentos Intermedios ~10 nm Proteínas de filamentos intermedios Tipo Proteína Tamaño (kd) Sitio de Expresión I Queratinas ácidas (~ 15 proteínas) 40-60 Células epiteliales II Queratinas Básicas o neutras (~ 15 proteínas) 50-70 Células epiteliales III Vimentina 54 Desmina Proteína fibrilar ácida gliales Periferina 53 51 Fibroblastos, glóbulos blancos, etc. Células musculares Células gliales 57 Neuronas periféricas IV Proteínas de neurofilamentos Internexina 67-200 66 Neuronas Neuronas V Laminas nucleares 60-75 Lamina nuclear interna VI Nestina 200 Células madre del SNC Estructura de las proteínas de FI Cola Variable en estructura y tamaño N-terminal Cabeza Variable en estructura y tamaño Dominio central en forma de bastón Hélices de 310-350 aác. C-terminal Ensamble de los FI cabeza Polipéptido Coiled coila Dímero Tetrámero Protofilamento Filamento cola Citoesqueleto Microtúbulos ¿Tubulinas procariotas? • Dos tipos de homólogos de tubulina: ejemplificados por FtsZ y las proteínas BtubA and BtubB de Prosthecobacter dejongeii • Las identidades de secuencia en relación a tubulinas eucarioticas son de ~17% para FtsZ y ~35% para BtubA/B • Los análisis filogenéticos sugieren que FtsZ y tubulina habrían evolucionado de un ancestro común y habrían divergido tempranamente mientras que BtubA y BtubB lo habrían hecho mas recientemente siguiendo transferencia horizontal de un precursor eucarionte Organización Celular de FtsZ Microtúbulos ~25 nm Tipos de MT Microtúbulos citoplasmáticos Microtúbulos del axonema • Mantenimiento de los axones • Mantenimiento de la polaridad durante la migración • Orientación del depósito de las miofibrillas de celulosa • Formación del huso mitótico y meiótico • Disposición espacial y movimiento de organelas y vesículas • Cilias • Flagelos • Corpúsculos basales Dímero de tubulinas α y β Diagrama de cintas basado en un modelo de una resolución de 3.7 Angstrom desarrollado por científicos del laboratorio de Berkeley, que muestra que la tubulina es un heterodímero que consiste de dos monómeros de estructuras casi idénticas (alfa y beta). Cada monómero está formado por un núcleo de dos láminas beta (verde), rodeado por hélices α (azul), y cada uno se une a un nucleótido guanina (rosa). Además de un sitio de unión a un nucleótido, cada monómero tiene otros dos sitios de unión, uno para proteínas y el otro para la droga anti-cancerosa taxol Estructura de los microtúbulos 1 2 13 3 12 11 4 10 5 6 7 Tubulina- 8 9 Tubulina- Estructura de los microtúbulos Tinción Negativa Tinción Positiva Citoesqueleto basado en microtúbulos Inestabilidad dinámica de los microtúbulos Alta concentración de tubulina unida a GTP Baja concentración de tubulina unida a GTP Inhibición de la polimerización por colchicina colchicina Proteínas motoras de microtúbulos (+) (–) Dominios cabeza Coiled – coil Cadenas liviana e intermedia base Dineína cola Cadena liviana Quinesina Transporte de vesículas a lo largo de los microtúbulos quinesina Núcleo centrosoma dineína Microtúbulos polares Movimiento de los cromosomas en la Anafase A Microtúbulos Astrales Microtúbulos Quinetocóricos Proteína motora direccionada hacia el extremo (-) Desensamble de MT cromosoma Separación de MT polares en anafase tipo B Microtúbulos astrales Motor direccionador al extremo (–) Microtúbulos polares Microtúbulos cinetocóricos Motor direccionador al extremo + Motor direccionador al extremo (–) Cilias y flagelos cilio cilio Protofilamento Microtúbulo protofilamentos Doblete Tiplete Centríolo Doblete ciliar Triplete centriolar Corte transversal de un cilio Fibra externa (doblete) Brazo externo Membrana Brazo interno Puente de Nexina Fibra central Cabeza del Ligamiento Radial Ligamiento Radial Vaina Central Cilios y Cuerpos basales a b Movimiento de microtúbulos en cílias y flagelos Dineína (+) Movimiento de la cabeza de Dineína hacia el extremo (-) del tubo B (–) Ligamiento de nexina curvamiento de microtúbulo Centríolos Los cuerpos basales y centríolos se replican autónomamente