- Ninguna Categoria
Mecanismos de modulación por atención selectiva entre áreas
Anuncio
Mecanismos de modulación por atención selectiva
entre ár eas corticales
Un modelo biofísico de red
Joan Salvador Ardid Ramírez
Memoria del trabajo de investigación para la
obtención del D.E.A.
Índice
Capítulo 1: Introducción
1
1.1 Campos receptivos y curvas de sintonía
3
1.2 Atención
4
1.2.1 Atención Voluntaria y Memoria de Trabajo
1.3 Atención basada en Característica
1.3.1 Atención basada en Característica Global
1.3.2 Atención basada en Característica Local
1.4 Modelo computacional
Capítulo 2: Métodos
5
8
8
11
13
15
2.1 El modelo de red
16
2.2 Estímulos visuales y tareas de atención
21
2.3 Materiales usados
26
Capítulo 3: Resultados
3.1 Resultados con RDPs simples
29
30
3.1.1 Cruce de las curvas de sintonía de la población
31
3.1.2 Efecto multiplicativo de las curvas de sintonía neuronales
32
3.2 Resultados con RDPs transparentes
35
3.2.1 Modulación por atención con RDPs transparentes
38
3.2.2 Modelo de Competición Sesgada
40
3.3 Mecanismos
41
Capítulo 4: Discusión
45
Referencias
51
Capítulo 1: Introducción
El sistema visual es el sistema sensorial más ampliamente estudiado históricamente, y
por lo tanto no es casual que el análisis de la atención se haya abordado especialmente
desde la perspectiva de la percepción visual. Se sabe que lo que se percibe depende
críticamente de dónde se dirige la atención. Así, la atención a un determinado lugar
aumenta la precisión y la velocidad de detección de cualquier estímulo visual. Se ha
demostrado también que la atención no sólo aumenta la sensibilidad ante la percepción
de estímulos, sino que a su vez disminuye la interferencia provocada por estímulos
distractores cercanos. Además, la atención es altamente flexible, en cuanto a su foco (un
determinado lugar en el espacio, una determinada dimensión del estímulo) y a su
dinámica (cambio rápido y preciso de foco de atención), para la correcta consecución de
la finalidad buscada en cada momento.
Pero, ¿cómo se producen todos estos efectos en el cerebro? ¿cuáles son las áreas
involucradas y los mecanismos responsables? Aún se está lejos de obtener respuestas
definitivas a estas preguntas, pero ya son muchos los trabajos que se han venido
publicando recientemente sobre cómo la atención influye en la representación de los
estímulos visuales en el cerebro. Algunas investigaciones muestran que, ante condiciones
de estimulación visual idénticas, la actividad de las neuronas de la corteza visual depende
de sobre qué zona del campo visual se encuentra localizada la atención. Otras
investigaciones muestran que la atención sobre una determinada región visual afecta a la
actividad de neuronas que cubren todas las demás regiones del campo visual. Y otras
más, muestran que la región del espacio visual en la cual estímulos adecuados generan la
activación de una determinada neurona, su campo receptivo, se ve influenciada por
medio de la atención.
Así, la mayor parte de estos estudios de atención visual pueden ser clasificados
(Boynton, 2005), o bien como de Atención Espacial, o sea, es determinante la posición en
el espacio que el estímulo visual atendido ocupa como fuente de la modulación de la
actividad neuronal (Motter 1993; Treue y Martinez-Trujillo, 1999; Reynolds et al., 2000);
o bien como Atención basada en Característica, en inglés Feature-based Attention, que
afecta a todas las neuronas selectivas a la dimensión atendida del estímulo
independientemente de la posición de su campo receptivo con relación al foco de
atención. La característica hace mención al valor concreto que toma una determinada
dimensión del estímulo: por ejemplo, si la dimensión del estímulo es el color, posibles
Mecanismos de modulación por atención selectiva entre áreas corticales: Un modelo biofísico de red
características serían rojo, azul, verde...; otras posibles dimensiones de estímulo son
textura, movimiento, forma... Para poner de manifiesto esta segunda modalidad de
atención, típicamente más débil que la espacial, se realizan experimentos en los que los
sujetos prestan atención a objetos siempre en la misma zona de la escena visual, pero
con distinta característica. Al comparar condiciones en las que la posición de la atención
no varía, sino sólo la característica atendida, las modulaciones en la respuesta neuronal
que se observan son achacables a la Atención basada en Característica.
La Atención basada en Característica, a su vez, se ha venido dividiendo en otras dos,
según la atención que modula la actividad de la neurona esté dentro del campo receptivo,
Atención basada en Característica Local, o fuera, Atención basada en Característica
Global. Si está dentro, la atención afecta al propio campo receptivo (Moran y Desimone,
1985; Desimone y Duncan, 1995), estrechándolo más al estímulo atendido, en cambio si
está fuera la modulación afecta principalmente a la actividad neuronal (Treue y MartinezTrujillo, 1999; Saenz et al., 2002; Martinez-Trujillo y Treue, 2004).
De manera que, en estos últimos años, ha aparecido cierta controversia entre los
científicos que defienden cada una de estas visiones sobre atención visual, puesto que
son aproximaciones aparentemente contradictorias entre ellas. En este estudio nos vamos
a centrar en la Atención basada en Característica.
Esta división entre Atención Espacial y Atención basada en Característica, de hecho, no
es tan clara en la literatura como Boynton refleja. Así pues, algunos científicos, como
Treue, consideran que una determinada localización espacial es otra característica más
del estímulo, y otros, como Desimone, realizan experimentos en los que la modulación
depende también de aspectos espaciales, no únicamente de la característica. No
obstante, para los aspectos que en este trabajo se van a tratar, la clasificación dada por
Boynton es una aproximación muy buena ya que no se van a considerar aspectos de
localización: o no existen porque los estímulos están solapados, o estos serán muy
pequeños por estar separados pero dentro de un mismo campo receptivo.
Históricamente han aparecido dos teorías que se centran en cada una de las ramas de
la Atención basada en Característica, son el Modelo de Competición Sesgada, o en inglés
Biased Competition Model, para la Atención basada en Característica Local; y el Modelo de
Ganancia dependiente de la Similitud respecto a Característica, o en inglés Featuresimilarity Gain Model, para la Atención basada en Característica Global.
El principal objetivo de este estudio es implementar un modelo computacional,
plausible desde el punto de vista biológico, que sustente mecanísticamente y a la vez
unifique de forma consistente las dos posturas enfrentadas.
Para ello, empezaremos describiendo cómo se mide la actividad neuronal frente a un
determinado estímulo visual. Se introducirán las definiciones de campo receptivo y curva
de sintonía. Antes de entrar a considerar la modulación de la actividad, nos convendrá
introducir el concepto de atención: ¿qué es lo que entendemos por atención? ¿qué
relación mantiene con la memoria de trabajo? ¿dónde se genera? ¿dónde se aplica? ¿qué
la sustenta?, etc. Posteriormente, veremos ya cómo es la modulación que la atención
genera en cada una de las dos situaciones, Atención basada en Característica Local y
Atención basada en Característica Global, y a partir de ellas, qué defiende cada modelo.
Por último, en esta introducción se comentará brevemente cuál es la aproximación de
nuestro modelo computacional, cómo se relaciona con el contexto de atención visual, en
concreto la Atención basada en Característica, y por qué se ha escogido un modelo de
estas características.
2
Capítulo 1: Introducción
1.1 Campos receptivos y curvas de sintonía
La actividad neuronal se mide a partir de la tasa de disparo, o sea el número de
potenciales de acción (PAs) por unidad de tiempo. Se dice que una neurona está activa
cuando recibe un estímulo que hace cambiar su tasa de disparo respecto de la tasa de
disparo media en ausencia de estímulo, que llamaremos Actividad Espontánea. No
obstante, no todos los estímulos son válidos. Cada neurona codifica una, o varias
dimensiones concretas del estímulo (dirección, orientación, color, forma...) (figura 1.1).
Si el estímulo en cuestión no lleva ese tipo de información, la neurona no se activará.
Además de esta especialización en la respuesta de las neuronas (que llamamos
selectividad al estímulo), también se da una especialización a nivel de áreas cerebrales.
Las neuronas de diferentes áreas del cerebro codifican un tipo u otro de información
característica presente en el estímulo: así en área visual extraestriada V4 del macaco
encontramos neuronas selectivas a la orientación, la frecuencia espacial y el color,
mientras que las neuronas del área visual MT no responden ante esas dimensiones del
estímulo y en cambio son altamente selectivas a la dirección y magnitud del movimiento.
Además, para que la neurona reciba el estímulo adecuado, éste ha de estar situado en
una determinada región del espacio. Típicamente una neurona de la corteza visual no
abarca todo el campo visual sino una sola región. El rango total se reparte entre
diferentes neuronas de la misma área. Cada una de estas regiones del espacio reciben el
nombre de campo receptivo, receptive field (RF) en inglés (figura 1.1). De manera que
cada neurona de la corteza visual tiene su propio campo receptivo específico, localizado
de forma fija en coordenadas retinotópicas.
Pero este campo receptivo que abarcan las neuronas varía en tamaño según el área en
que se encuentren. De hecho existe una jerarquía entre diferentes áreas de una misma
vía, o sea, a medida que la información sensorial avanza a través de diferentes áreas
sensoriales, aumenta el procesamiento de ésta información sensorial y a su vez el tamaño
promedio del campo receptivo de las neuronas (Felleman y Van Essen, 1991).
Además, se sabe que diferentes neuronas en una misma área comparten el mismo
campo receptivo. Estas neuronas que codifican una misma dimensión del estímulo,
difieren entre sí por su nivel de actividad ante los valores particulares que toma la
dimensión del estímulo en cuestión. Cada valor particular es una característica diferente
del estímulo. De manera que cada neurona tendrá una característica preferida y su tasa
de disparo variará en función de cómo la dimensión del estímulo se acerque a ella. Este
es el concepto de curva de sintonía, o tuning curve en inglés (figura 1.2).
Figura 1.1: Dimensiones del estímulo y campo receptivo. Estímulo, en este caso visual, con varias
dimensiones: forma rectangular, color azul, y según el caso, cambio de orientación (izquierda), o de
dirección de movimiento (derecha). Cada valor particular de una determinada dimensión del estímulo es
una característica (azul, orientación a 135o, movimiento hacia arriba,...). La linea discontinua de color
negro representa el campo receptivo de una hipotética neurona.
3
Mecanismos de modulación por atención selectiva entre áreas corticales: Un modelo biofísico de red
Así, por ejemplo, para un estímulo que se desplaza en una determinada dirección,
tendremos que las neuronas selectivas a dirección, que se concentran principalmente en
el área temporal medial, área MT, se activan cuando el estímulo entra en su campo
receptivo, y que un subconjunto de las que tienen el mismo campo receptivo están más
fuertemente activas, puesto que son éstas las que prefieren la dirección en la que se
mueve el estímulo.
Feature
Figura 1.2: Curva de sintonía. Existe una relación entre el valor que toma la dimensión del estímulo en
cuestión, característica, y la respuesta de la neurona que a ella es selectiva. El pico de la curva ocurre
para la característica preferida para esta neurona, y variará de unas a otras.
1.2 Atención
No existe una definición completamente unificada del concepto de atención y, además,
ahora se sabe que probablemente existen diversos circuitos interrelacionados formando
un complejo sistema de atención (Posner y Petersen, 1990). Así nos encontramos que
existen matices entre las diferentes definiciones existentes según las distintas
aproximaciones al problema, o sea, según el campo de trabajo de donde provengan, o
también según el contexto histórico en que surgieron (extraído de Raz, 2004):
● 'La atención es el afinamiento de los sentidos'. Aristoteles, 370 AC.
● 'La atención actúa como un filtro, dada la alta cantidad de información existente
localizada en sistemas sensoriales paralelos y la limitada capacidad de percepción'.
Broadbent, 1958.
● 'Atención como mecanismo para la preparación de la respuesta motora'. Rizzolatti et
al., 1987.
● 'La atención deriva de la influencia de la memoria a corto plazo, o memoria de
trabajo, sobre representaciones sensoriales en la corteza cerebral'. Desimone, 1998.
Una definición reciente y bastante general es la siguiente:
● 'La atención es la habilidad para seleccionar, de entre otros muchos, los estímulos,
acciones, recuerdos y pensamientos que son relevantes desde el punto de vista del
comportamiento'. Raz, 2004.
4
Capítulo 1: Introducción
Como ya se ha dicho, la atención no implica un único mecanismo, sino que se trata de
un sistema complejo que reside sobre un número de circuitos neuronales distintos. Para
poder abordar cuáles son los mecanismos neuronales que la sustentan, desde el punto de
vista anatómico y fisiológico, se necesitan unas estrategias muy bien definidas basadas
en la creación de determinados paradigmas, o tareas, en los que se puedan separar las
señales relacionadas con aspectos de atención de otras puramente sensoriales o motoras.
Además, por estar distribuido en diferentes zonas del cerebro, se ha necesitado de
nuevos avances técnicos, desarrollados a lo largo de los últimos veinte años, y que han
permitido medir de forma no invasiva la actividad de todo, o gran parte del cerebro
durante el comportamiento. Gracias a ello, se está estudiando cuáles son las áreas que se
activan en tiempo real según el paradigma usado. Existen diferentes herramientas de
imagen de este tipo como: ERP (Event-Related Potential), fMRI (functional Magnetic
Resonance
Imaging),
MEG
(Magneto-Encephalography),
NIRS
(Near-InfraRed
Spectroscopy), PET (Positron Emission Tomography), SPECT (Single Photon Emission
Computed Tomography). Las principales conclusiones, respecto al sistema de atención, a
las que se ha llegado con esta nueva tecnología se pueden resumir en tres enunciados
(Posner, 2004):
● Las
redes neuronales involucradas en tareas cognitivas, y emocionales, se
distribuyen ampliamente en diferentes áreas del cerebro, pero no lo ocupan todo.
Cada área de la red global procesa diferentes aspectos, aunque es conjuntamente
como se realiza la tarea.
● Algunas redes controlan a otras, así existen redes neuronales relacionadas con la
atención que seleccionan y controlan redes sensoriales y de memoria.
● Estas redes cambian con el desarrollo, el aprendizaje, las lesiones cerebrales y las
patologías.
De hecho, efectivamente, las últimas investigaciones han demostrado que distintas
áreas del cerebro median diferentes procesamientos de la atención, por tanto es posible
examinar la atención selectiva como un sistema con su propia función, anatomía,
estructura celular y conectividad en el cerebro (Posner y Fan, 2004).
Conviene conjugar los resultados de este tipo de estudios en paralelo con los registros
electrofisiológicos, que aportan mayores detalles a nivel de una o pocas neuronas. No
obstante, la noción sobre atención que se tiene ahora, gracias a estas nuevas técnicas, es
mucho más amplia, pues han permitido analizar aspectos de la atención que antes no se
podía, como la comparación de la actividad entre áreas corticales según la tarea a realizar
o el estado cognitivo del sujeto. Y aún más, se han podido realizar estas medidas en
cerebro humano, al tratarse de instrumentos no invasivos. Compaginar todos estos tipos
de medidas junto con la implementación de modelos computacionales parece ser la mejor
aproximación para estudiar los mecanismos neuronales que sustentan la atención, dadas
las complejas características de este sistema.
1.2.1 Atención Voluntaria y Memoria de Trabajo
Acabamos de ver que la atención es un sistema distribuido y complejo que está
interrelacionado, entre otros aspectos, con la memoria. Vamos a ver ahora que existe un
tipo de atención, la Atención Voluntaria o Dirigida (Directing Attention), que está
fuertemente ligada con la Memoria de Trabajo (en inglés Working Memory, WM), lo cual
ha originado la idea de que comparten mecanismos neuronales (Mesulam, 1981;
Mesulam, 1990; Desimone y Duncan, 1995; Desimone, 1998; Awh y Jonides, 2001).
