T11 protegido
Anuncio

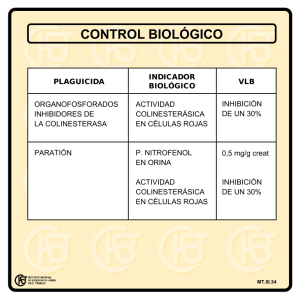
INHIBIDORES • Moléculas que disminuyen la V de reacción enzimática Afectan a la cinética de reacción al interferir con: INHIBICIÓN ENZIMÁTICA - La formación del complejo ES - Con su disociación TEMA 11 - Con ambos procesos a la vez • Existen varios mecanismos. Veremos los efectos que producen sobre Enzimas de cinética M-M TIPOS DE INHIBICIÓN Inhibiciones reversibles Inhibiciones irreversibles 1.- INHIBICIÓN COMPETITIVA 2.- INHIBICIÓN NO COMPETITIVA 3.- INHIBICIÓN ACOMPETITIVA Ó INCOMPETITIVA 4.- INHIBICIÓN POR SUSTRATO • Unión no covalente del I, no altera la estructura del E. Al desaparecer, se recupera la actividad catalítica • Unión covalente del I, altera la estructura del E de forma definitiva •Tipos 1; 2; 3 1 Inhibición Reversible (IR) La IR puede afectar a Vmax ó Km ó a ambas a la vez 1.- INHIBICIÓN COMPETITIVA. Si sólo afecta Km IR Competitiva • El I compite directamente con un sustrato por un sitio de fijación enzimático ( centro activo del E) Si sólo afecta Vmax IR No-Competitiva • Se parece al S ( forma y tamaño similares = isóstero) Si afecta tanto a Km y a Vmax IR Acompetitiva IR Mixta • No es reactivo ( si lo fuera sería un S alternativo competitivo) 1.- INHIBICIÓN COMPETITIVA. Modelo General A. Compitiendo con el sustrato por el sitio de unión al centro activo S Enzima E +S ES E+P + S I Ki I Enzima no activa Ki ´ EI +S No hay reacción 2 B. Modificando la conformación del centro activo El grado de inhibición competitiva depende de la fracción de E que se una al I de modo que impida la unión del sustrato K1 E +S K2 + K3 E+P I S Ki ´ Ki Enzima No hay reacción I se une de forma reversible al E y está rápidamente en equilibrio con ella EI es catalíticamente inactivo I EI S Enzima no activa KI = • La v de formación EI será igual a la v de descomposición [E] [I] en el KI = Ki= [I] [E]= Ki´[EI] KI= Ki´/ Ki equilibrio [EI] • Puede ser superada a >> [S] • El I competitivo ↑ valor aparente de Km : para obtener la misma v de reacción se necesita > [S] α=1+ [I] es la concentración de inhibidor KI es la cte de disociación para el complejo EI Km ap es la Km observada en presencia de I [E] [I] [EI] +S KI es la cte de disociación o cte de equilibrio de fijación de I a E Inhibidor competitivo reduce la concentración de E libre disponible para la unión del sustrato Ecuación de Michaelis- Menten para reacción inhibida competitivamente cte de disociación • Inhibición mas potente cuanto < KI Km ap = Km (1 + [I]/KI) ES Se deduce como antes (M-M) pero con un término adicional que representa la fracción de [E]T que une a I para formar EI [E]T = [E] + [ES] + [EI] [I] KI V0 = Vmáx [S] α Km + [S] α=1+ resulta [I] KI α Km = Kmap 3 1.- INHIBICIÓN COMPETITIVA. Reacción de M-M V0versus [S] en presencia de diferentes concentraciones de I Valores crecientes de α 9 α , una función de concentración del I y su afinidad por el E no puede ser < 1 9 Presencia de I hace que [S] parezca menor de lo que es en realidad. Hace que Km parezca mayor de lo que es en realidad como consecuencia de que la unión de I y S al E es mutuamente excluyente 9 El aumento de [S] puede superar al I competitivo α es el factor por el que [S] debe aumentar para superar el efecto de la presencia del I V0 ~ Vmáx para cualquier valor de α (cualquier concentración del I) 9 El I no afecta el nº de recambio del E 9 Cuando [S] ~ ∞ ; Gráfico Lineweaver-Burk KI puede medirse Vmax= Kcat [ET] Vmáx [S] V0 = 1 = V0 Transformación en doble recíproca α Km + [S] α Km 1 Vmáx [S] + 1 α= 1 + Vmáx [I] KI Corresponde a gráfico de ecuación de una recta: - Pendiente es α Km / Vmáx - Intersección en el eje 1/[S] es – 1 / α Km - Intersección en el eje 1/V 0 es 1/Vmáx 4 Ejemplos -OOC-CH 2-CH2 -COO- succinatoDH Succinato succinatoDH Malonato -OOC-CH 2-COO -OOC-CH=CH-COO- Fumarato SIN REACCIÓN - Ibuprofeno inhibidor competitivo de la ciclooxigenasa, interviene en la respuesta inflamatoria Lehninger. 