clase nora 2
Anuncio

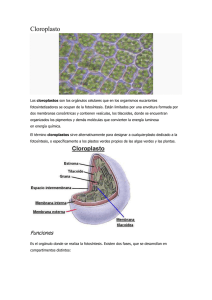
27/08/2015 NÚCLEO NÚCLEO EUCARIÓTICO TÍPICO Núcleo mesocarionte • Cromosomas permanentemente condesados • ADN circular • No hay histonas • Mitosis sin huso http://www.botany.ubc.ca/Biol320/ultra/nucl03.htm 1 27/08/2015 Estructura interna MITOCONDRIAS matriz crestas Membr. interna Espacio intermembranas Membr. externa Origen endosimbiótico a partir de algo semejante a Daptobacter o Bdellovibrio Tipos de crestas mitocondriales en la Naturaleza CRESTAS MITOCONDRIALES chatas Discoides saco tubulares ramificadas burbuja Burbuja-Chrysolepidomnas Tubular-Actinomonas Chatas-Hemiselmis 2 27/08/2015 PEROXISOMAS • Organela donde se produce la transformación del glicolato (proveniente de la fotorespiración) en glicina (que luego pasará a glicerato), mediante la glicolato oxidasa • La glicolato oxidasa aparece en Glaucophyta, Rhodophyta, Phaeophyceae, Charophyceae y Plantae FOTOSINTETIZAR…. CON QUÉ? • • En las otras algas no interviene el peroxisoma y actúa la glicolato dehidrogenasa en mitocondrias) No contiene ácidos nucleicos CYANOPHYTA (incluyendo Prochlorophyta) Los tilacoides varían en estructura y ordenamiento según la especie. También la cantidad (según condiciones de crecimiento, la especie y el tipo celular (células vegetativas, heterocistos, acinetas): Las membranas tilacoidales en Prochloron (“Prochlorophyta”) están organizadas en regiones apiladas y no apiladas, que recuerdan los grana de las planta superiores. a. 3-6 capas paralelas periféricas, forman una red anastomosada de vainas concéntricas (cromoplasma) separada del centroplasma (ej. Synechococcus planctibus y Anabaena sp.) b. perpendiculares a la pared celular (ej. Oscillatoria y Arthrospira) c. ordenamiento radial (ej. Phormidium retzi) 3 27/08/2015 Las ficobilinas son pigmentos accesorios para el funcionamiento del fotosistema II (también existen en Glaucophyta, Cryptophyta y Rhodophyta). Synechococcus. 3b y c: adaptaciones cromáticas a luz verde y azul Los ficobilisomas son tres núcleos cilíndricos de cuatro moléculas de aloficocianina, próximos a la membrana del tilacoide, sobre la que convergen conjuntos de moléculas hexaméricas en forma de bastones de ficocianina sola o de ficocianina y ficoeritrina Int J Biol Sci 2007; 3:434-445 • Si hay apilamiento, este solo puede darse en regiones limitadas. http://mmbr.asm.org/cgi/content/full/73/2/249/F5 CLOROPLASTOS: • Durante la división celular los tilacoides deben separarse y dividirse en las células hijas. • Proteínas, lípidos, carotenoides y clorofila a son los principales componentes de los tilacoides. Organelas que contienen los pigmentos para absorber la luz y canalizar la energía de las moléculas de pigmento excitadas en una serie de reacciones fotoquímicas y enzimáticas 4 27/08/2015 • En eucariotas, los tilacoides, libres o apilados, están encerrados por membranas formando cloroplastos • Los cloroplastos contienen ácidos nucleicos y ribosomas • El ADN no asociado a proteínas • Aparece como pequeños nucleoides dispersos o anillos periféricos • Los pigmentos se organizan en complejos proteicos en la membrana de los tilacoides (~24 nm de espesor, con lumen ~ 10 nm de ancho). • Replican su propio ADN , no relacionado con la división del plasto. • Autoreplicación de los cloroplastos: coordinada o no con la de la célula. Usualmente solo provienen del lado materno en la reproducción sexual • Las superficies de todos los tilacoides corren paralelas al plano mayor del cloroplasto. • Los genes plastidiales se transcriben y traducen dentro de los plastos. • Síntesis de ARN y proteínas puede ocurrir dentro del plasto pero no es estrictamente autónoma del genoma nuclear Forma y número de extremadamente variable: cloroplastos • cupuliforme (Chlamydomonas) • La maquinaria para la síntesis proteica está parcialmente compuesta por productos genéticos importados desde el núcleo (⇒no son totalmente independientes) • acintado y espiralado (Spirogyra) • estrellado (Zygnema) • Discoides y (Acetabularia) numerosos (ca. 108) • ~1/2 genoma plastidial son genes que contribuyen a la maquinaria de la expresión génica, por ejemplo, genes para rARN, tARN, subunidades de ARN polimerasa y proteínas ribosomales. 