5
Mecanismos de modulación por atención selectiva entre áreas corticales: Un modelo biofísico de red
Como se verá posteriormente, estos conceptos serán muy útiles para la implementación
de un modelo computacional biológicamente plausible de la Atención basada en
Característica.
La Atención Voluntaria, también llamada Atención Endógena, hace referencia al tipo de
atención en el que el sujeto busca una determinada finalidad durante el proceso, como
por ejemplo, encontrar una cara familiar en un recinto deportivo. En oposición a otro tipo
de atención llamado Atención Refleja, o Atención Exógena, en la cual, un cambio súbito
por sí mismo en un estímulo sensorial, previamente no atendido, es capaz de atraer
nuestra atención de forma inconsciente. Mediante esta disgregación entre Atención
Voluntaria y Atención Refleja la evolución ha optimizado el sistema visual de los primates.
Se sabe que existen alrededor de un millón y medio de axones que salen de la retina
aportando una riqueza inmensa de detalles de la escena visual. No obstante, en todo
momento existe multitud de información irrelevante. La forma con la que se ha resuelto
el control del flujo de la información visual ha sido mediante la focalización de la atención
en la zona central de la retina, la cual está altamente desarrollada, junto con la habilidad
de realizar movimientos rápidos y precisos de los ojos de forma refleja hacia alguna parte
periférica que se mueve, por ejemplo, de forma abrupta (Yantis y Johnson, 1990;
Remington et al., 1992). Este es un aspecto clave y necesario desde el punto de vista de
la supervivencia. Esta habilidad para cambiar rápidamente y de forma refleja también
aparece en otras modalidades como el sistema auditorio, por ejemplo cuando en una
conversación a la que no prestamos atención, de repente oímos nuestro nombre. El que
seamos capaces de mantener la concentración en una determinada tarea dependerá del
grado de Atención Voluntaria que le apliquemos frente a la aparición de estímulos
periféricos que puedan aparecer en el entorno en principio irrelevante. De hecho se ha
demostrado la falta de habilidad para detectar grandes cambios de la escena visual
cuando estos ocurren fuera del foco de atención (Rensink et al., 1997).
La WM se refiere al proceso activo, mediante el cual se mantiene algún tipo de
información relevante en el cerebro durante un breve periodo de tiempo, pasado éste, la
memoria a corto plazo se libera; como por ejemplo cuando mantenemos en nuestra
'mente' un número de teléfono que se ha dicho previamente hasta el momento en que lo
anotamos.
Tanto la Atención Voluntaria, como la WM, se consideran 'Top-down' ('de arriba a
abajo'), pues son procesos que buscan una determinada finalidad y dependen de
conocimiento previo y no de estímulos sensoriales. Estos procesos se contraponen a los
procesos 'Bottom-up' ('de abajo a arriba'), que están conducidos por el estímulo sensorial
y, si es el caso, por la Atención Refleja. Esta nomenclatura, a grandes rasgos, deriva del
hecho de que la atención proviene de áreas corticales jerárquicamente superiores y el
estímulo sensorial de las áreas corticales primarias.
Utilizando las herramientas de imagen que se han enunciado más arriba, juntamente
con estudios electrofisiológicos de neurona única y estudios de pacientes con lesiones
cerebrales, se ha observado que las estructuras de control para atención difieren de
acuerdo con el tipo de procesamiento, Atención Voluntaria o Atención Refleja, mediante
circuitos corticales separados en las regiones de la corteza cerebral frontal y parietal (ver
citas más adelante). A la vez que se ha visto que existe superposición entre los sustratos
neuronales que controlan la WM con los que controlan la Atención Voluntaria (ver citas
más adelante), lo cual apoya la idea que los mecanismos que sustentan una y otra son
los mismos (Desimone, 1998).
De forma cronológica, lo primero que se empezó a estudiar fue la WM en monos,
donde se puso de relieve la importancia de la corteza prefrontal. Así, lesiones en la
6
Capítulo 1: Introducción
corteza prefrontal dorsolateral impiden en gran medida la aparición de WM (Goldman y
Rosvold, 1970; Bauer y Fuster, 1976; Funahashi et al., 1993). A la vez, registros
extracelulares de neurona única han demostrado que las neuronas en esta región
muestran actividad sostenida en tareas en las que se requiere recordar alguna
característica de un estímulo cuando éste ya está ausente (Fuster y Alexander, 1971;
Kubota y Niki, 1971). Además, en ensayos en los que los monos fallan en la tarea a
realizar, se ha observado que la actividad sostenida se pierde (Fuster 1973; Rosenkilde et
al., 1981; Watanabe, 1986; Funahashi et al., 1989). Esta actividad sostenida ha sido
interpretada como el correlato neuronal de la WM (Fuster, 2001). De todos modos, la
actividad sostenida también se ha observado en otras áreas dependiendo del tipo de
estímulo o de la tarea a realizar, como la corteza temporal inferior (Fuster y Jervey,
1982; Chelazzi et al., 1998), la corteza parietal (Gnadt y Andersen 1988; Chafee y
Goldman-Rakic, 1998; Chafee y Goldman-Rakic, 2000), o la corteza premotora (Bruce y
Goldberg, 1985).
En humanos, con técnicas de imagen, se han obtenido idénticos resultados de
actividad sostenida. En el área prefrontal (Cohen et al., 1997; Courtney et al., 1997), en
el área temporal inferior (Courtney et al., 1998), en el área parietal (D'Esposito et al.,
1998; Jonides et al., 1998; Rowe et al., 2000), y en el área premotora (Courtney et al.,
1998; Petit et al., 1998). En concreto, en las áreas: corteza prefronal dorsolateral
(DLPFC), campos oculares frontales (FEF), campos oculares suplementarios (SEF), lóbulo
parietal superior (SPL), y en el surco intraparietal (IPS) (Pessoa et al., 2002b) (figuras
1.3 y 1.4 B). Al igual que se vio con registros electrofisiológicos, también con técnicas de
imagen se observa que la región DLPFC, no sólo participa en la WM, sino que también es
un indicador del comportamiento del sujeto (Pessoa et al., 2002b).
Figura 1.3: Áreas activas durante tareas de WM. Las regiones activas son el resultado de la comparación
entre ensayos de WM y ensayos control de visión directa al punto de fijación (Pessoa et al., 2002b).
Abreviaciones: DLPFC, dorsolateral prefrontal cortex; DO, dorsal occipital; FEF, frontal eye field; IPS,
intraparietal sulcus; ITG, inferior temporal gyrus; P. MFG, posterior middle frontal gyrus; SPL, superior
parietal lobule.
En cuanto a la Atención Voluntaria, ésta se manifiesta justo en las mismas zonas
dorsales (SPL, FEF, SEF) (Kastner y Ungerleider, 2000; Pessoa et al., 2002a) (figura 1.4
C versus B), mientras la Atención Refleja se manifiesta en zonas más ventrales como la
circunvolución frontal medial (MFG), la corteza cingulada anterior (ACC -Anterior
Cingulate Cortex), y la circunvolución frontal inferior (IFG -Inferior Frontal Gyrus)
(Corbetta y Shulman, 2002; Pessoa y Ungerleider, 2004). Un sitio donde ocurre un
solapamiento entre los dos tipos de atención es la zona anterior IPS, lo cual se ha
7
Mecanismos de modulación por atención selectiva entre áreas corticales: Un modelo biofísico de red
interpretado como que podría resultar clave para la dinámica de la atención, o sea,
cuando un estímulo periférico pasa a ser el foco de atención (Pessoa y Ungerleider,
2004).
Figura 1.4: Regiones involucradas en atención visual y WM. A. Actividad cortical en tareas WM. B. Áreas
activas durante la WM, contraste entre la fase sin estimulación respecto al caso con estimulación. C.
Regiones activas durante tareas de atención visual. Obtenido a partir del análisis de diferentes
experimentos previos: (1) Corbetta et al., 1993; (2) Fink et al., 1997; (3) Nobre et al., 1997; (4)
Vandenbergh et al., 1997; (5) Corbetta et al., 1998; (6) Kastner et al., 1999; (7) Rosen et al., 1999; (8)
Corbetta et al., 2000; (9) Hopfinger et al., 2000. Extraído de Pessoa y Ungerleider, 2004.
1.3 Atención basada en Característica
Como se ha comentado anteriormente, la Atención basada en Característica hace
referencia a las modulaciones de la actividad neural observadas experimentalmente en
áreas sensoriales, debidas a que la atención apunta al valor que posee una determinada
dimensión del estímulo visual. Cuando esto ocurre, aparece un aumento de la actividad
de las neuronas que responden preferentemente a esa característica (Motter 1994;
Duncan y Nimmo-Smith, 1996; Beauchamp et al., 1997; O'Craven et al., 1997; Reynolds
y Desimone, 1997; Valdes-Sosa et al., 1998). Este tipo de efectos se ha observado a lo
largo de las áreas V1, V2, V4, IT (corteza temporal inferior - InferoTemporal cortex), en
la parte posterior de la corteza temporal inferior, TEO, y en el área temporal medial MT
(Middle Temporal).
No obstante, tradicionalmente, la Atención basada en Característica se ha dividido
según la atención que modula la actividad esté dentro o fuera del campo receptivo que se
está registrando. Así se consideran la Atención basada en Característica Local y la
Atención basada en Característica Global, respectivamente. Vamos ahora a describir cada
una por separado junto con los modelos teóricos asociados.
1.3.1 Atención basada en Característica Global
Treue propone el Modelo de Ganancia dependiente de la Similitud respecto a
Característica en el que la regulación de la ganancia de una neurona sensorial refleja la
similitud entre las características del estímulo atendido y las características sensoriales
8
Capítulo 1: Introducción
preferidas por la neurona (forma, color, movimiento, textura...). Esta modulación se da,
tanto si el estímulo atendido está dentro, como si está fuera del campo receptivo (Treue y
Martinez-Trujillo, 1999), y por esta razón hablaríamos de Atención basada en
Característica Global.
Vamos a describir los experimentos de Treue donde se ponen de relieve este tipo de
modulaciones en el área visual MT, selectiva a dirección de movimiento (Martinez-Trujillo
y Treue, 2004). En ellos, la atención permanece siempre fuera del campo receptivo de la
neurona que se está registrando. Hay dos diferentes paradigmas en este tipo de
experimentos.
En el primero (figura 1.5 A) se mantiene la atención sobre estímulos en movimiento,
por tanto, hay siempre modulación de la respuesta sensorial. Lo que cambia entre
diferentes ensayos es la relación entre la dirección de movimiento del estímulo sensorial,
la del estímulo atendido y la dirección preferida de la neurona. Lo que se observa es que
hay un efecto multiplicativo entre el caso de atención sobre la dirección preferida de la
neurona respecto al caso de atención sobre la dirección opuesta, o nula. Y esto ocurre
para las dos posibles direcciones del estímulo sensorial, preferida o nula. Es decir, si la
atención está focalizada sobre la característica preferida de la neurona, ésta multiplica su
respuesta a cualquier estímulo, sea o no preferido, por un factor mayor que uno.
Probablemente este resultado sea extrapolable para todos los estímulos sensoriales
posibles, obteniendo así un efecto multiplicativo en las respectivas curvas de sintonía,
cuando comparamos atención sobre la dirección preferida versus nula.
Figura 1.5: Diseño experimental para el estudio de la Atención basada en Característica Global (MartinezTrujillo 2004). A. Se comparan los casos correspondientes a las dos filas, esto es cuando cambia la
dirección a la que se atiende, en la primera fila la atención recae sobre la dirección preferida y en la
segunda sobre la dirección nula. B. En este caso la comparación es entre el caso de dirección atendida
idéntica a la dirección sensorial respecto caso de atención sobre otra dimensión del estímulo (el punto de
fijación ocular), se puede considerar no atención porqué no se relaciona con ningún tipo de movimiento.
Es importante notar que en cualquiera de los diseños experimentales descritos en esta figura, la atención
esta siempre localizada fuera del campo receptivo de modo que la atención espacial no puede ser
responsable de los efectos que se observen.
9
Mecanismos de modulación por atención selectiva entre áreas corticales: Un modelo biofísico de red
En el segundo (figura 1.5 B) se compara el caso en que la atención recae sobre un
estímulo en movimiento, idéntico al estímulo sensorial pero fuera del campo receptivo,
respecto al caso en que la atención recae sobre otra dimensión en un estímulo fijo
(normalmente el color del punto de fijación ocular), caso que se puede considerar sin
atención en lo que respecta a las respuestas de MT, que son sólo selectivas a la
dimensión movimiento. En este caso lo que se observa es un cruce entre las curvas de
respuesta en los dos casos diferentes (figura 1.6 A).
Figura 1.6: Ejemplo de las curvas de respuesta para una neurona típica en el diseño experimental de la
figura 1.5 B (Martinez-Trujillo 2004). A. Se observa el cruce de las curvas de respuest entre los casos de
atención sobre un estímulo idéntico al estímulo sensorial respecto al caso de atención sobre el punto de
fijación ocular. B. Relación punto a punto entre las dos curvas. Estos factores se pueden considerar como
factores multiplicativos fruto de la modulación por atención. Y su variación se puede interpretar como que
la modulación aumenta o disminuye de forma multiplicativa en función de la separación de ésta con la
dirección preferida de la neurona.
Este resultado y el anterior se pueden conciliar en un mismo mecanismo multiplicativo
si se interpreta que éste aparece cuando la dirección del estímulo atendido permanece fija
a medida que se modifica el estímulo sensorial en el campo receptivo. En cambio, si ésta
no existe o varía manteniendo fija la separación con la dirección del estímulo sensorial lo
que ocurrirá será que en cada punto de la curva de respuesta habrá un factor
multiplicativo diferente, resultando que la curva con atención se cruza respecto a la otra
(figura 1.6 B) (Martinez-Trujillo y Treue, 2004; Boynton, 2005). Esta misma tendencia se
ha observado en una población (N=135) (Martinez-Trujillo y Treue, 2004).
10
Capítulo 1: Introducción
Análogas modulaciones por atención se han observado mediante técnicas de imagen de
resonancia magnética nuclear funcional (fMRI) en humanos (Saenz et al., 2002; Kamitani
y Tong 2005) y en otros estudios fisiológicos (Motter 1993; McAdams y Maunsell, 1999).
1.3.2 Atención basada en Característica Local
Desimone propone el Modelo de Competición Sesgada que se trata de un modelo que
se aplica a la atención selectiva, con la premisa de que cualquier efecto que provoca la
atención en las respuestas neuronales se entiende mejor en un contexto competitivo
entre diferentes estímulos (Moran y Desimone, 1985; Desimone et al., 1990; Desimone y
Duncan 1995; Desimone 1996; Duncan 1996; Luck et al., 1997; Chelazzi et al., 1998).
Este modelo se basa en cinco aspectos clave (Desimone, 1998):
● Si
dos estímulos se presentan simultáneamente dentro del campo visual,
inicialmente estos activan sus representaciones neuronales en paralelo. No obstante,
si los dos estímulos son independientes y la misma región localizada de la corteza
sensorial recibe a ambos, los dos estímulos entrarán en competencia y las
respuestas neuronales en promedio se verán mutuamente suprimidas.
● Esta supresión aumentará aún más cuando los dos estímulos caen dentro del mismo
campo receptivo.
● Estas interacciones competitivas pueden ser sesgadas hacia uno u otro estímulo en
función de donde recaiga la atención, a través de señales tanto 'Bottom-up' como
'Top-down'.
● El sesgo no está limitado únicamente a la localización de los estímulos en el campo
visual, también influyen otros aspectos relevantes como el color, la textura, la
forma, etc.
● La principal fuente del sesgo 'Top-down' deriva de estructuras de la corteza
prefrontal que generan actividad persistente, o sea, memoria de trabajo.
La mayor parte de las bases del modelo provienen de estudios con tareas de búsqueda
visual en la vía visual ventral (áreas V2, V4 e IT de macacos) (Chelazzi et al., 1993;
Chelazzi y Desimone, 1994; Chelazzi et al., 1998; Reynolds et al., 1999) (figura 1.7).