4ª Ed Intoxicación por metanol Estatinas inhiben enzima clave de la biosíntesis de colesterol 2.- INHIBICIÓN NO COMPETITIVA. ADH • No existe competitividad entre S y el I • I no tiene relación estructural con S ceguera • I se une al E en sitio diferente al que lo hace el S ( en un lugar no activo, diferente al centro activo del E) • I no impide la unión del S. Pudiéndose unir simultáneamente al E tanto el S como el I ADH • I inactiva el E tanto si el S está presente como si no = distorsiona el sitio activo del E reduciendo su poder catalítico 5 Inhibidores reversibles Inhibición no competitiva. Esquema de reacción K1 2. No competitivos Se puede unir tanto a la enzima libre como al complejo enzima-sustrato E + S K2 + I ES Enzima I I EI EI I S K´I ESI No hay reacción I El I puede unirse al E libre EIS E + P + I KI S K3 ES El I puede unirse al complejo ES E+I ES + I EI ESI Enzima no activa Inhibición no competitiva. Esquema de reacción • Vmáx disminuye en proporción a un factor α = 1 + [I] / K´I • Km permanece inalterable porque el E se une a S con la misma afinidad • K´I es la cte de equilibrio de disociación del complejo ESI 6 Inhibición no competitiva. Esquema de reacción Inhibición no competitiva. Esquema de reacción • Representación de las inversas para obtener la ecuación de Lineweaver-Burk, se transformará en: a) Considerando el caso mas sencillo en que KI = K´I (se disocian a igual velocidad EI y ESI) • Aplicando las mismas consideraciones que en la Inhib. Competitiva [I] 1 1 1+ = KI V0 Vmáx [E]T = [E] + [ES] + [EI] + [ESI] • Expresión de la Ecuación de M-M como: V0 = Vmáx [S] [I] [I] + 1+ 1+ KI KI Km + [I] [S] [I] α (Km + S) [I] = 0, Pendiente = Km / Vmáx - 1 / Km [S] [I] KI 2.- INHIBICIÓN NO COMPETITIVA MIXTA. b) Considerando el caso en que KI ≠ K´I (se tendrá que considerar las dos ctes de disociación) • La Km se verá afectada por α y α´ 1 / Vmáx Vmáx ap 1 KI • La Vmáx, se modifica, disminuye (la E menos eficiente con I) Vmáx ap KI [I] • El punto de corte es el mismo en presencia o ausencia de I, lo que justifica que no se modifica la Km Vmáx [S] Sin I 1+ • La recta corta al eje de las abcisas en su lado negativo KI [I] creciente, Pendiente = Km / Vmáx 1 + 1/v 1 / Vmáx 1 + = [I] α=1+ + Km Vmáx α=1+ 1/[S] • Para calcular Vmáx se debe obtener el punto de corte de la recta al eje 1/V. • Al aumentar [I] aumenta el valor de 1 / Vmáx 1 + ya que es el inverso de Vmáx [I] α´ = 1 + KI [I] K´I • La ecuación de M-M: [I] KI V= Vmáx [S] α Km + α´ S = Vmáx [S] α α´ Km + S 7 2.- INHIBICIÓN NO COMPETITIVA MIXTA. 2.- INHIBICIÓN NO COMPETITIVA MIXTA. • La ecuación de las inversas en: Con I 1/V 1 V = α Km 1 Vmáx S α´ + Vmáx 1 [I] 1+ K´I Vmáx • Representación gráfica daría un conjunto de rectas que se cortan al lado izdo del eje 1/V = α´ [I] creciente Pendiente = Km / Vmáx .α Sin I Pendiente = Km / Vmáx Vmáx 1/Vmáx • Se modifican las Vmáx y las Km • Se denomina mixta porque se modifican las dos • Probablemente el I se una a sitios relacionados con la catálisis (modificandola) y repercusión en la fijación del S a ese lugar - α´ α Km 1/[S] Se denomina Mixta porque se modifican Vmáx y Km Si KI< K’I Lehninger. 4ª Ed 8 3.