5 27/08/2015 Origen endosimbiótico UN POCO DE HISTORIA En 1883, Andreas Schimper fue el primero en estudiar y describir la naturaleza potencialmente endosimbiótica de las células. En 1905, Konstantin Mereschkowsky fue el que tuvo la idea de que los plastos se habrían originado por endosimbiosis. Esencialmente él fue el primero en formular la idea de la simbiogénesis (la simbiosis sería la principal forzante de la evolución). En 1926, en colaboración con Ivan Wallin, aplicó esta idea a las mitocondrias, en su libro “Symbiogenesis and the Origin of Species.” Wallin fue realmente el primero en sugerir que la célula eucariota estaría compuesta por microorganismos Esta idea fue el motor de la posteriormente llamada Hipótesis Endosimbiótica. En 1927, él también explicó cómo las bacterias podrían representar la causa primaria del origen de las especies A. Primaria B. Secundaria C. Terciaria Protista engulle un alga con plasto 2io Protista engulle un alga eucariota Protista engulle una cianobacteria nucleomorfo Transferencia de genes nucleomorfo Pérdida de nucleomorfo Estas teorías fueron rechazadas en su momento debido a la creencia de que mitocondrias y cloroplastos carecían de ADN La presencia de ADN recién se probó en los 60’s. http://endosymbiotichypothesis.wordpress.com/history-the-formation-of-the-endosymbiotic-hypothesis/ Alga 2 membranas 3-4 membranas 3 membranas Origen de los plastos vía endosimbiosis A: primaria, B: secundaria y C: terciaria. 6 27/08/2015 Origen endosimbiótico ENDOSIMBIOSIS SECUNDARIA ENDOSIMBIOSIS PRIMARIA Transferencia de genes Cyanobacteria Alga roja Mecanismo de Endosimbiosis 2ia Alga con cloroplasto primario Vacuola alimenticia 1er huésped Nucleomorfo Proteobacteria Cryptophyta 20 huésped Eucariota heterótrofo Vacuola digestiva Diatomea HN: núcleo del huésped; EN núcleo del endosimbionte ORGANIZACIÓN DE LOS PIGMENTOS EN LOS DISTINTOS GRUPOS 7 27/08/2015 GLAUCOPHYTA • Las cianelas funcionan como cloroplastos. Cada una, rodeada por una pared reducida de péptidoglicano (excepto en Glaucosphaera sp.), está encerrada en una vesícula dentro del citoplasma del huésped. •Las cianelas no fijan nitrógeno, contienen gránulos de polifosfato y un carboxisoma conspicuo en el centro, semejante al pirenoide de otras algas. • Los tilacoides no están apilados sino que son solitarios y equidistantes en un ordenamiento concéntrico • Contienen clorofila a y ß -caroteno, que es el principal carotenoide • Los ficobilisomas intertilacoidales contienen aloficocianina y ficocianina. No hay ficoeritrina aunque algunas especies presentan ficoeritrocianina. RHODOPHYTA • Los cloroplastos tienen doble membrana y no se relacionan con el retículo endoplasmático. • En Floridophyceae el número, forma y posición de los plastos (muchos, discoides y parietales) es bastante uniforme, puede o no haber pirenoides • Algunas unicelulares con un plasto estrellado, como en Phorphyridium. • Puede haber una lamela periférica • Tilacoides normalmente paralelos entre sí aislados, • Hay clorofila a; ß-caroteno y luteína. • El ADN forma numerosos nucleoides dispersos. • Tienen ficobilisomas en su superficie, con aloficocianina, ficocianina y 5 formas de ficoeritrina 8 27/08/2015 CHLOROPHYTA • Doble membrana (~Rhodophyta y Glaucophyta) • Gran variación en forma y tamaño: axiales o parietales, cupuliformes, laminares, reticulados, estrellados, etc. • Tilacoides de a 2-6 o más, interconectados (~ grana de plantas) • No hay lamela periférica • Uno o más pirenoides, a menudo atravesados por tilacoides • ADN en pequeños nucleoides dispersos en la matriz • Clorofilas a y b; ß-caroteno y otros carotenoides siempre presentes (luteína, zeaxantina, violaxantina) OCHROPHYTA (=HETEROKONTOPHYTA) 9 27/08/2015 • Cloroplasto rodeado por 4 membranas, con las dos externas como parte del REC; continuo con la carioteca si los plastos están cerca del núcleo. • Los tilacoides se agrupan de a 3 (2 en Raphidophyceae) • Los tilacoides de lamelas adyacentes suelen interconectarse •ADN: anillo entre la lamela periférica y las otras (excepto en Eustigmatophyceae, donde forma nuceoides puntiformes) • En todos los casos hay una lamela periférica, excepto en Chattonella (Raphidophyceae) y Eustigmatophyceae. Nannochlopsis sp Pigmentos principales: • Clorofila a (Eustigmatophyceae) • Clorofila a + c1 y c2 (Chrysophyceae, Rhaphidophyceae, Phaeophyceae, Dictyocophyceae y Xanthophyceae) • Clorofila a + c2 (Bacillariophyceae). Otros pigmentos accesorios: • ß-caroteno y • xantofilas (diadinoxantina, heteroxantina, vaucheriaxantina, anteeraxantina y luteina). Las especies marinas y de agua dulce de Raphidophyceae difieren en sus pigmentos accesorios: El pigmento accesorio más importante: • Fucoxantina (Chrysophyceae, Bacillariophyceae, Dictyocophyceae y Phaeophyceae) • Violaxantina • Vaucherioxantina (Eustigmatophyceae) marinas agua dulce Fucoxantina Violaxantina Diadinoxantina Heteroxantina Vaucheriaxantina (Xanthophyceae). 10 27/08/2015 •Puede haber uno o más plastos laminares (Chrysophyceae y Eustigmatophyceae), discoides (Xanthophyceae, Raphidophyceae y Dictyochophyceae) o acintados (Phaeophyceae), que pueden ser parietales •En Bacillariophyceae el número y la forma tienen importancia para diferenciar géneros. •Pueden estar o no en estrecha conexión con el núcleo • En todas las clases hay pirenoides, excepto en las zoosporas de Eustigmatophyceae y las especies de agua dulce de Raphidophyceae. • Son de tipo semi-inmerso, unidos a la cara interna del cloroplasto, en forma de pera en Phaeophyceae o pedunculados en Eustigmatophyceae. • Puede haber uno o muchos PRYMNESIOPHYTA (=HAPTOPHYTA) Generalmente hay uno o dos cloroplastos por célula, con tilacoides agrupados de a 3 Están rodeados por el reticulo endoplasmático confluyendo con la carioteca (el núcleo está siempre cerca de los plastos) No hay lamela periférica Pirenoides en forma de huso están comunmente inmersos en el cloroplasto, penetrados por 1-3 pares de tilacoides pero en algunos géneros pueden protruir. Contienen clorofilas a, c1 y c2 y, en algunos géneros también c3; ß-caroteno y xantofilas (la más común es la fucoxantina) El ADN forma numerosos nucleoides dispersos 11 27/08/2015 DINOPHYTA CRYPTOPHYTA Hay 1-2 por célula, rodeados por 4 membranas • Puede o no haber cloroplastos, dependiendo del tipo de alimentación Entre los dos pares hay un espacio que contienen ribosomas 80S, gránulos de almidón y el nucleomorfo • Cuando están presentes, están cubiertos por 3 membranas no conectadas al RE Los tilacoides están de a pares (aunque se han visto solitarios y apilados en números mayores) y no hay lamela periférica • Los tilacoides generalmente se agrupan de a 3 y no suele haber lamela periféricoa El pirenoide se proyecta desde el interior del plasto • Hay varios tipos de pirenoides, pedunculados o embebidos en el cloroplasto Los pigmentos son clorofilas a y c2; ficoeritrina, ficocianina y aloficocianina; carotenos y varias xantofilas. Estas ficobilinas son diferentes de las de las cianobacterias y las algas rojas porque tienen menor peso molecular y no forman ficobilisomas; además están ubicadas en el lumen tilacoidal. • Hay clorofila a y c2, b-caroteno y varias xantofilas, entre ellas peridinina • El ADN forma numerosos nódulos dispersos Esta descripción corresponde al tipo de plasto más común, el tipo peridinial El ADN forma numerosos nucleoides dispersos. Alrededor del 50% de los dinoflagelados adquieren sus plástidos por endosimbiosis secundarias con una variedad de eucariotas fotosintéticos (como Amphidinium carterae) mientras que otros lo hacen por endosimbiosis