La figura 1.7 (a) muestra el diagrama de la tarea utilizada en la que el mono ha de
elegir entre dos estímulos diferentes en función de las instrucciones previas. Hay tres
fases: 'Cue' donde aparece el estímulo a elegir; 'Delay' es un periodo de latencia o pausa;
y 'Target' es la fase final en la que el mono ha de dirigir la mirada al mismo estímulo
aparecido en 'Cue'. Durante las fases 'Cue' y 'Delay' el mono mantiene la mirada fija
sobre un punto de fijación que aparece en pantalla. Las figuras 1.7 (b) y (c) muestran las
respuestas promediadas de una población de neuronas obtenidas con este tipo de tareas.
Para cada neurona hay uno de los estímulos que se considera preferido y otro
antipreferido en función de la respuesta puramente sensorial que evocan, como se ve en
la fase 'Cue' de la tarea. Durante la fase 'Delay' la respuesta es alta sólo si la atención
recae sobre el estímulo preferido. En la fase 'Target' las respuestas promediadas
empiezan igual, señalando que los estímulos activan en paralelo diferentes
representaciones corticales. Pero posteriormente éstas divergen en función del estímulo
atendido, si el estímulo atendido es el preferido, entonces la respuesta permanece alta
durante todo el ensayo, no así para el antipreferido.
11
Mecanismos de modulación por atención selectiva entre áreas corticales: Un modelo biofísico de red
Figura 1.7: Respuestas de neuronas en tareas de búsqueda visual (Desimone, 1998). (a) Representación
esquemática de la tarea. Hay tres fases, A: 'Cue' donde aparece el estímulo a elegir; B, 'Delay' es un
periodo de latencia o pausa; y C, 'Target' es la fase final en la que el mono ha de dirigir la mirada al
mismo estímulo aparecido en 'Cue'. Durante las fases 'Cue' y 'Delay' el mono mantiene la mirada fija
sobre un punto de fijación que aparece en pantalla. Para cada neurona hay uno de los estímulos que se
considera preferido y otro antipreferido en función de la respuesta puramente sensorial que evocan. Hay
determinados ensayos en los cuales ninguno de los estímulos en la fase 'Target' coincide con el estímulo
previo de la fase 'Cue' , en ese caso el mono ha de mantener la mirada en el punto de fijación. (b)
Respuesta de una población de N=88 neuronas registradas individualmente en el área IT durante la fase
'Cue'. Las neuronas muestran un mayor nivel de actividad para estímulos preferidos. (c) Cuando se
presentan los estímulos a elegir, las mismas células aumentan inicialmente su respuesta
independientemente de la elección. Al cabo de unos 170ms, las respuestas divergen dependiendo de si la
elección recae sobre el estímulo preferido o el antipreferido de la neurona. Esto ocurre antes del
movimiento ocular hacia el estímulo elegido, que se marca con la pequeña linea vertical sobre el eje x.
(d) Cuando ninguno de los estímulos coincide con el previamente mostrado, la respuesta a los dos
estímulos juntos es menor que la respuesta ante el estímulo preferido solo.
La figura 1.7 (d) muestra la comparación entre las respuestas dentro de la fase
'Target' cuando no hay atención, es decir cuando ésta recae en otro estímulo
completamente diferente. Se observa una supresión en el caso competitivo respecto al
caso en que sólo aparece el estímulo preferido.
12
Capítulo 1: Introducción
Cuando se comparan las respuestas ante estímulos únicos frente a estímulos
competitivos con atención en uno de ellos, lo que se obtiene es que el efecto de escoger
el estímulo preferido elimina la supresión en la respuesta que provoca el estímulo
antipreferido. Y cuando se escoge el estímulo antipreferido lo que se elimina es el efecto
excitador que provoca el estímulo preferido en la respuesta neuronal al estímulo
antipreferido, o nulo. Los efectos que se observan en tareas de competición sesgada
como las que hemos descrito son siempre cuantitativamente más importantes que las que
se observan con los paradigmas de estímulo único descritos previamente. Esto ha llevado
a proponer que los efectos producidos por la atención serían mejor entendidos en un
contexto competitivo entre estímulos y que su acción ante un estímulo individual
resultaría menos relevante (Desimone, 1998). Esto se contrapone al modelo de Atención
basada en Característica Global que enfatiza los efectos de la atención sobre la respuesta
neuronal a un estímulo único.
El aspecto crítico que distingue los experimentos que se han llevado a cabo al analizar
la atención local, en comparación con aquellos que han servido para definir la atención
global, es el hecho que en los primeros es fundamental presentar dos estímulos dentro
del campo receptivo, mientras que en los segundos sólo hay un estímulo en el campo
receptivo de la neurona. Explotaremos esta diferencia en nuestro modelo para conciliar
ambos cuerpos de resultados: al presentar un solo estímulo buscaremos reproducir los
resultados de la atención global basada en característica, y luego veremos cómo responde
el sistema a dos estímulos para interpretarlo según el modelo de la competición sesgada
y así evaluaremos el grado de compatibilidad o divergencia de estas dos visiones
contrapuestas.
1.4 Modelo computacional
Como se ha comentado en secciones anteriores, la atención visual ha sido estudiada
extensamente mediante experimentos de registro extracelular de neurona única en
monos y por medio de estudios con técnicas de imagen en humanos, demostrando que la
atención afecta la actividad en áreas sensoriales de la corteza cerebral que procesan
determinadas dimensiones de los estímulos sensoriales, como el color, el movimiento, la
textura y la forma. Asimismo, se ha determinado que la fuente de estos efectos proviene
de circuitos distribuidos en áreas superiores de la corteza cerebral. En concreto la corteza
frontal y parietal están involucradas en la generación y el control de señales de atención
recurrentes 'Top-down'. Existe un sustrato anatómico para estas influencias 'Top-down',
pues se ha demostrado que existen proyecciones recurrentes directas a las áreas visuales
V4 y TEO desde la corteza parietal (LIP, Lateral IntraParietal area); y a la parte anterior
de la corteza temporal inferior, TE, desde la corteza prefrontal; así como proyecciones
recurrentes indirectas desde la corteza prefrontal hasta las áreas V4 y TEO vía la corteza
parietal (Cavada y Goldman-Rakic, 1989; Ungerleider et al., 1989; Webster et al., 1994).
No se han estudiado este tipo de conexiones con el área MT, pero todo hace suponer que
éstas puedan existir.
Así pues, nuestra aproximación al problema de la Atención basada en Característica se
ha desarrollado mediante un modelo computacional que ha tenido en cuenta todos los
aspectos descritos anteriormente en este capítulo, y que está compuesto de dos redes
neuronales biofísicamente plausibles. Una de ellas representa un circuito de un área
superior, frontal o parietal, la cual por simplificación llamaremos únicamente red PFC
(PreFrontal Cortex), en vez de PFC/PPC (Posterior Parietal Cortex). La otra área simula la
parte sensorial, que en nuestro caso será el área visual selectiva a dirección de
movimiento, área MT.
13
Mecanismos de modulación por atención selectiva entre áreas corticales: Un modelo biofísico de red
Como se muestra en la figura 1.8, el modelo consiste básicamente en las tres
siguientes ideas clave:
● Entrada sensorial a MT de las neuronas de V1 selectivas al movimiento
● Conexión anterógrada de MT a PFC 'Bottom-up' que aporta la señal a almacenar en
WM
● Conexión retrógrada de PFC a MT 'Top-down' que genera la modulación por atención
en las neuronas de MT
La elección del área MT como área sensorial se justifica por varias razones:
primeramente porque es muy conocido su comportamiento tanto a nivel únicamente
sensorial (es el área visual más estudiada después de la corteza visual primaria) como su
modulación por atención (Born y Bradley, 2005). En segundo lugar, porque una de las
principales teorías que intentan explicar esta modulación proviene directamente del
estudio de esta área específica (Atención basada en Característica Global). Además, este
proyecto se desarrolla en un marco de colaboración con un laboratorio experimental que
estudia las modulaciones por atención en esta área sensorial (ver más abajo).
Finalmente, numerosos estudios de registro electrofisiológico y microestimulación en MT
han establecido una relación estrecha entre la actividad neural en esta área y la
percepción del movimiento (Britten et al. 1992; Salzman et al. 1990), por lo que es
plausible que también juegue un papel central en tareas de atención selectiva.
Figura 1.8: Esquema del modelo implementado para el estudio de la modulación Atención basada en
Característica en el área sensorial MT. Las ideas clave del modelo son las conexiones 'Bottom-up' y 'Topdown' entre las redes PFC y MT, las cuales generan la señal al sistema de WM y la modulación de la
respuesta sensorial, respectivamente.
Este trabajo, que en los próximos capítulos se va a detallar, ha sido fruto de la
colaboración internacional entre tres grupos de investigación y del apoyo económico de la
Fundación Volkswagen. Nuestros colaboradores en este marco son: un grupo
experimental (Centro de Primates de Alemania, en Göttingen) experto en el área MT y su
modulación por atención, liderado por el profesor Stefan Treue; y otro grupo
computacional experto en modelización de redes neuronales, encabezado por el profesor
Xiao-Jing Wang (Universidad de Brandeis, Massachusetts, Estados Unidos).
14
Capítulo 2: Métodos
Como ya se ha indicado en el capítulo introductorio, el principal objetivo de esta
investigación es estudiar qué mecanismos permiten a la atención modular la respuesta
sensorial conforme a las observaciones experimentales y para que podría llegar a ser útil
esta modulación desde el punto de vista del procesamiento de la información. Para poder
abarcar el estudio necesitamos restringir el problema a una región sensorial específica del
cerebro e implementar también, de forma biológicamente específica, la señal de atención
que producirá la modulación de la respuesta. En este trabajo se ha escogido el área
cortical temporal medial MT por ser una de las áreas sensoriales que más se ha estudiado
experimentalmente, tanto a nivel de la respuesta sensorial estricta, como a nivel de su
modulación por la atención. Las neuronas del área MT se caracterizan por responder
robustamente a la magnitud y dirección de la velocidad de los estímulos en su campo
receptivo. En este trabajo vamos a centrarnos en la selectividad a la dirección del
movimiento y en su modulación por atención. En cuanto a la señal de atención,
recogemos una de las hipótesis más plausibles actualmente, que ésta proviene de un
circuito en la región de la corteza prefrontal PFC, que tal y como se ha comentado en el
capítulo anterior, se trata de una región también ampliamente estudiada como fuente de
actividad persistente, supuestamente el correlato neural de la memoria de trabajo WM
(Goldman-Rakic, 1987; Fuster, 1988). Es por tanto requisito previo obtener modelos
plausibles para cada una de estas zonas independientemente, si se quiere estudiar con
rigor los mecanismos por los que la atención modula la respuesta sensorial.
En primera instancia, se desea que la red MT sea capaz de reproducir determinadas
características que se han observado experimentalmente, como: la actividad espontánea
en ausencia de estímulo visual (Maunsell y Van Essen, 1983; Mikami et al., 1986); la
curva de sintonía de estas neuronas ante estímulos de distinta dirección de movimiento, y
vamos a centrarnos en un determinado tipo de estímulo que consiste en pequeños puntos
luminosos distribuidos al azar en la escena (RDP: Random dot pattern) que se mueven en
una determinada dirección (Albright, 1984; Britten et al., 1993; Britten y Newsome,
1998; Treue et al., 2000); así como la respuesta cuando el estímulo consta de dos RDPs
superpuestos sobre el campo receptivo con movimiento transparente, esto es, cuando se
pueden percibir independientemente los dos movimientos (Snowden et al., 1991;
Snowden et al., 1992; Qian y Andersen, 1994; Treue et al., 2000).
Mecanismos de modulación por atención selectiva entre áreas corticales: Un modelo biofísico de red
Respecto a la red PFC, ésta ha de implementar la capacidad de generar actividad
persistente como correlato de WM, es decir, una red capaz de ser biestable, donde un
estado es el espontáneo, en el cual todas las neuronas disparan con tasas de disparo
bastante bajas, y el otro es el estado activo, en el cual una región localizada de neuronas
en la red dispara de forma mucho más notoria. La activación dependerá de un estímulo
externo localizado, tras el cual, si este es suficientemente intenso y duradero, la red por
sí sola será capaz de mantener la información relevante aunque el estímulo haya
desaparecido. Este modelo ya ha sido publicado previamente (Compte et al., 2000).
La modulación de la respuesta en base a la atención se implementará mediante la
unión de las dos redes individuales por medio de corrientes sinápticas, de forma que se
obtenga un bucle recurrente entre las redes MT y PFC. Tal y como se observa en la figura
2.1, el modelo se va a restringir a un solo campo receptivo, o lo que es lo mismo, un solo
módulo que representa una hipercolumna del área MT. Todas las neuronas de la red MT
comparten el mismo campo receptivo y su selectividad cubre todas las posibles
direcciones de movimiento. En cambio se supone que el circuito PFC recibe y emite
aferencias a todas las hipercolumnas de MT selectivas a la dirección del movimiento pero
de distinto campo receptivo.
PFC
MT
MT
MT
MT
RF
RF
RF
RF
...
MT
RF
Figura 2.1: Aproximación al problema. Se considera un único módulo MT, que representa una única
hipercolumna selectiva a la dirección del movimiento (sus neuronas comparten el campo receptivo) de
todo el conjunto que hay en el área MT (línea discontinua). En cambio, el circuito PFC se supone
específico de la dimensión del estímulo en cuestión, o información relevante (en este caso dirección de
movimiento), es decir puede que haya diferentes circuitos PFC pero éste será el único con aferencias
entrantes y salientes a todos y cada uno de los diferentes módulos MT selectivos a la dirección del
movimiento. Puesto que en el modelo sólo se va a considerar un módulo MT, las señales de los demás
módulos MT a PFC en el caso que se requieran serán consideradas como entradas externas.
2.1 El modelo de red
El modelo implementa la conexión recurrente, en forma de bucle, entre dos diferentes
áreas de la corteza cerebral, la primera representa una hipercolumna selectiva a dirección
de movimiento en el área MT, y la segunda modeliza un circuito local de la región
dorsolateral de PFC también selectivo a la dirección del movimiento (figura 2.2).
16
Capítulo 2: Métodos
Figura 2.2: Esquema del modelo desarrollado. El modelo forma un bucle recurrente entre dos redes que
representan dos regiones de la corteza cerebral, una de las cuales es una hipercolumna del área MT y la
otra un circuito local de la corteza prefrontal. Se puede observar que hay dos tipos de células
(representadas por los círculos), las piramidales (en rojo) y las interneuronas (en verde), y que éstas
están ordenadas en la red según la dirección preferida del estímulo (las flechas negras interiores).
También aparecen representadas todas las corrientes sinápticas entrantes y salientes de cada red
(flechas, con el mismo código de colores, rojo cuando las corrientes son excitadoras y verdes si son
inhibidoras; el grosor de la flecha indica la amplitud de la corriente) y la estructura que éstas poseen, en
forma de campana o plana según el caso (lineas azules).
Cada una de ellas contiene NE = 1024 células piramidales, o neuronas excitadoras, y NI
= 256 interneuronas, o neuronas inhibidoras. El número de neuronas excitadoras en el
modelo es cuatro veces superior al número de inhibidoras, manteniendo así la relación
observada experimentalmente en la corteza cerebral (Braitenberg y Schütz 1991; Abeles
1991). Las neuronas están ordenadas uniformemente en cada red según la dirección del
estímulo a la que son más sensibles, su dirección preferida, cubriendo uniformemente el
espectro de direcciones posibles, esto es, desde 0º hasta 360º. Por lo tanto, cada
neurona recibe como etiqueta el ángulo de su dirección preferida. Así pues, las neuronas
quedan distribuidas sobre una estructura de anillo, donde cada posición en el anillo tiene
una relación lineal con cada una de sus respectivas direcciones preferidas. Si uno va más
allá y relaciona la posición de las neuronas en la red con su localización espacial sobre la
corteza cerebral, esta arquitectura sería consistente con la organización columnar tanto
vista en áreas sensoriales (Lorente de Nó 1938; Mountcastle 1957; Mountcastle 1997)
como propuesta en las regiones dorsolaterales de la PFC (Goldman-Rakic, 1995; Rao et
al., 1999; Constantinidis et al., 1999; Ó Scalaidhe y Goldman-Rakic, 1999).