- INHIBICIÓN ACOMPETITIVA Ó INCOMPETITIVA. • El I se fija directamente al complejo ES, pero no a la E libre • La etapa de fijación de I presenta una cte de disociación K´I K1 E + S • Se modifica Km y Vmáx K2 • Se forma un compuesto terminal ESI K1 E + S K2 E + P K´I ESI K3 ES K3 ES + I E + P K´I = + I [ES] [I] [ESI] ESI No hay reacción K´I [ES] + [I] Ki= [ES][I]=Ki´[ESI] K´I No hay reacción Km = [ESI] [E] [S] [ES] [E] = Km [ES] [S] Ki´/Ki = KI´ Deducción de la Ecuación M-M • El I, que no tiene que parecerse al S provoca una distorsión del sitio activo del E que conduce a una inactivación del mismo • Se fija en sitio ≠ de S • La presencia de I altera tanto la Vmáx y la Km [ET] = [E] + [ES] + [ESI] [ET] = Km [ES] [S] [ES][I] + [ES] + [ET] = [ES] Km + 1 + [I] K´I [S] [ES] = [ET] Km [I] + 1 + [S] K´I v = K3 . [ES] Sacando factor común [ES] K´I Despejando [ES] (a) Sustituyendo en (a) 9 v = Vmáx = Km + 1 + [I] [S] K´I K3 E T Km + 1 + [I] [S] K´I = Vmáx . [S] Km + [S] 1+ [I] Vmáx . [S] v = K´I α´ 1 v v = Vmáx . [S] 1 Km + α´ [S] v 1 v = = = Expresando en forma de inversos: Km + α´[S] Km + α´ [S] Vmáx . [S] Km Vmáx . [S] + α´ [S] Vmáx . [S] 1 1 α´ Km + Vmáx Vmáx [S] La pendiente con I es igual que sin I: Km/Vmáx Vmáx ap = Kap = Vmáx [I] 1 + K´ I Km 1 + [I] K´I Para las tres rectas -1/Km 10 Resumen de los distintos tipos de inhibiciones con enzimas de comportamiento cinético M-M Resumen de los distintos tipos de inhibiciones con enzimas de comportamiento cinético M-M Tipo de inhibición Kmap Ninguna Vmáx Competitiva Vmáx No competitiva (mixta) Vmáx 1 + [I] K´I Km . 1 + [I] KI =α 1 + [I] Km KI 1 + [I] = α´ K´I Vmáx 1 + [I] K´I Km 1 + [I] K´I Incompetitiva 4.- INHIBICIÓN POR SUSTRATO. Vmáxap Km =α 4.- INHIBICIÓN POR SUSTRATO. • 2 Hipótesis: Las E pueden inhibirse por [S] altas Representación de la cinética en forma acampanada Representación linealizada ó de dobles recíprocos no dá una línea recta V 1ª. E puede disponer de 2 lugares de unión para el mismo S: - el sitio catalítico y - el de inhibición 2ª. E adopte 2 configuraciones diferentes reversibles entre sí. : 1/V -activa (Ea) - inactiva (Ei) [S] 1/[S] 11 4.- INHIBICIÓN POR SUSTRATO. 2ª Hipotesis El S se une indistintamente a una u a otra forma. Solo se transformará en P cuando está unido a Ea Ej: catálisis de la hexoquinasa de cerebro hexoquinasa Glucosa-6-fosfato + ADP P + Ea Ea + <S Ei + >S EaS EiS • Si el E está libre en citoplasma ⇒ inhibida por uno de los sustratos (G-6-P) • Si el E está unido a la pared externa de mitocondria ⇒ no inhibida • Inhibidores de la DHFR. Antifolatos INHIBIDORES ENZIMÁTICOS COMO AGENTES QUIMIOTERAPEUTICOS • ATP + Glucosa ametopterina METOTREXATO/ AMINOPTERINA NH2 COOH N Como análogos estructurales del S bloquean la acción enzimática específica N H 2N Ej: Metotrexato (ametopterina), agente anticanceroso. Inhibidor competitivo parecido químicamente al dihidrofolato (DHF CONH N R N COOH N hasta 10000 veces mayor afinidad por la enzima NH2 OCH3 N TRIMETOPRIM DHF reductasa DHF THF H2N Ác. timidilico (dTMP) Bacteriostáticos.inhiben la sintesis de acido folico bacteriano OCH3 N OCH3 DNA OCH3 NH2 TRIMETREXATO R=H H2N CH3 N R N N OCH3 OCH3 12 INHIBICIÓN REVERSIBLE EN ENZIMAS BISUSTRATO • E bisustrato, posee 2 sitios de fijación de S distintos • Mecanismo de inhibición mas complejo • En E reversibles bisustrato se producen ≠ situaciones - Inhib. Competitiva - Inhib. Acompetitiva - Inhibición no competitiva a) E, S1 y S2 b) Fijación ordenada. - S1 y S2 pueden fijarse aleatoriamente ó al azar. - I para uno de los sustratos (ej. para S2) + S1 + + I Solo se une a ES1 S1 ES1 S2 I + S2 Inhibición acompetitiva Inhibición competitiva 