terciaria (como Dinophysis, que tiene un plasto con 2 membranas derivado de una criptofita); Dinoflagelado con diatomea endosimbionte Karenia sp y otros tienen fucoxantina como pigmento accesorio y los plastos derivan de haptofitas, con 2 o 4 membranas Karenia mikimotoi Endosimbionte Peridinium quinquecorne Kryptoperidinium sp. y taxones relacionados que tienen un plasto con 5 membranas y un núcleo (derivado de una diatomea) M: membrana EN núcleo EM: mitocondria C: cloroplasto Huésped DN núcleo E: mancha ocular G: golgi s: almidón 12 27/08/2015 Origen del plasto-diatomea • Un dinoflagelado, con un plasto peridinial típico y estigma característico (C), adquiere una Nitzschia como endosimbionte (hace menos de 20-25 MA, Oligoceno/Mioceno) • El cloroplasto original queda reducido a una mancha ocular de otro tipo (B) • Análisis moleculares indican que las especies con este “plasto” son monofiléticas • En una sola de estas spp, el plasto deriva de un Chaetoceros, que supuestamente habría reemplazado a la Nitzschia original (endosimbiosis seriada) Endosimbiosis secundaria serial: captura de un nuevo endosimbionte con plasto. KLEPTOPLASTIDIA • Ej. Lepidodinium sp. y taxones afines Amphidinium latum • El plasto peridinico original es reemplazado por un plasto 2io, con 2 membranas, probablemente derivado de una prasinofita (Chlorophyta) Amphidinium acidotum latum y Gymnodinium Endosimbiosis con una criptofita, adquirida como presa y retenida por un largo periodo • Hay escamas en la superficie del cuerpo (~Prasinophyceae), atípicas en esta División, lo que sugiere que se expresan genes adquiridos del endosimbionte • En Noctiluca sp. también se halló una Prasinophyceae como endosimbionte. Lepidodinium viride 13 27/08/2015 Al igual que en Dinophyta los cloroplastos tienen 3 membranas. EUGLENOPHYTA En general hay muchos por célula y son diversos tanto en tamaño, forma y morfología. 5 tipos principales 1. Discoides sin pirenoide (Phacus) 2. Elongados o peltados con un pirenoide central desnudo (Trachelomonas) 3. Grandes y laminares con pirenoides lenticulares con una placa de paramilon a cada lado (Euglena obtusa) 4. Laminares con un gran pirenoide que se proyecta desde el interior, cubierto por una copa cilíndrica o esférica de paramilon (Colacium) 5. Cloroplastos acintados que radian de 1-3 centros de paramilon (Eutreptia) 14 27/08/2015 • Nunca están conectados con el núcleo vía RE • Los tilacoides normalmente se agrupan de a 3 • No hay lamela periférica CHLORARACHNIOPHYTA • Hay 5-7 plastos bilobulados en posición periférica • Están rodeados membranas que separadas o con chatas entre ellos, por un sistema de pueden parecer 4 un par con vesiculas o como 3 membranas • Siempre hay 4 membranas separadas cerca del extremo proximal del pirenoide y sobre él • Los pigmentos fotosintéticos son clorofilas a y be; carotenos y xantofilas • El par externo, cuando hay 4, es del tipo REC pero el par interno se interpreta como la membrana del cloroplasto • El ADN forma pequeños gránulos dispersos en todo el estroma HISTORIA DE LA EVOLUCIÓN ALGAL • Los tilacoides están a menudo laxamente apilados de a 3, sin lamela periférica • Cada cloroplasto porta un pirenoide piriforma central que se proyecta notablemente. Alrededor de él, a menudo hay vesículas con ß-1,3-glucano que es el principal carbohidrato de reserva. • Un nucleomorfo con ADN y un cuerpo tipo nucleolo están presentes entre la 2ª y la 3ª envolturas de cada plasto, en una depresión en la superficie del tilacoide • Pigmentos: clorofilas a y b y xantofilas Flechas: orígenes de los linajes eucarióticos clave, determinados por el registro fósil (A, 1- 4) o datos moleculares (B-D). (A a D) Algas: A, archaeplastidios; B, dinoflagelados; C, haptofitas; D, diatomeas. (0 y 2 -4). Plantas: 0, plantas terrestres primitivas; 2, vasculares; 3, coniferas; 4, plantas con flores. Fig. basada en datos del proyecto ENSEMBLE de H. Griffiths, University of Cambridge 15