17
Mecanismos de modulación por atención selectiva entre áreas corticales: Un modelo biofísico de red
Se ha utilizado un modelo plausible biológicamente en el que las neuronas disparan
potenciales de acción (PAs), en comparación con otros modelos más simples en los cuales
no se generan los PAs sino que se calcula directamente la tasa de disparo. En concreto el
modelo que se ha empleado es el de integración y disparo con fugas (LIF: Leaky
Integrate-and-Fire) (Tuckwell, 1988), en el cual se integran las entradas sinápticas para
obtener el potencial de membrana Vm de la neurona por debajo de un determinado
umbral Vth. Una vez superado este umbral fijo, se considera que se ha producido un PA, e
inmediatamente después se actualiza el valor del potencial a otro valor subumbral,
comúnmente llamado reset Vr. Cada célula está caracterizada por seis parámetros
intrínsecos: los ya mencionados VTh y Vr, la capacitancia Cm, la conductancia de fugas gL,
el potencial de inversión de fugas VL y el tiempo refractario ref. Los valores de estos
parámetros en el modelo son los siguientes: VTh = - 50 mV, Vr = - 60 mV, Cm = 0.5 nF, gL
= 25 nS, VL = -70 mV y ref = 2 ms para células piramidales, y VTh = - 50 mV, Vr = - 60
mV, Cm = 0.2 nF, gL = 20 nS, VL = -70 mV y ref = 1 ms para interneuronas (Troyer y
Miller, 1997; Wang, 1999).
En cuanto a las corrientes sinápticas que reciben las neuronas, existen dos grandes
grupos: las externas a cada red y las recurrentes, o internas de cada red. Dentro de las
externas tenemos las que vienen de la otra red, las no-específicas que vienen de otras
zonas corticales (y contribuyen sólo a la actividad espontánea) y, en el caso de la red MT,
la entrada sensorial proveniente de zonas visuales previas. Todas ellas son entradas
excitadoras puesto que llegan de otras áreas corticales y la evidencia anatómica indica
que las neuronas de proyección cortico-corticales son de tipo piramidal, luego excitatorias
(Abeles, 1991). Además, en mi modelo he supuesto que estas corrientes están mediadas
únicamente por el receptor AMPA de glutamato, salvo en dos casos determinados en los
que por simplificar el modelo se ha supuesto una corriente no sináptica, como si se
hubiese inyectado directamente la corriente con un electrodo intracelular. Este tipo de
corriente inyectada se utiliza tanto para la entrada sensorial a MT, como para la señal de
compuerta o de alerta a PFC, entrada que indica un aumento en la actividad global de las
aferencias externas sin estructura a PFC debida al estado de alerta del sujeto sometido a
tareas de atención. Ésta es la forma de diferenciar los casos de atención y no atención en
la red PFC para generar, o no, la WM ante entradas sensoriales iguales, de ahí el nombre
de señal de compuerta. Los dos tipos de entrada que acabamos de mencionar se
estudiarán con más detalle en la próxima sección. En cuanto a las entradas internas, las
hay de dos tipos, excitadoras e inhibidoras. Las excitadoras van mediadas por los
receptores AMPA y NMDA, y las inhibidoras por el receptor GABAA.
En primer lugar, vamos a considerar las corrientes recurrentes de cada red, estas
corrientes postsinápticas son modeladas del siguiente modo (Wang, 1999):
Isyn t=gsyn stV m−V syn
[2.1]
donde gsyn s(t) es la conductancia sináptica en el instante de tiempo t, gsyn es la
conductancia máxima y s(t) es el grado de apertura del conjunto de canales en el
instante de tiempo t, y su rango está entre 0 y 1; Vsyn es el potencial de inversión
sináptico (Vsyn = 0 mV para sinapsis excitadoras y Vsyn = -70 mV para las inhibidoras).
Cuando un PA presináptico se genera, s(t) se incrementa instantáneamente para las
sinapsis mediadas por receptores AMPA y GABAA, y posteriormente decrece
exponencialmente con decay = 2 ms para AMPA (Hestrin et al., 1990; Spruston et al.,
1995) y decay = 10 ms para GABAA (Salin y Prince, 1996; Xiang et al., 1998).
18
Capítulo 2: Métodos
La conductancia NMDA es dependiente del voltaje (Jahr y Stevens, 1990):
gsyn=g syn
1
[Mg ]exp−0.062 V m
[2.2]
2
1
3.57
donde [Mg2+] = 1.0 mM. La dinámica del canal se modeliza según las siguientes
ecuaciones:
ds −1
=
ss x 1−s
dt s
[2.3]
dx −1
=
x∑ t−ti
dt x
i
[2.4]
donde s es la fracción de canales abiertos, x es una variable intermedia, ti son los
tiempos en que ocurren los PAs presinápticos, s = 100 ms es el tiempo de decaimiento, x
= 2 ms controla el tiempo de apertura de los canales NMDA y s = 0.5 kHz controla las
propiedades de saturación de los canales NMDA para altas frecuencias de disparo
presináptico.
La fuerza con la que se conectan las neuronas entre sí en general puede depender de
la diferencia entre sus direcciones preferidas. Esta propiedad se implementa considerando
que la conductancia sináptica entre la neurona i y la neurona j es:
gsyn, i, j=Wi− j Gsyn
[2.5]
donde W(i - j) representa la conectividad entre las dos neuronas, esto es como de
fuerte es la conexión. Esta conectividad está normalizada:
1
360
360
∫ Wi− j d j=1
[2.6]
0
La estructura que puede adoptar la conectividad puede ser plana (W constante) cuando
no existe estructura en las conexiones, esto es, cuando no hay dependencia con la
diferencia entre direcciones preferidas; o la suma de una gaussiana centrada en i - j =0,
más una constante:
−
−
Wi− j =J J −J exp
[
−i− j 2
2 2
]
[2.7]
J- representa la conectividad entre las células más dispares, J+ la conectividad entre las
más similares y es la anchura de la gaussiana, o lo que es lo mismo, si el incremento de
la conectividad respecto a la constante está muy localizado para neuronas muy similares
o, por el contrario, la estructura es cuasi plana. Dados dos parámetros el tercero se
puede calcular imponiendo la condición de normalización [2.6]. De todas las corrientes
sinápticas recurrentes de los modelos de área aislada, MT y PFC, la única con estructura
en la conectividad es la corriente sináptica entre neuronas excitadoras de la red PFC, con
19
Mecanismos de modulación por atención selectiva entre áreas corticales: Un modelo biofísico de red
J+ = 1.62 y = 36º. Cabe recalcar que en la red PFC juega un papel muy importante el
hecho de que la corriente recurrente excitadora a células excitadoras tenga este tipo de
estructura local () y una conductancia suficientemente fuerte (J+), pero no demasiado,
para que la actividad local reverbere y así se pueda generar biestabilidad, es decir, que
sea capaz de generar los dos estados espontáneo y activo en función de si le llega o no
entrada sensorial. Si la conectividad es demasiado débil o demasiado ancha, por fuerte
que sea la entrada sensorial, no se generará nunca actividad persistente, sólo será
estable el estado espontáneo, y si, en cambio, ésta es demasiado fuerte o demasiado
local, la generación del estado activo se generará espontáneamente sin necesidad de
ninguna entrada sensorial, o sea, el estado estable ahora será únicamente el activo. Para
ver más detalles de como se genera la biestabilidad se puede consultar la referencia
(Compte et al., 2000). Los valores concretos de conductancia recurrente para la red PFC
son: GAEE = 0.391 nS, GNEE = 0.732 nS, GAEI = 0.293 nS,, GNEI = 0.566 nS, GIE = 3.740 nS,
GII = 2.871 nS (el primer subíndice indica el tipo de conductancia, excitadora o inhibidora,
y en el caso de conductancias excitadoras aparece un superíndice indicando si se trata de
receptor AMPA o NMDA; el segundo subíndice hace referencia a que tipo de neurona
postsináptica se aplica). Las conductancias AMPA son lo suficientemente grandes como
para generar oscilaciones y sincronía entre las neuronas que disparan más rápidamente
en el estado activo (Compte et al., 2000).
En cuanto a la red MT, la corriente recurrente que juega un rol más importante es la
inhibidora a neuronas excitadoras, pues debe ser suficientemente grande como para que
la red MT por medio de ella reproduzca la normalización de la actividad de la población
observada experimentalmente. Además, esta corriente juega un papel importante en
como se modula la respuesta en MT por medio de la señal de atención proveniente de
PFC. Estos dos efectos, la normalización y la modulación en MT, se estudiarán con más
detalle en el siguiente capítulo de resultados. Los valores concretos de conductancia
recurrente para la red MT son: GAEE = 0.005 nS, GNEE = 0.093 nS, GAEI = 0.005 nS,, GNEI =
0.195 nS, GIE = 1.465 nS, GII = 0.391 nS.
Vamos ahora a considerar las corrientes de entrada externas, y empezaremos por las
corrientes recurrentes entre las dos redes. Estas conexiones son excitadoras y mediadas
por el receptor AMPA, con lo cual, se mantiene lo que se ha dicho más arriba sobre la
dinámica de este tipo de sinapsis (ecuaciones [2.1] y [2.5]). Lo que cambia es la
estructura de la conexión, pues ahora ésta es Gaussiana:
Wi− j =exp
[
−i− j 2
2 2
]
[2.8]
manteniéndose la normalización descrita en [2.6]. Estas entradas son muy
importantes, pues son las responsables, en el caso en que la atención está presente, de
la generación (junto con la señal compuerta) de la WM en PFC y de la modulación de la
respuesta en MT, pero deben, a su vez, ser suficientemente pequeñas para que su
influencia no sea notoria cuando no hay atención. Para la generación de la actividad
persistente, es necesaria y suficiente la conexión excitadora de MT únicamente a
neuronas excitadoras de PFC. No obstante, para obtener la modulación adecuada de la
respuesta sensorial en base a la atención, necesitamos que la proyección excitadora de
PFC incida tanto en neuronas excitadoras como inhibidoras de MT. Dada la dominación de
la inhibición recurrente en MT, se necesita que la conexión excitadora sea mucho más
fuerte. Los valores usados para las conductancias son: GMT-PFC = 0.005 nS, GPFC-MTE = 0.098
nS, GPFC-MTI = 0.024 nS (el primer superíndice hace referencia a la neurona presináptica y
el segundo a la neurona postsináptica, el subíndice se aplica para diferenciar entre las
neuronas postsinápticas excitadoras e inhibidoras, en los casos en que éste no aparece se
20
Capítulo 2: Métodos
entiende que las neuronas diana son excitadoras); y las anchuras de las respectivas
conectividades son: MT-PFC = 36º, PFC-MTE = 72º, PFC-MTI = 72º.
Como ya se ha mencionado anteriormente, todas las neuronas reciben además
corrientes externas de entrada desde otras áreas corticales. Estas entradas son
necesarias para obtener unos valores adecuados en las respuestas espontáneas de las
dos redes MT y PFC en torno a 9 Hz y 2 Hz, respectivamente. En el modelo estas entradas
se modelizan mediante un proceso Poisson, a través de trenes de PAs no correlacionados.
La tasa de disparo del tren de espigas es ext = 1.8 kHz, (equivalente a 1000 entradas
diferentes no correlacionadas cuya tasa de disparo sea 1.8 Hz). Esta entrada externa es
mediada únicamente por receptores AMPA, con una máxima conductancia a neuronas
excitadoras que difiere según la red considerada: gext,E = 3.1 nS en la red PFC y gext,E = 10
nS en la red MT, en cambio el valor para las neuronas inhibidoras se mantiene igual para
las dos redes, gext,I = 2.38 nS. La diferencia entre las conductancias excitadoras de las dos
redes deriva de dos factores: la tasa de disparo espontánea es mayor en MT, y el dominio
de la inhibición recurrente en MT, hecho que se ha de compensar con el incremento de
corriente externa.
2.2 Estímulos visuales y tareas de atención
En este apartado se va a describir el tipo de estímulo y las tareas que se han utilizado
experimentalmente, con las cuales se ha observado tanto la normalización de la actividad
de la población (Treue et al., 2000), como la modulación de la respuesta neuronal debida
a la atención (Martinez-Trujillo y Treue, 2004).
Este estímulo, ya mencionado anteriormente y que designamos con las siglas RDP,
consiste en un conjunto de puntos luminosos, con forma de pequeños cuadrados,
situados al azar dentro del campo receptivo de la neurona. Estos puntos, en el caso que
nos contempla, se mueven coherentemente en una determinada dirección. En el caso del
movimiento transparente se tienen dos RDPs independientes con igual número de puntos
cada uno pero con distinta distribución en el espacio, y moviéndose por separado en
direcciones diferentes. Observar la figura 2.3.
Single RDP
Transparent RDPs
Figura 2.3: Estímulo sensorial. El estímulo utilizado en los experimentos es el llamado RDPs, en el cual
los puntos están situados al azar dentro del campo receptivo y moviéndose en una determinada
dirección. Para el caso transparente hay dos RDPs diferentes en direcciones igualmente diferentes.
En cuanto a las tareas utilizadas, hay dos tipos diferentes según se utilicen RDPs
simples o transparentes (figura 2.4). Con RDPs simples se puede estudiar como es la
curva de sintonía de las neuronas de MT y su modulación con la atención. En este caso la
atención asociada al movimiento está siempre fuera del campo receptivo y en el modelo
la aparición del estímulo a atender será tratada como una entrada externa a PFC, de otro
21
Mecanismos de modulación por atención selectiva entre áreas corticales: Un modelo biofísico de red
supuesto módulo MT no incluido, pero que reproducirá la misma respuesta en el circuito
PFC que la que generaría la propia red MT. Una corriente externa de amplitud 0.025 nA y
= 36º simula bien la entrada de MT en este caso. Con RDPs transparentes podemos
estudiar la normalización de la actividad de la población y además se puede predecir, ya
que aún no se han hecho experimentos en este sentido, como influye la atención en este
caso. En este caso la atención asociada al movimiento está siempre dentro del campo
receptivo.
RDPs simples
Att on fixation
t
Att-Sens = constant
Att fixed
+
+
+
+
+
+
+
+
+
RDPs transparentes
t
Att on fixation
Att-Sens = constant
Att fixed
+
+
+
+
+
+
+
+
+
Figura 2.4:Esquema de las tareas de atención. Se consideran dos tipos de tareas, los que consideran sólo
estímulos simples y los que consideran estímulos transparentes. Cada flecha representa un RDP
moviéndose en una determinada dirección dentro de un conjunto de ensayos. El color rojo representa el
estímulo atendido y el negro al sensorial. Hay tres etapas diferentes en cada ensayo señalizadas a través
de la flecha del tiempo (flecha vertical en azul): aparición del estímulo a atender (RDP o punto de
fijación); pausa, sin ningún tipo de movimiento en la escena; y finalmente aparición del estímulo
sensorial juntamente con el atendido. Consideramos caso de atención únicamente cuando ésta recae
sobre RDPs y no sobre el punto de fijación, puesto que este tipo de atención no afectaría la respuesta en
nuestro módulo de MT. Se necesitan una serie de ensayos para generar las curvas de sintonía (serie de
flechas que cubren todas las direcciones posibles del estímulo sensorial). En cuanto a la atención, ésta se
puede mantener fija entre los diferentes ensayos, o, en cambio, lo que se puede mantener fijo es la
relación entre las direcciones de movimiento de los RDPs atendido y sensorial. Según el caso, la
modulación observada será diferente.