13 c) forma ordenada I se puede unir al E directamente ó al complejo ES1 INHIBIDORES IRREVERSIBLES • Bloquean o destruyen definitivamente el E + + I S1 • Enlace covalente entre el E y el I irreversible EI + + S1 • Su disociación es muy lenta ( ligazón muy fuerte) I ES1 Inhibición no competitiva INHIBIDORES IRREVERSIBLES 1º Grupo: Unión al centro activo con bloqueo irreversible del E • Clasificación Ejemplos: -1º grupo: Unión al centro activo con bloqueo irreversible del E • Acetil colinesterasa • Gases neurotóxicos (sarin ) e insecticidas (malatión) -2º grupo: Análogos del estado de transición Acetilcolina - 3º grupo: Inhibidores suicidas Cuadro de estimulación permanente Parálisis muscular Tetania 14 ACh + enzima (Acetilcolinesterasa) ------> Colina + Acetilcolinesterasa acetilada 2º Grupo. Análogos del estado de transición • El I debe parecerse al estado de transición del S Acetilcolinesterasa acetilada + H2O ------> Acetilcolinesterasa + ácido acético O O2N O - O2N C H3C N+ CH3 O C O Ser CH CH2 H 3C N+ CH3 O OH + - O- O O2N O HO P O C H3C F N+ CH3 Productos CH3 H3C N+ CH3 CH3 Ser CH CH2 H3 C O Análogo de estado de transición H3C OH Estado de transición CH3 O O2N Compuestos organofosforados Sustrato CH3 O- O • Ej: DFP ó DIFP = diisopropil fluorofosfato - actúan como inhibidores competitivos CH3 CH O P O H3C CH CH3 P O CH H3 C CH3 DIFP: diisopropil fluorofosfato CH3 CH - Actúan sobre enzimas serínicas (Acetilcolinesterasa) - Únicamente sobre la Ser activa - Insecticidas Organofosfóricos: Parathion, Malathion - Gases neurotóxicos (sarin) 15 diisopropil fluorofosfato Inhibición enzimatica irreversible 3º Grupo. Inhibidores suicidas = Inactivadores basados en el mecanismo de reacción enzimática • Poco reactivos hasta que se unen al sitio activo del E Modo de acción de los inhibidores suicidas E+I 1 EI 2 EI* 3 E’ + I* • Utilizan el m. de reacción enzimática para inactivar el E • Papel central en el diseño de nuevos fármacos y • Pocos efectos secundarios. Tienen por tanto: a) la especificidad del inhibidor competitivo y b) la potencia de los inhibidores irreversibles 1. El inhibidor se fija a la enzima igual que el sustrato o un inhibidor competitivo convencional 2. La acción catalítica de la enzima convierte al inhibidor I en una especie altamente reactiva I* 3. I* modifica covalentemente a la enzima, inactivándola de forma definitiva al igual que un inhibidor irreversible. 16 Penicilina (activa) Ejemplos de inhibidores suicidas(1) S R CO NH - Sistema de la β-lactamasa bacteriana CH3 CH3 N O La utilización masiva de antibióticos b-lactámicos (penicilinas, sus derivados semisintéticos y cefalosporinas) ha conducido a la aparición de resistencias a los mismos. Los microorganismos resistentes a estos antibióticos lo son por producir una enzima, la b-lactamasa, que inactiva a los antibióticos β-lactámicos. COO- β-Lactamasa R CO NH O C HN O- S CH3 CH3 COO- Ác.peniciloico (inactivo) Muy a menudo los preparados de penicilinas o penicilinas semisintéticas se formulan añadiendo un inhibidor suicida de la β -lactamasa, el ácido clavulánico O CH2OH C N O β-Lactamasa H O O C O CH2OH C HN H COO- Ác.clavulánico CH2OH C HN H - Sistema de la monoamino oxidasa (MAO) cerebral encargadas de la degradación de ciertos neurotransmisores O C O- COO- CH CH2 Ser O Ejemplos de inhibidores suicidas(2) Esta molécula reacciona con la serina activa de la β-lactamasa, produciendo su inactivación La inhibición de la monoamino oxidasa se emplea como terapéutica de los estados depresivos. Se han desarrollado muchos inhibidores suicidas de la MAO - COO 17 18