22
Capítulo 2: Métodos
En cada tarea se tienen tres etapas diferentes durante las cuales se fija la vista sobre
un punto inmóvil en el centro de la pantalla: en la primera de las etapas aparece el
estímulo al que se tiene que atender (que aparece en rojo en la figura 2.4), este puede
ser un RDP moviéndose en una determinada dirección, o el mismo punto fijo al cual se
mira directamente; en la segunda hay como una pausa, durante la cual, sea cual sea el
estímulo atendido, no aparece ningún movimiento en la pantalla; en la tercera y última
etapa, vuelve a aparecer el mismo estímulo atendido junto con otro RDP al que
llamaremos RDP sensorial, puesto que este está siempre dentro del campo receptivo
(aparece en negro, o gris, en la figura 2.4). La tarea termina una vez cambia de alguna
manera el estímulo atendido, si se trata del RDP, el cambio puede darse en dirección de
movimiento o en la velocidad. Si se trata del punto fijo, este puede cambiar de color.
Cabe recalcar que en lo que respecta a la respuesta de las neuronas en MT, la modulación
por atención únicamente se dará cuando la dimensión atendida del estímulo esté
relacionada con el movimiento, así consideraremos el caso de atención al punto fijo como
caso 'sin atención'.
Para obtener la curva de sintonía de una determinada neurona de MT, se necesitan un
conjunto de ensayos, en los cuales el RDP sensorial recorre todo el conjunto de
direcciones posibles desde 0º hasta 360º (diferentes flechas negras en la figura 2.4). En
cambio la respuesta de la población a un determinado estímulo requiere un solo ensayo.
Para el caso de atención, hay dos diferentes posibilidades, que la atención esté fija
entre ensayos (una sola flecha roja en la figura 2.4) o que ésta varíe manteniendo fija la
diferencia respecto al RDP sensorial (múltiples flechas rojas en la figura 2.4). Según sea
una u otra la situación, la modulación de la respuesta neuronal cambiará, tal y como se
verá en el capítulo siguiente de resultados. Conviene recordar que consideramos curva de
sintonía a la representación gráfica de la respuesta de una única neurona respecto al
cambio de valor de una deteminada dimensión del estímulo, manteniendo todo lo demás
en condiciones constantes. En presencia de atención, tradicionalmente se ha considerado
como curvas de sintonía, o curvas de sintonía neuronales, a las curvas de respuesta en
las se ha mantenido constante, o fija, la característica atendida. En cambio, en los casos
en que la atención es intrínseca al estímulo, como cuando atención y estímulo van
ligados, se ha considerado que esta curva de respuesta representa la actividad de la
población, o curva de sintonía de la población, porque resulta equivalente a la respuesta
que tendrían las neuronas en su conjunto frente a un único de estos estímulos atendidos
(Martinez-Trujillo y Treue, 2004). Esta equivalencia ha sido testeada en el modelo.
Como se ha mencionado más arriba el modelo necesita alguna manera para diferenciar
entre atención y no atención, es por tanto necesaria una señal de alerta o compuerta. Tal
y como se describe en la figura 2.5, esta señal sólo aparece en la primera etapa de la
tarea de atención, no tiene estructura y por sí sola es incapaz de generar el estado activo
en PFC, no obstante es indispensable que coincida con la señal sensorial proveniente de
MT para que se genere actividad persistente en PFC. Esta señal puede ser interpretada
como un incremento en la fuerza de las aferencias externas de otras zonas del cerebro a
la red PFC debido al estado de alerta del sujeto. En el modelo un valor de corriente de
0.01125 nA es adecuado.
Vamos ahora a considerar como se ha modelizado la entrada sensorial a MT para los
estímulos visuales RDPs. Esta corriente representa la entrada proveniente de áreas
visuales previas, en concreto del área V1, en la que existen una tercera parte de células
que son selectivas a dirección de movimiento (Hawken et al., 1988), y que forman la
principal aferencia a MT (Zeki, 1974; Maunsell y Van Essen, 1983; Albright, 1984; Shipp
y Zeki, 1985; Fries et al., 1985; Mikami et al., 1986; Born y Bradley, 2005). La forma
que se ha supuesto para esta entrada está íntimamente relacionada con la actividad de
23
Mecanismos de modulación por atención selectiva entre áreas corticales: Un modelo biofísico de red
este tipo de neuronas de V1 ante estímulos visuales RDPs, tanto simples como
transparentes. Se han observado experimentalmente tres importantes características: la
saturación de la respuesta máxima que puede alcanzar una neurona de V1 con el
aumento del número de puntos del RDP moviéndose en la dirección preferida (Snowden
et al, 1992) (figura 2.6); la supresión en la actividad neuronal cuando se comparan las
respuestas entre los casos de RDPs simples preferido y nulo (este último es un RDP en
igual dirección pero sentido opuesto al preferido), la cual indica la selectividad a dirección
de movimiento; y la inexistencia de ningún tipo de supresión notoria cuando se comparan
las respuestas entre el caso de RDPs transparentes preferido+nulo con respecto al caso
RDP preferido simple (Snowden et al, 1991) (figura 2.7).
Attentional case
Gating
t
Gating alone
Gating
Non-attentional case
Gating
PFC
PFC
PFC
MT
MT
MT
Gating
Gating
Gating
PFC
PFC
PFC
MT
MT
MT
Gating
Gating
Gating
PFC
PFC
PFC
MT
MT
MT
Figura 2.5: Señal de alerta o compuerta. Se necesita este tipo de señal para diferenciar los casos de
atención y no atención. Así, ni la señal proveniente de MT, ni la señal de compuerta solas son capaces de
generar la actividad persistente, sólo la combinación síncrona de las dos es capaz de producir WM. La
señal de compuerta sólo aparece en la primera etapa del ensayo y no posee estructura localizada, al
contrario de la señal sensorial entrante desde MT.
Estas dos figuras 2.6 y 2.7 nos indican que para los casos extremos de estímulos
transparentes hay una normalización a la actividad máxima posible, la cual coincide con
la respuesta al estímulo RDP simple en la dirección preferida. Así el caso extremo de dos
RDPs desplazándose en la dirección preferida genera la misma actividad que uno solo
siempre que, como ocurre, la densidad de puntos sea suficiente. Y en el otro extremo,
cuando el otro RDP se desplaza en la dirección nula, éste tampoco afecta la respuesta
neuronal.
Una suposición plausible usada en el modelo es que este tipo de normalización a la
máxima respuesta posible se cumple, no sólo para las separaciones extremas entre
direcciones de movimiento de los RDPs superpuestos, sino para todas las intermedias
24
Capítulo 2: Métodos
(figura 2.8) y que la entrada a MT es lineal con este tipo de actividad en V1 para RDPs.
(He estado estudiando este tipo de normalización en V1, con modelos similares a los que
aquí se han descrito para MT y PFC, obteniendo resultados con muy buen grado de
aproximación a las respuestas representadas en la figura 2.8. En este estudio se ha
supuesto que la entrada a esta población de células de V1 es ya selectiva a dirección pero
que ésta no está normalizada para RDPs transparentes, sino que es lineal. Es la propia
red la que se encarga de conseguir este tipo de normalización, que podría ser descrita
como normalización al contraste. Este estudio fue mi proyecto de investigación en el
curso de verano: Advanced Course in Computational Neuroscience. Agosto 1-26, 2005,
Arcachon, Francia. Espero añadir en breve la red V1 a mi modelo).
normalized
responses
Number of dots
Figura 2.6: Normalización de la actividad de las neuronas de V1 selectivas a dirección de movimiento
para RDPs con número de puntos variable que se desplazan en la dirección preferida. Se puede observar
como a medida que se aumenta la densidad de puntos, la respuesta aumenta hasta llegar a saturarse
para valores suficientemente altos. (Snowden et al., 1992)
Se ha utilizado para representar la entrada sensorial de cada RDP a MT, una corriente
con forma de campana que marca la selectividad a la dirección proveniente de V1. Esta
entrada ha de ser periódica, como la dirección de movimiento, por lo tanto no se puede
usar la curva Gaussiana. La ecuación utilizada ha sido una que se aproxima mucho a ella:
[
I=I0I1 exp cos−c −1
]
[2.9]
Donde . Y para RDPs transparentes, la normalización al máximo se consigue
sumando las dos campanas y reescaleando el resultado para que no supere el valor I0+I1.
Los parámetros adecuados para esta corriente han sido: I0 = 0.5 nA, I1 = 0.6 nA, =
36º para las neuronas excitadoras y la quinta parte del valor de esta corriente para las
inhibidoras. Debido a la dominación de la inhibición recurrente en MT, se necesita que la
entrada excitadora sea mucho más fuerte.
25
Mecanismos de modulación por atención selectiva entre áreas corticales: Un modelo biofísico de red
Representative V1 cell
Figura 2.7: Respuesta de una neurona representativa de V1 selectiva a dirección de movimiento expuesta
a estímulos RDPs simples y transparentes. Se puede observar en primer término que la neurona es
selectiva a dirección comparando la actividad para RDPs entre la dirección nula (columna izquierda) y la
preferida (columna central). Otra importante observación es que no se aprecia supresión en la actividad
cuando comparamos el estímulo transparente preferido+nulo (columna derecha) respecto al caso
preferido simple. (Snowden et al., 1991)
25
Resp
(Hz)
0
Direction ( º )
Figura 2.8: Respuesta de una hipotética población de V1 ante RDPs transparentes. Las separaciones
entre los RDPs se codifican en los colores de las curvas de respuesta: marrón, RDP simple y dual sin
separación; cián, RDPs transparentes con separación de 60º; azul, RDPs transparentes con separación de
120º; y rojo, RDPs transparentes con separación de 180º. Se puede observar que el máximo de la
respuesta es constante independientemente de la separación entre los RDPs.
2.3 Materiales usados
El modelo que se ha desarrollado ha sido implementado en un ordenador con sistema
operativo Linux y procesador Pentium IV a 2.8 GHz con 512 kB de memoria cache y 1024
MB de memoria RAM. Las simulaciones que requerían repeticiones sucesivas de ensayos
se han completado en un cluster de computación Linux+OpenMosix formado por 28 CPUs
de 3.0 GHz. El código del programa ha sido escrito en lenguaje C++. El método de
26
Capítulo 2: Métodos
integración usado ha sido el algoritmo Runge-Kutta de segundo orden con interpolación
de los tiempos de disparo (Hansel et al., 1998) y el paso de integración ha sido 0.02 ms.
Correr una simulación de 5 s en este sistema tarda aproximadamente unos 30 minutos.
Me gustaría recalcar que al tiempo que se ha desarrollado el modelo descrito en este
capítulo, también se ha implementado una aproximación de campo medio, la cual
reproduce los resultados estacionarios de la simulación en un grado muy notable y con la
ventaja que es mucho más rápida (unos pocos segundos). La desventaja es que no
proporciona información acerca de aspectos dinámicos de la actividad, como oscilaciones
o inestabilidades. Se ha usado este modelo para explorar eficientemente el conjunto de
parámetros utilizado en la simulación. Describir a fondo este modelo en su fundamento
teórico queda fuera de esta memoria por resultar demasiado matemático (para
profundizar en este tipo de modelos de campo medio se puede consultar la referencia
Renart et al., 2003).
27
Capítulo 3: Resultados
La finalidad última del capítulo es proponer una base para la comprensión de, por un
lado, como actúa la atención visual en cuanto a la base biológica y los mecanismos
involucrados; y por otro, dónde y cómo influye ésta, en qué zonas del cerebro actúa y
cuáles son las modulaciones de actividad observadas. Para ello se partirá de la teoría
propuesta por Treue descrita previamente en el capítulo de introducción, el Modelo de
Ganancia dependiente de la Similitud respecto a Característica (Treue y Martinez-Trujillo,
1999; Martinez-Trujillo y Treue, 2004). En nuestro caso la característica visual atendida
es una determinada dirección de movimiento. Posteriormente, trataremos de relacionar
nuestro análisis con otra teoría vigente, también comentada en la introducción, el Modelo
de Competición Sesgada (Moran y Desimone, 1985; Desimone, 1998). Veremos que estas
dos teorías pueden ser vistas como dos caras de una misma moneda. De hecho, en los
casos en los que hay competición entre diferentes estímulos visuales, demostraremos que
la modulación de la atención descrita por Treue reproduce a la vez las tesis del modelo de
Desimone. Esta idea ha aparecido recientemente en la literatura (Boynton, 2005). La
diferencia entre ese estudio y el nuestro, radica en que el modelo de Boynton es
puramente ad hoc sin preocuparse de qué base fisiológica permite este tipo de efectos, en
cambio nuestro modelo es plausible desde el punto de vista biológico, lo que nos permite
estudiar cuáles son las bases y mecanismos del cerebro que implementan las teorías de
Treue/Desimone y su conciliación en la línea de Boynton.
Para ello se han realizado una serie de simulaciones con el modelo. Como se mencionó
en el capítulo anterior, existen dos grandes tipos de tareas de atención, según se
consideren RDPs simples o transparentes. En este capítulo vamos a ver cuales son los
resultados obtenidos con las simulaciones y los compararemos con las observaciones
experimentales que sustentan la teoría de Treue, e iremos más lejos, formulando
predicciones a nuevos experimentos nunca realizados previamente, los cuales, por un
lado, vendrán a confirmar las tesis de Treue, y por otro, nos permitirán relacionarlas
también con la teoría de Desimone.
Veremos qué tipo de información aportan las simulaciones mediante ejemplos de la
salida del modelo. Analizaremos los tres casos diferentes que se pueden encontrar en
cada tipo de tarea: no atención, atención fija y relación (señal atención-estímulo
sensorial) fija (ver métodos, figura 2.4), en cada una de ellas se verá que se obtiene un
tipo de modulación diferente, como ocurre en los casos observados experimentalmente.
Mecanismos de modulación por atención selectiva entre áreas corticales: Un modelo biofísico de red
3.1 Resultados con RDPs simples
Empezaremos mostrando dos ejemplos característicos de este tipo de simulaciones. En
ellas se muestran los esquemas de las tareas respectivas (al estilo de la figura 2.4) y cuál
es la salida de las dos redes en los dos casos (figura 3.1). En una situación la atención
influye en la respuesta modulando la respuesta sensorial, y en la otra no, puesto que se
atiende a otra dimensión no relacionada con el movimiento.
attention
non-attention
+
t
+
t
+
+
+
+
rate
55
PFC
rate
55
PFC
location
location
55
MT
55
MT
location
location
time (s)
0
time (s)
5
0
5
Figura 3.1: Resultados de dos simulaciones características para RDPs simples. En la parte superior se
muestran los diagramas de las tareas, al estilo de la figura 2.4 del capítulo anterior. En la parte inferior
se muestra la actividad de las poblaciones mediante un código de gradiente de color, las respuestas
neuronales con mayor tasa de disparo aparecen en rojo y aquellas con menor tasa de disparo en azul
oscuro. Las tres diferentes etapas de la tarea se separan con lineas verticales negras. La parte izquierda
de la figura considera un caso en que la atención se centra en una dirección del movimiento y afecta al
procesamiento sensorial modulando la actividad en el circuito MT, mientras que la parte derecha refleja la
situación de no atención, y el procesamiento en MT no se ve afectado por la actividad en PFC.
De la observación directa de estos resultados se puede extraer información:
● Aparece actividad sostenida en PFC sólo en el caso de atención.
● Ésta se generaría por las entradas externas simultáneas: la entrada sensorial
proveniente de otra red MT (puesto que la atención está fuera del campo receptivo
considerado), juntamente con la señal de alerta o compuerta.
● El caso de no atención no genera actividad sostenida debido a la falta de la entrada
sensorial en la primera fase, y a la falta de señal de compuerta en la tercera.
● Si comparamos la actividad en MT en los dos casos, se observa que la respuesta
neuronal máxima aumenta con la atención, y que esto ocurre a lo largo de las tres
etapas de la tarea, también en ausencia de estímulo visual, con actividad
espontánea. Algo similar, pero menos notorio a simple vista, es lo que ocurre con la
respuesta mínima, que disminuye en el caso atendido.
30
Capítulo 3: Resultados
3.1.1 Cruce de las curvas de sintonía de la población
Así como acabamos de ver, en el caso de atención sobre la dirección del movimiento
del estímulo, se produce una disminución de la respuesta mínima y un aumento de la
respuesta máxima, de forma que el efecto neto de esta combinación es un cruce en las
curvas de respuesta de la población MT entre los dos casos, atención/no atención,
aumentando así la selectividad al estímulo sensorial (Figura 3.2).
Representative MT cell
MT model network
att.
no att.
Neuron
Figura 3.2: Modulación por atención de la actividad en MT. En la parte izquierda se muestra la
modulación de la curva de respuesta de una célula característica del área MT cuando la atención recae
sobre el estímulo (Martinez-Trujillo y Treue, 2004). En la parte derecha se muestra la modulación de la
actividad de la red MT en el modelo. Las dos curvas, curva de respuesta y actividad de la población son
equivalentes, pues se puede demostrar que la actividad de la población en la red para un estímulo dado
reproduce la curva de respuesta neuronal siempre que se mantenga la relación entre señal de atención y
estímulo sensorial constante. Se observa como el modelo es capaz de reproducir los datos
experimentales: cruce de las curvas de sintonía de la población entre los casos de atención sobre el
estímulo y no atención, aumentando la selectividad al estímulo.
En la figura 3.2, se está comparando la curva de respuesta experimental por un lado
(Martinez-Trujillo y Treue, 2004), con la actividad de la población en el modelo por el
otro. En el caso de no-atención, es fácil ver que esto se puede hacer así porque se puede
demostrar que la actividad de la población para un estímulo dado coincide con la curva de
respuesta de una neurona. Recordemos que la curva de respuesta se obtiene a partir de
múltiples ensayos para todos los estímulos posibles a lo largo de todo el rango de
direcciones.
Cuando además de estímulo sensorial tenemos señal de atención (entrada proveniente
de PFC cuando hay actividad sostenida), esta correspondencia entre curva de respuesta
neuronales y actividad de la población en la red se mantiene sólo si la curva de respuesta
de la neurona se obtiene manteniendo siempre fija la relación entre el estímulo sensorial
y la señal de atención para los diferentes ensayos. Así, en la figura 3.2, la señal atención
y el estímulo sensorial siempre van parejos.
Se observa como el modelo es capaz de reproducir los datos experimentales: cruce de
las curvas de respuesta entre los casos de atención sobre el estímulo y no atención,
aumentando la selectividad al estímulo.
31
Mecanismos de modulación por atención selectiva entre áreas corticales: Un modelo biofísico de red
Dadas las curvas de respuesta, nos podemos preguntar cómo están moduladas, cuáles
son los diferentes factores de modulación punto a punto. A simple vista se observa un
cruce en las curvas, por tanto en unas situaciones encontraremos un efecto multiplicativo
(factor mayor que uno) y en otros efecto divisivo (factor menor que uno). La figura 3.3
muestra la forma de los diferentes factores de modulación en función de la distancia a la
que se encuentra la señal de atención.
MT population
MT model network
sep. from attention signal dir.
Figura 3.3: Factores de modulación en la actividad de la población. En la parte izquierda se muestran los
factores de modulación promediados sobre una población de neuronas de MT (N=135) (Martinez-Trujillo
y Treue, 2004). En la parte derecha se muestran estos factores para la red MT del modelo. La curva
resultante en el modelo mantiene una forma similar a lo que ocurre en la población de MT, aunque la
máxima modulación en el modelo y la separación para la cual no hay modulación (factor uno) son un
poco mayores, quedando entre el promedio de la población MT y la célula representativa de la figura 3.2
(Martinez-Trujillo y Treue, 2004).
La curva de los factores de modulación en el modelo reproduce lo que ocurre en la
población promediada de MT (N=135), aunque se puede observar que la máxima
modulación y la separación para la cual no hay modulación (factor uno) son un poco
mayores para el modelo, quedando estos parámetros entre los resultantes para el
promedio de la población MT y la neurona característica de la figura 3.2 (Martinez-Trujillo
y Treue, 2004).
A la vista de la figura 3.3, en la que vemos que los factores de modulación decrecen
con la distancia relativa a la señal de atención, se puede prever que las curvas de
respuesta con atención se cruzarán también, y cada vez más, a medida que alejemos el
estímulo sensorial de la señal de atención. Podemos comprobar que esto ocurre así en el
modelo (Figura 3.4). La máxima selectividad al estímulo se obtendrá cuando la atención
recae sobre él.
3.1.2 Efecto multiplicativo de las curvas de sintonía neuronales
Como se ha visto en el capítulo de la introducción, en diferentes zonas del cerebro la
modulación de la respuesta selectiva a la característica es, muchas veces, multiplicativa
(McAdams y Maunsell, 1999; Treue y Martinez-Trujillo, 1999). Pero acabamos de ver en
MT que la modulación produce un aumento de la selectividad, o sea el cruce de las curvas
de respuesta. La hipótesis de Treue para reconciliar ambas observaciones experimentales
32
Capítulo 3: Resultados
radica en los factores de modulación y su dependencia con la separación respecto a la
señal de atención (figura 3.3). Según el Modelo de Ganancia dependiente de la Similitud
respecto a Característica, con atención cada neurona modifica su respuesta de forma
multiplicativa, siendo el factor multiplicativo función sólo de la similitud entre la
característica atendida y la preferencia de la neurona, independientemente de la
naturaleza del estímulo que se presente. Así, cuando se atiende a movimiento hacia
arriba, las neuronas que son selectivas a esa dirección del movimiento multiplicarán su
respuesta por un factor mayor que uno, mientras que las neuronas selectivas al
movimiento hacia abajo reducirán su respuesta multiplicando por un factor menor que
uno.
Así si obtenemos las curvas de sintonía manteniendo fija la señal de atención sobre
una característica mientras variamos la posición del estímulo sensorial, obtendremos un
efecto neto multiplicativo o divisivo según si estamos mirando las respuestas de una
neurona con preferencia coincidente o divergente con la característica atendida,
respectivamente. Esto sería así puesto que los factores de modulación de cada neurona
se mantendrían constantes al variar la localización del estímulo (y no la de la atención) y
resultarían en curvas escaladas multiplicativa o divisivamente.
att.-stim
0
90
180
Neuron
+
+
+
+
Figura 3.4: Cruce de las curvas de respuesta. El gradiente de colores marca la separación entre el
estímulo sensorial respecto a la señal de atención. A medida que aumenta la separación, la respuesta
máxima disminuye y la mínima aumenta, de manera que el cruce entre las curvas aumenta. La máxima
selectividad al estímulo se obtiene cuando la atención recae sobre él. Los dibujos de abajo muestran
como tendrían que ser los distintos ensayos para reproducir los resultados para curvas de sintonía en vez
de para la actividad de red. Como se trata de reproducir la actividad de la población, los ensayos
mantendrían en todo momento una relación fija entre el estímulo que se presenta en el campo receptivo
(círculo discontinuo) y el estímulo que se atiende fuera del campo receptivo. Cuando esta relación es de
paralelismo (esquema más a la izquierda, color marrón en el gráfico de arriba) se obtiene la máxima
selectividad, mientras que cuando esta relación es de antiparalelismo (esquema más a la derecha, color
azul oscuro en el gráfico de arriba) se obtiene el perfil de respuesta más chato, es decir la mínima
selectividad.
Bajo este formalismo no se han obtenido aún las curvas de sintonía experimentalmente
para la atención basada en característica sobre la dirección del movimiento (todos los
experimentos previos que calculaban la curva de sintonía incluían aspectos importantes
de atención espacial, que aquí no tratamos), pero, tal y como se ha comentado en el
33
Mecanismos de modulación por atención selectiva entre áreas corticales: Un modelo biofísico de red
capítulo de introducción, sí se han obtenido unas relaciones multiplicativas en las
respuestas cuando se comparan determinados ensayos, entre los cuales lo único que
cambia es que la atención pasa de la dirección nula de la neurona a la preferida
(Martinez-Trujillo y Treue, 2004).
El mismo modelo capaz de reproducir el cruce de las curvas de sintonía de la
población, del apartado anterior, es capaz de generar curvas de sintonía neuronales
multiplicativas, sin ningún tipo de cambio en sus parámetros intrínsecos, únicamente por
medio de mantener la señal de atención fija en una determinada dirección de
movimiento, mientras se presentan a la red estímulos sobre todo el rango de direcciones
de movimiento posibles para calcular las curvas de sintonía de las neuronas de la red.
En la figura 3.5 se muestran las curvas de sintonía extraídas del modelo para
diferentes simulaciones en las cuales la atención se ha mantenido fija en: la dirección
preferida; la dirección nula; y la dirección preferida más 90º, más o menos la separación
para la cual el factor de modulación vale uno en el modelo (figura 3.3). Se puede
observar que la relación es multiplicativa entre unas y otras con un alto grado de
aproximación, puesto que con un escalado respecto a una de ellas se obtiene que las tres
coinciden de forma notable. Aparecen pequeñas desviaciones entre ellas que indican que
el escalado no es perfectamente multiplicativo, pero el grado de aproximación es al
menos tan bueno como en el experimento y, de forma muy significativa, concilia las
observaciones de escalado multiplicativo y cruce de curvas de respuesta que podían
parecer contradictorias a simple vista.
MT model neuron response
scaled responses
pref.
no pref.
at. 90
Figura 3.5: Efecto multiplicativo de las curvas de sintonía. El mismo modelo capaz de obtener el cruce en
las curvas de sintonía cuando la relación (estímulo sensorial- señal de atención) permanece constante, es
capaz de generar curvas de sintonía multiplicativas cuando la atención es fija. La atención se fija a la
dirección preferida (en rojo), a la nula (en azul) y a la preferida más 90º (en verde). Se puede ver que
son multiplicativas cuando se escalan las curvas de sintonía (parte derecha de la figura).
El efecto multiplicativo se puede generalizar cualquiera que sea la dirección en la cual
se mantiene fija la atención (figura 3.6).
34
Capítulo 3: Resultados
att
0
90
180
+
+
+
+
Figura 3.6: Efecto multiplicativo en las curvas de respuesta. El gradiente de colores marca en qué
dirección está fijada la señal de atención en relación a la dirección preferida de la neurona. A medida que
la atención se aleja de la dirección preferida, el factor multiplicativo disminuye. Los dibujos de abajo
muestran como son los distintos ensayos para reproducir los resultados para las curvas de sintonía. Es
importante hacer notar la diferencia sustancial y crítica entre estos ensayos y los que se indican en la
Figura 3.4: en los ensayos que indicamos aquí, la característica que se atiende al calcular una curva de
respuesta neuronal (el estímulo fuera del campo receptivo) se mantiene fija según se varía los estímulos
sensoriales en el campo receptivo. En cambio, en la Figura 3.4 ambos estímulos, dentro y fuera del
campo receptivo, covariaban al desplazarse y mantenían fija la relación entre señal de atención y
estímulo sensorial.
Si observamos la figura 3.3 vemos que el cruce por uno de la curva de los factores de
modulación se da para una separación entre estímulo y atención próxima a 90º. Así pues
esperamos que la curva de respuesta con atención fija a la dirección preferida más 90º y
la de no atención sean muy similares (figura 3.7). El ajuste sería casi perfecto si el cruce
se diese exactamente a 90º y no hubiese variabilidad en la respuesta.
3.2 Resultados con RDPs transparentes
Vamos a mostrar primero como se comporta la red MT ante estímulos transparentes
(figura 3.8).
El modelo MT obtenido implementa una normalización de la actividad de la población
manteniéndo su actividad promedio constante cualquiera que sea la forma de estímulo
sensorial que se le presente, RDP simple/RDPs transparentes. Este tipo de normalización
ha sido ya utilizada en otro modelo de MT, aunque impuesta de forma ad hoc (Simoncelli
y Heeger, 1998). De manera que al aumentar la entrada sensorial con los RDPs
transparentes cada vez más separados, la respuesta de la red es disminuir la actividad de
las neuronas que los codifican.
35
Mecanismos de modulación por atención selectiva entre áreas corticales: Un modelo biofísico de red
att.
0
90
180
Figura 3.7: Comparación no atención/atención preferida más 90º. El ajuste a una Gaussiana del caso con
atención a la dirección preferida más 90º es un buen ajuste también para el caso sin atención. La
pequeña diferencia entre ellas proviene de que el factor de modulación para la separación de 90º es un
poco superior a uno (figura 3.3).
Input coming from V1
MT model output
Figura 3.8: Actividad de la red MT ante estímulos RDPs transparentes. En la parte izquierda de la figura,
aparece la entrada sensorial desde V1 a MT para este tipo de estímulos, y en la parte de la derecha, la
actividad de la red en cada caso. Los diferentes colores representan las distintas separaciones entre las
direcciones de los dos componentes del estímulo transparente. La línea negra horizontal y discontinua
representa al valor de la actividad espontánea en la red. Se puede observar que aparece una fuerte
supresión en la actividad máxima de la red a medida que la separación aumenta. No obstante, la
actividad promedio no cambia.
Veamos ahora que esto es lo que ocurre experimentalmente (figura 3.9).
Según las observaciones experimentales (Treue et al., 2000), la repuesta promediada
en MT para RDPs transparentes se ajusta a la suma de dos Gaussianas dividida por dos,
tendencia que el modelo reproduce. Cabe destacar que este tipo de ajuste a dos
Gaussianas mantiene el área bajo la curva, o sea, que la actividad promedio de la
población no es suprimida (sino conservada) aunque sí lo hagan las respuestas máximas
en la línea de (Simoncelli y Heeger, 1998).
36
Capítulo 3: Resultados
De hecho esto no es completamente cierto, puesto que como las Gaussianas tienen un
dominio infinito, el área en una región finita del espacio se calcula a partir de la función
error, de manera que para grandes anchuras de las Gaussianas, no se conservaría el área
en esa región. No obstante, para la anchura promedio observada experimentalmente en
MT, y que el modelo reproduce, la aproximación supuesta es muy buena.
Experiment
MT model output
Figura 3.9: Curvas de respuesta promedio de las neuronas MT ante estímulos RDPs transparentes. En la
parte izquierda de la figura, aparece el promedio de la actividad neuronal en el área MT ante diferentes
separaciones entre componentes de los estímulos RDPs transparentes (N=152) (Treue et al., 2000). En
la parte de la derecha, para comparar se repite la figura para la actividad de la red. Se puede observar
que el modelo reproduce muy bien las tendencias observadas en la actividad neuronal ante este tipo de
estímulos.
Ahora ya vamos a ver como influye la atención cuando lo que tenemos son estímulos
transparentes. Comencemos, otra vez, mostrando dos ejemplos característicos de este
tipo de simulaciones. En ellas se muestran los esquemas de las tareas respectivas (al
estilo de la figura 2.4) y cual es la salida de las dos redes en los dos casos (figura 3.10).
En una situación la atención influye en la respuesta modulando la respuesta sensorial, y
en la otra no, puesto que se atiende a otra dimensión no relacionada con el movimiento.
De la observación directa de estos resultados se puede extraer información:
● En este tipo de tareas con RDPs transparentes todos los estímulos sensoriales caen
dentro de la red MT ya que todos aparecen en el mismo campo receptivo. Por tanto
no es necesaria ninguna entrada externa que emule otras conexiones MT a PFC.
● La actividad sostenida en PFC, en el caso de atención, se consigue con la
superposición, en la primera etapa del ensayo, de la entrada sensorial (RDP simple)
con la señal de alerta o compuerta.
● El caso de no atención no genera actividad sostenida debido a que no coincide la
entrada sensorial simultáneamente con la señal de compuerta.
● Si comparamos la actividad en MT entre los dos casos de atención/no atención, se
observa que las neuronas de la población que codifican la información del RDP
37
Mecanismos de modulación por atención selectiva entre áreas corticales: Un modelo biofísico de red
atendido, y sus vecinas más próximas, incrementan su actividad. En cambio, las
neuronas más alejadas la disminuyen. Esto ocurre a lo largo de las tres etapas de la
tarea: cuando hay un solo RDP simple; cuando no hay más que actividad
espontánea; y cuando el estímulo es transparente. No obstante, con RDPs
transparentes el efecto se hace mucho más notorio, así es fácil ver que el RDP
atendido se ve potenciado mientras que el otro se ve deprimido, justamente en la
linea de a lo que apuntan las tesis de Desimone sobre atención selectiva con
competitividad entre estímulos (Moran y Desimone, 1985; Desimone, 1998).
attention
non-attention
+
t
+
t
+
+
+
rate
55
PFC
location
+
location
55
MT
rate
55
PFC
55
MT
location
location
time (s)
0
time (s)
5
0
5
Figura 3.10: Resultados de dos simulaciones características para RDPs transparentes. En la parte
superior se muestran los diagramas de las tareas, al estilo de la figura 2.4 del capítulo anterior. En la
parte inferior se muestra la actividad de las poblaciones mediante un código de gradiente de color, las
respuestas neuronales más altas aparecen en rojo y las más bajas en azul. Las tres diferentes etapas de
la tarea se separan con líneas verticales negras. La parte izquierda de la figura considera un caso en que
la atención afecta modulando la actividad, mientras que la parte derecha refleja la situación de no
atención.
3.2.1 Modulación por atención con RDPs transparentes
Así como acabamos de ver, para estímulos transparentes se produce una disminución
de la respuesta para las neuronas que codifican al estímulo no atendido y un aumento
para las neuronas cuya respuesta codifican al estímulo sobre el cual recae la atención. El
efecto neto se puede observar mejor en la siguiente figura 3.11.
Recordemos que este tipo de experimento no se ha realizado todavía, de manera que
lo que aquí se muestra es la predicción según el modelo. Una forma de obtener estas
curvas experimentalmente se muestra en el esquema de la figura 3.11.
Pero, ¿Cuál sería la predicción desde el punto de vista de la teoría de Treue? Como se
ha visto en el caso de los estímulos simples, la teoría de Treue se basa en los factores de
modulación generados por la señal de atención. Estos factores dependían de la separación
entre la selectividad de cada neurona con respecto a la señal de atención, y no en la
naturaleza concreta del estímulo. Por tanto, podemos realizar otra clase de predicción, no
la del modelo sino la basada en las propias hipótesis de Treue: partiendo de los factores
multiplicativos que se extraen de la estímulación con un solo RDP (Figura 3.3),
38
Capítulo 3: Resultados
construimos la modulación por atención a la respuesta a estímulos de RDPs transparentes
de la Figura 3.9. Esta sería la predicción del Modelo de Ganancia dependiente de la
Similitud respecto a Característica de Treue. Esto es lo que mostramos en la figura 3.12.
att.
+
no att..
Cue
Neuron
location
Figura 3.11: Modulación por atención de la actividad de la población ante RDPs transparentes. En rojo
tenemos el caso con atención y en azul el caso sin atención. La separación entre los dos componentes del
estímulo es de 120º. Observamos que la modulación influye de manera que el estímulo atendido se
potencia mientras que el otro se deprime. La forma de obtener estas curvas experimentalmente aparece
dibujada esquemáticamente (cuadro superior derecho). La idea es correr todas las posibles direcciones
manteniendo la separación entre los componentes del estímulo fija, y a la vez, la señal de atención
siempre sobre el mismo componente (en rojo en el diagrama).
att.
no att..
prediction
Cue
Neuron
neuron
Figura 3.12: Predicción en base a los factores de modulación. En rojo tenemos el caso con atención, en
azul el caso sin atención y en negro la predicción. La separación entre los dos componentes del estímulo
es de 120º. La curva de predicción se calcula multiplicando todos y cada uno de los puntos de la señal sin
atención por los factores de modulación (figura 3.3), teniendo siempre en cuenta cual es la distancia
entre las direcciones del punto de la curva y la señal de atención.
En esta figura lo que se ha hecho es partiendo de la respuesta del modelo a RDPs
transparentes sin atención (figura 3.9), por un lado, y de los factores de modulación
(figura 3.3) que relacionan los casos atención/no atención para RDPs simples, se ha
39
Mecanismos de modulación por atención selectiva entre áreas corticales: Un modelo biofísico de red
generado una curva que representa el caso de atención para RDPs transparentes. El
proceso pasa por multiplicar punto a punto la curva sin atención por el factor de
modulación pertinente, en base a la separación existente entre la dirección que
representa la correspondiente neurona respecto a la dirección en la cual permanece la
señal de atención.
Como vemos el solapamiento entre la predicción del modelo en base a las simulaciones
biofísicas y la predicción de la teoría de Treue en base a los cálculos de factores de
modulación es casi perfecta, teniendo en cuenta la variabilidad intrínseca del modelo.
Hecho que demuestra que el modelo implementa de forma biológicamente plausible la
modulación de la actividad neuronal mediada por Atención basada en Característica, o
sea, la hipótesis de Treue.
El hecho más significativo del buen ajuste entre predicciones, y por ende, de la teoría
de Treue, es que la modulación no depende del tipo de estímulo, en estos casos RDPs
simples o transparentes, sino únicamente de como difiere la característica preferida de
cada neurona del circuito respecto a la señal de atención.
3.2.2 Modelo de Competición Sesgada
Vamos a mirar la modulación por atención en el caso de RDPs transparentes desde
otra perspectiva. Se ha comentado en el apartado anterior que en este caso, el estímulo
atendido se potencia y el otro se deprime, el mismo efecto observado en las áreas IT y V4
fueron la base para el Modelo de Competición Sesgada de Desimone. La idea principal de
Desimone, es que la atención tiene un papel interesante, sobretodo, cuando hay
competición entre estímulos, siendo ésta altamente selectiva a uno de todos ellos. Por
tanto, su influencia principal actúa durante la competición, acercando la respuesta del
estímulo selecto a la que tendría éste en el caso en que sólo estuviese él sin ningún tipo
de distracción.
Empezaremos viendo la actividad puramente sensorial (sin atención) en coordenadas
polares (figura 3.13). Para después comparar estos casos con los que engloban atención.
+
Figura 3.13: Diagramas polares de los casos no atendidos. En rojo tenemos el caso de RDP simple y en
verde el caso de RDPs transparentes con una separación de 120º entre cada uno de sus componentes. La
curva de la derecha muestra el diagrama en coordenadas polares, ampliamente usado para comparar los
estímulos que están compitiendo.
En la figura 3.14 se muestra en diagramas polares que ocurre cuando la señal de
atención interviene.
40
Capítulo 3: Resultados
No atención
Atención
Figura 3.14: Efecto de la atención desde el punto de vista del Modelo de Competición Sesgada. En el
diagrama polar de la izquierda se compara la respuesta para el caso de RDPs transparentes con las
respectivas respuestas para los RDPs simples que lo componen. En el diagrama de la derecha vemos que
ocurre cuando la señal de atención se aplica a uno u otro de los componentes. Se observa que el
componente atendido ve incrementada la respuesta que evoca acercándose a la que generaría si
estuviese sólo en el campo visual. Esto ocurre a costa de que la respuesta evocada por el otro
componente se reduzca.
Como vemos en la figura 3.14, nuestro modelo implementa el Modelo de Competición
Sesgada de Desimone en el área MT. Ante una competición entre estímulos con atención
focalizada sobre uno de ellos, la respuesta que evoca el estímulo atendido se acerca a la
que tendría éste en el caso en que estuviera solo. Debido a que la red MT mantiene la
actividad promedio, el componente distractor (estímulo no atendido) disminuye su
respuesta, aumentando así la diferencia entre las respuestas evocadas por los dos
estímulos simultáneos.
Y aún más, se acaba de mostrar que el Modelo de Ganancia dependiente de la Similitud
respecto a Característica de Treue y el Modelo de Competición Sesgada de Desimone son
las dos caras de una misma moneda cuando en un mismo campo receptivo hay
competencia entre estímulos (Boynton, 2005). Nuestro modelo es el primer circuito
biofísicamente plausible que es capaz de reconciliar estas dos ideas dentro de una
arquitectura consistente con nuestros conocimientos acerca de cómo se organiza y
estructura la corteza cerebral.
3.3 Mecanismos
En este momento estamos abordando el estudio en detalle de los mecanismos que
hacen posible todos los resultados del modelo mostrados en los apartados anteriores. Por
tanto, lo que ahora se va a mostrar forma parte de resultados aún preliminares.
Destaca que en la red MT predomina fuertemente la corriente sináptica inhibidora,
como se observa en las figuras 3.15 y 3.16.
41
Mecanismos de modulación por atención selectiva entre áreas corticales: Un modelo biofísico de red
Figura 3.15: Corrientes promedio entrantes a las neuronas excitadoras en MT en ausencia de estímulo
sensorial. Vemos que en cuanto a corrientes sinápticas recurrentes, la inhibidora es la que predomina.
Para compensar el efecto y obtener la actividad observada experimentalmente (~9 Hz) necesitamos una
alta entrada externa.
Figura 3.16: Corriente sináptica recurrente total promedio entrante a las neuronas excitadoras en MT en
presencia de estímulo sensorial con y sin señal de atención. Vemos que la contribución neta es inhibidora
y que aumenta con la señal de atención.
La inhibición aumenta a medida que aumentan las diferentes entradas excitadoras,
tanto sensoriales como de atención. Esta característica de la red MT es la que hace
posible que la actividad de la población permanezca aproximadamente constante. A
mayores entradas sensoriales, mayor grado de inhibición, la cual compensa el efecto neto
a lo largo de la toda la red, de manera que la actividad localmente aumenta pero no en
promedio, así en las zonas alejadas al estímulo la respuesta está por debajo de la
actividad espontánea. Cuando comparamos los casos atención/no atención, un argumento
42
Capítulo 3: Resultados
similar permite explicar porqué se produce el cruce de las curvas de sintonía de la
población.
Este comportamiento se ve acompañado de la disminución de la anchura de la
respuesta (figura 3.17). Se puede observar, en cambio, que para el caso de atención la
anchura aumenta respecto al caso de no atención. Esto se debe a que la conexión a MT
proveniente de PFC es más ancha que la entrada sensorial, hecho que aumenta
ligeramente la anchura resultante de la respuesta en MT.
Figura 3.17: La anchura de las respuestas decrece respecto a la entrada sensorial. Esta anchura se
calcula como la de la Gaussiana que mejor se ajusta a cada curva.
Se ha propuesto que mecanismos de tipo celular podrían ser capaces de generar el
efecto multiplicativo mediante determinados cambios internos a la célula, frente a la otra
posibilidad que es que el cambio se de en las corrientes sinápticas que le llegan. Existen
en la literatura dos tipos de mecanismos celulares propuestos que se basan en la relación
existente entre la corriente sináptica total entrante a una determinada neurona y su tasa
de disparo. Este tipo de relación se suele representar gráficamente y se suele citar como
curva fI (firing rate versus Input). En uno de los dos mecanismos, la curva fI es no lineal,
en concreto, la relación cumple una ley de potencias, de manera que un mismo
incremento aditivo en la corriente entrante genera, a diferentes tasas de disparo,
modulaciones que resultan multiplicativas entre ellas, y que estan mediadas por la ley de
potencias (Murphy y Miller, 2003) (figura 3.18 A). El otro mecanismo hace referencia a
cuando dicha relación es lineal pero su pendiente cambia entre las dos situaciones a
comparar, de manera que aumenta la ganancia de la curva fI y se obtiene a su vez el
efecto multiplicativo en la respuesta (Chance et al., 2002) (figura 3.18 B).
Pero como demuestra las figura 3.19, ninguno de estos mecanismos parece ser el
responsable, puesto que las curvas fI son muy poco no-lineales y la pendiente de estas
curvas aunque cambia entre los diferentes casos, este cambio es demasiado pequeño
para ser por si solo el mecanismo responsable del efecto multiplicativo (cambio máximo
entorno al 6'5 %).
Se ha de estudiar más a fondo cual es realmente el mecanismo que sustenta el efecto
multiplicativo observado, pero todo apunta a que se trata de un nuevo mecanismo no
aparecido aún en la literatura, no de tipo celular sino de red, basado en la estructura de
las conectividades recurrentes y en el fuerte predominio de la inhibición.
43
Mecanismos de modulación por atención selectiva entre áreas corticales: Un modelo biofísico de red
A
B
FR
FR
I
I
Figura 3.18: Mecanismos celulares propuestos para la obtención de ganancias multiplicativas. A. Relación
según una ley de potencias en la curva Corriente-Respuesta (fI). Un mismo cambio en la corriente genera
un cambio diferente en la respuesta, el cual es multiplicativo (observar como la diferencia entre las lineas
verticales azul y roja son iguales en corriente pero no en la respuesta) (Murphy y Miller, 2003). B. Las
curvas fI son lineales, lo que cambia es la pendiente entre los casos a comparar, resultando así un efecto
de ganancia multiplicativa (Chance et al., 2002).
Neuron
FR (Hz)
FR (Hz)
I (nA)
I (nA)
Figura 3.19: Relación altamente lineal entre la tasa de disparo y la corriente total entrante para todas las
diferentes situaciones de atención. Los mismos efectos observados en las curvas de sintonía de la
población, cruce (arriba izquierda: en rojo el caso de atención y en azul el caso sin atención) y efecto
multiplicativo (arriba derecha: atención en la dirección preferida en rojo, antipreferida azul y a 90º
verde), se observan también con la corriente total sináptica entrante. A la vez, las curvas fI en todas las
situaciones indican que existe una relación lineal entre la respuesta y la corriente de entrada, y que esta
es la misma independientemente de la condición de atención en la que se esté. Se demuestra así que el
mecanismo responsable del efecto multiplicativo en el modelo no es ninguno de los mecanismos celulares
anteriormente descritos. Probablemente se trate de un nuevo mecanismo que se base en la estructura de
red no aparecido antes en la literatura.
44
Capítulo 4: Discusión
En este trabajo, nuestro principal objetivo ha sido el estudio de la modulación de la
actividad neuronal durante tareas de atención selectiva, centrándonos en la Atención
basada en Característica y poniendo especial cuidado en evaluar las modificaciones sobre
la selectividad de las neuronas y de las poblaciones de neuronas que induce la
focalización de la atención. Para ello, se ha elaborado un modelo computacional que tiene
en cuenta los aspectos conocidos más relevantes, y a la vez más contrastados, de este
tipo de atención según la literatura previa. De acuerdo con esta información, nuestra
principal hipótesis para la implementación del modelo ha sido la necesidad de utilizar dos
redes corticales interrelacionadas en forma de bucle, una de las cuales describe el área
sensorial y la otra es capaz de generar actividad persistente, como supuestamente
ocurriría en un área ejecutiva. Pero además, dado que lo que más nos ha interesado no
es tanto reproducir experimentos, sino ir más allá para comprender la base biológica que
subyace a esta fenomenología, hemos considerado como requisito imprescindible utilizar
un modelo de neurona y de red que sea razonable desde el punto de vista de la
electrofisiología, sin ser, a su vez, demasiado complejo, lo cual dificultaría la
interpretación de los resultados. Así, nos hemos restringido a una región cortical donde
los efectos de modulación por la atención han sido expuestos de forma clara y
significativa, el área MT, y hemos supuesto que la actividad persistente asociada se
localiza en la corteza prefrontal o parietal posterior. De manera que éstas han sido las
áreas que se han representado en nuestro modelo de forma plausible, las cuales serían
las responsables de la generación de las señales 'Bottom-up', o señal sensorial, y 'Topdown', o señal de atención, respectivamente.
Como hemos visto en la introducción, la Atención basada en Característica genera dos
tipos de modulación distintas según la atención esté fuera, o dentro, de un determinado
campo receptivo (Atención basada en Característica Global y Atención basada en
Característica Local, respectivamente). Como ya se ha comentado, esto ha generado la
aparición de dos teorías independientes (Modelo de Ganancia dependiente de la Similitud
respecto a Característica y Modelo de Competición Sesgada, respectivamente) y,
consecuentemente, cierta controversia entre ellas. Así en principio, estas distintas
modulaciones podrían recaer sobre diferentes mecanismos. Para poder estudiar si esto es
así, o si en realidad se pueden explicar bajo el mismo mecanismo, se han modelizado dos
diferentes paradigmas, uno con estímulos simples frente al otro que considera estímulos
transparentes.
Mecanismos de modulación por atención selectiva entre áreas corticales: Un modelo biofísico de red
Dentro del primer paradigma (estimulación simple y atención fuera del campo
receptivo), el primer paso ha sido reproducir el cruce de las curvas de sintonía de la
población, obtenido por Treue, entre el caso en que atención y estímulo van ligados,
respecto al caso sin atención. En realidad, este cruce en las curvas de sintonía resultó en
su momento bastante controvertido, e incluso contradictorio, pues anteriormente la
modulación basada en característica obtenida había aparecido como un efecto
multiplicativo entre curvas de sintonía, en áreas como V4 (McAdams y Maunsell, 1999), o
en MT por el mismo Treue (Treue y Martinez-Trujillo, 1999). La interpretación propuesta
por Treue fue que existe un efecto multiplicativo a nivel neuronal, que depende sólo de la
separación entre la característica del estímulo atendido y la dirección preferida de la
neurona, así cuando construimos una curva de sintonía desplazando el foco de atención
con el estímulo existirá un factor diferente sobre la respuesta neuronal para cada posición
del foco de atención, en unos casos mayor que la unidad y en otros menor, y esta
modulación del factor multiplicativo sería la responsable del cruce (Martinez-Trujillo y
Treue, 2004). No obstante, en ese artículo, no se llegaron a explorar las curvas de
sintonía completas cuando la atención permanecía fija respecto a la selectividad de la
neurona, y sólo investigaron los casos extremos, pero en estos se obtenían efectivamente
relaciones multiplicativas. En nuestro modelo, se ha llevado esta hipótesis hasta las
últimas consecuencias mostrándose el efecto multiplicativo de las curvas de sintonía para
atención fija.
Conviene recordar que consideramos curva de sintonía a la representación gráfica de la
respuesta de una única neurona respecto al cambio de valor de una deteminada
dimensión del estímulo, manteniendo todo lo demás en condiciones constantes.
Tradicionalmente, se ha obtenido que las curvas de sintonía son multiplicativas por
atención porque se ha mantenido constante, o fija, la característica atendida. En cambio,
en los casos en que la atención es intrínseca al estímulo, como cuando atención y
estímulo siempre van ligados, podremos aún hablar de curva de sintonía pero hay que
diferenciar muy claramente las dos situaciones. En este caso, la característica atendida
junto al estímulo sensorial cambia en cada punto y, por tanto, la modulación obtenida es
el refinamiento de la selectividad y no la modulación multiplicativa. Este tipo de curva de
sintonía ha dado en llamarse actividad de la población o curva de sintonía de la población,
porque es equivalente a la respuesta que tendrían las neuronas de una población frente a
un único de estos estímulos atendidos (Martinez-Trujillo y Treue, 2004). Esta equivalencia
ha sido comprobada en el modelo, de forma que con una única simulación se ha obtenido
el cruce de las curvas de sintonía de la población.
Desde el punto de vista del comportamiento, lo que estos resultados demuestran es
que la modulación por atención resulta en un refinamiento de la selectividad de la
población (en inglés, sharpening), efecto que tantas veces se ha propuesto desde el
campo de la psicofísica, y no en un aumento multiplicativo de la actividad de todas las
neuronas que la forman, efecto que se ha venido proponiendo desde la electrofisiología.
Esto es así porque, en situaciones de relevancia comportamental el sujeto tiene que
tomar decisiones en base a los estímulos sensoriales instantáneos, los cuales toman
solamente una determinada característica como valor para cada dimensión, sobre la que
puede recaer la atención. Por lo tanto, el sistema no puede computar una curva de
sintonía presentando todas las características posibles con la atención prefijada sino que
tiene que valorar la actividad instantánea de la población de neuronas. Como hemos visto
en la población de neuronas, cada neurona del campo receptivo tendrá, en el caso de
atención, una modulación que dependerá de la relación entre la característica atendida
respecto a su característica preferida, resultando en un refinamiento de la selectividad de
la población.
46
Capítulo 4: Discusión
Así como en el primer paradigma el modelo básicamente reproduce resultados
experimentales (aunque también es cierto que los complementa, mediante la obtención
de las curvas de sintonía para el caso de atención fija, hecho que apoya el Modelo de
Ganancia dependiente de la Similitud respecto a Característica), en el segundo paradigma
el modelo predice, por un lado, la confirmación de la teoría de Treue y, por otro lado, el
modo en que se resuelve la controversia entre ésta y el Modelo de Competición Sesgada,
demostrando que ambos esquemas difieren en el punto de vista pero no en su base
mecanística. Este segundo paradigma experimental considera estímulos simultáneos en
un mismo campo receptivo, los cuales entran en competición. De manera que se cumplen
todas las bases del Modelo de Competición Sesgada. Explicado brevemente, lo que se
obtiene es una supresión en la actividad cuando se presentan los estímulos preferido y
antipreferido simultáneamente en relación al preferido solo; y la atención modula esta
actividad desplazándola hacia la que tendría si no existiera competencia entre los
estímulos. A la vez, la respuesta del modelo se puede reproducir modulando la obtenida
en el caso sin atención con los mismos factores multiplicativos obtenidos en el primer
paradigma (figura 3.12). Esto implica que para el Modelo de Ganancia dependiente de la
Similitud respecto a Característica, desde el punto de vista de los mecanismos, resulta
indiferente que la atención esté dentro o fuera del campo receptivo y, al mismo tiempo,
no importa si el estímulo sensorial se basa en movimiento unidireccional o bidireccional
transparente. Cabe recalcar que los resultados del modelo, bajo este paradigma, se basan
en el hecho de que la red MT reproduce de forma exitosa la respuesta puramente
sensorial ante estimulación con RDPs con movimiento transparente, que supone de forma
efectiva una normalización de la actividad de la población (figura 3.9).
Como se ha comentado en otras secciones anteriores, ha aparecido recientemente un
modelo matemático muy interesante capaz de reproducir el cruce de las curvas de
sintonía a partir del efecto multiplicativo propuesto por el Modelo de Ganancia
dependiente de la Similitud respecto a Característica, y donde se relaciona esto con las
bases del Modelo de Competición Sesgada (Boynton, 2005). No obstante, este modelo
está implementado de forma completamente ad hoc, sin tener en cuenta posibles bases
biológicas, de manera que no se puede extraer información respecto al modo en que esto
se podría implementar en el cerebro. Nuestro modelo aporta este tipo de información,
puesto que está basado desde un principio en los mecanismos fisiológicos que operan en
la corteza cerebral.
En realidad, nuestro estudio no está ni mucho menos cerrado. Como se ha visto en la
parte de mecanismos del capítulo de resultados, necesitamos aún estudiar a fondo cuál es
la base fisiológica que sustenta el efecto multiplicativo del Modelo de Ganancia
dependiente de la Similitud respecto a Característica en nuestra implementación
computacional. No obstante, a partir de los análisis realizados podemos extraer cierta
información, como que no parece que se trate de un mecanismo de origen celular, ni del
tipo propuesto por Miller (Murphy y Miller, 2003) ni del tipo propuesto por Abbott (Chance
et al., 2002). Probablemente se trate de un mecanismo de red no aparecido previamente,
donde tendrá un peso específico el hecho de que en la red MT predomina fuertemente la
inhibición recurrente de modo que la red normaliza sus respuestas para mantener la
actividad promedio aproximadamente constante.
Me gustaría hacer notar que nuestro modelo únicamente considera una dimensión del
estímulo, la dirección de movimiento, de manera que será incapaz de generar
modulaciones frente a otras características si éstas existieran. Cabe recordar, como se
comentó brevemente en el capítulo de introducción, que la localización espacial podía ser
interpretada como una característica más, aunque a día de hoy esto sea bastante
controvertido. Así pues, nuestro modelo no producirá directamente modulación espacial
cuando la atención pase de estar fuera del campo receptivo a estar dentro. Es decir, si
47
Mecanismos de modulación por atención selectiva entre áreas corticales: Un modelo biofísico de red
por ejemplo considerásemos la comparación entre la actividad para el caso de un RDP
simple cuando la atención pasa de estar fuera del campo receptivo a estar dentro, no
obtendríamos en el modelo diferencia alguna. Como decíamos, este tipo de experimentos
ha generado discrepancias en cuanto al tipo de modulación resultante. Algunos
experimentos muestran un efecto multiplicativo de la actividad (McAdams y Maunsell,
1999; Treue y Martinez-Trujillo, 1999), lo cual estaría de acuerdo con la consideración de
la localización como otra característica más (Treue y Martinez-Trujillo, 1999). Mientras
otros experimentos muestran un aumento de la actividad únicamente a niveles
intermedios de contraste, lo que supone el aumento del contraste efectivo pero
únicamente para este rango de contrastes intermedios (Reynolds et al., 2000; MartinezTrujillo y Treue, 2002). Esto, en principio, parecería estar en desacuerdo con suponer la
localización como característica, ya que no existiría efecto multiplicativo, y argumentaría
en favor de que se tratase de dos sistemas paralelos y diferenciados, tal como se ha
propuesto en un modelo matemático esquemático (Boynton, 2005). De todos modos, el
uso de RDPs transparentes es idóneo para evitar el hecho de tener que considerar el
efecto espacial en la modulación por atención, ya que para este tipo de estímulo no existe
separación espacial alguna. En cambio con otros tipos de estímulos, como los basados en
imágenes usados tradicionalmente en paradigmas de competición, la diferencia de
localización tiene un efecto neto en la actividad por atención, aunque este efecto
disminuye en gran medida, pero no completamente, si las imágenes están ambas dentro
del mismo campo receptivo.
Dado que, como se ha indicado en el capítulo de métodos, la entrada sensorial al área
MT proviene de V1, y ésta genera un tipo de normalización que parece ser dependiente
del contraste, esperamos en un futuro próximo poder ampliar el estudio en el modelo con
este tipo de efectos de la atención sobre el contraste, mediante la incorporación de otro
módulo de red cortical que represente el procesamiento de la información en área V1. Ya
he avanzado en esta línea de trabajo desarrollando un modelo de normalización en área
visual primaria como proyecto durante mi participación en el curso de verano patrocinado
por IBRO y FENS: Advanced Course in Computational Neuroscience. Agosto 1-26, 2005,
Arcachon, Francia. Probablemente se haga también necesaria otra ampliación del modelo
para considerar posibles efectos de la localización espacial como característica.
Existen dos importantes efectos dependientes de la atención que han aparecido
recientemente en la literatura y que pueden ser estudiados bajo el prisma de nuestro
modelo con el objetivo de marcar nuevas predicciones en el área MT. El primero es que se
ha observado un aumento de la sincronía, para frecuencias gamma (35-90 Hz), en
neuronas de V4 activadas por el estímulo atendido, en comparación con neuronas vecinas
activadas por el estímulo distractor. Se ha argumentado que esto podría ser útil en la
corteza cerebral para amplificar señales relevantes desde el punto de vista del
comportamiento (Fries et al., 2001). Resultados preliminares en nuestro modelo marcan
la misma tendencia en la red MT del modelo, si existen ciertas oscilaciones en las
neuronas que codifican el estímulo atendido en el circuito prefrontal. Exploraremos más a
fondo estos aspectos, incluyendo la relación que pudiese establecerse entre los dos
módulos del bucle (áreas MT y PFC) cuando ambos son capaces de desarrollar actividad
oscilatoria intrínseca.
El segundo aspecto que se quiere considerar es el desplazamiento (en inglés, shift) del
centro de la curva de sintonía provocado por la señal de atención. Existen evidencias
experimentales de que no únicamente se produce un aumento de la actividad en
situaciones de atención selectiva, sino que también existe un desplazamiento de los
campos receptivos efectivos hacia el estímulo atendido cuando éste se mantiene cercano
(Connor et al., 1996; Connor et al., 1997). Además, un modelo teórico aparecido
recientemente reproduce esta tendencia del desplazamiento en la actividad de la
48
Capítulo 4: Discusión
población hacia el estímulo atendido. No obstante, predice que la curva de sintonía de las
neuronas sufrirán un desplazamiento que dependerá de si en la población predomina un
aumento de su actividad, o un desplazamiento de su campo receptivo. De manera que,
en la curva de sintonía, el desplazamiento resultante que se generará será un
acercamiento o un alejamiento, respectivamente (Compte y Wang, 2005). Nuestro
modelo es ideal para este estudio, pues, a nivel de población, nos ha permitido
contrastarlo con observaciones experimentales obtenidas en otras áreas, lo cual es una
predicción de lo que puede ocurrir en MT, donde aún no se han estudiado los
desplazamientos de las curvas de sintonía por atención. Un estudio más detallado en un
futuro próximo, nos permitirá predecir que tipo de desplazamiento se da en el área MT a
nivel neuronal, si hacia o en contra del estímulo atendido. Además, debido a que nuestro
modelo tiene una mayor base fisiológica en comparación con este otro modelo
recientemente publicado, los resultados que podamos obtener en este sentido permitirán,
de alguna manera, contrastar las hipótesis de éste.
Además, en un futuro nos planteamos abordar los aspectos que relacionen la actividad
en nuestro modelo de bucle de áreas corticales con aspectos funcionales y conductuales
en humanos y monos en comportamiento. Para ello trabajaremos en dos frentes,
relacionando nuestro modelo con estudios realizados con humanos con técnicas de
imagen, por un lado; y los estudios psicofísicos, por otro. En cuanto a la relación de
nuestro modelo con las técnicas de imagen, nuestra intención es comparar medidas fRMI
sobre la actividad del área MT en tareas de Atención basada en Característica (Saenz et
al., 2002), con la actividad de la población de nuestra red MT. Para poder comparar
directamente estas medidas, necesitamos tener en cuenta cómo se relaciona primero la
respuesta hemodinámica, o vascular, con la actividad de las neuronas. Se sabe que esta
respuesta aparece retardada del orden de 5-8 s, que es transitoria y que se dispersa con
el tiempo (Bandettini et al., 1993; Friston et al., 1994). Se ha comprobado que una
función de distribución de Poisson representa de forma consistente esta latencia de la
respuesta vascular (Friston et al., 1994; Horwitz et al., 2001). De manera que nuestra
aproximación se basará en la integración del valor absoluto de la actividad sináptica
obtenida en la red MT y su convolución posterior con la dsitribución de Poisson. Este
método ya ha sido simulado en un modelo computacional similar del área V4 en tareas de
atención (Corchs y Deco, 2004).
Respecto a la relación con los estudios psicofísicos, cabe mencionar dos cosas: la
primera, que el grupo alemán con el que colaboramos, encabezado por el profesor Stefan
Treue, está realizando este tipo de experimentos tanto con monos, como con sujetos
humanos. La investigación es llevada a cabo en el centro de primates por el investigador
experimentalista Vladislav Kozyrev, con el cual he tenido la oportunidad de colaborar en
dos ocasiones, con sendas visitas a Göttingen, para el diseño de las tareas a realizar;
para la obtención de los primeros datos experimentales con sujetos humanos; y en el
posterior análisis de los datos, obtención y representación gráfica de las funciones
psicométricas. Por tanto, es obvio que nos puede resultar de gran utilidad realizar
comparaciones entre los resultados de estos experimentos y los que podamos obtener de
nuestro modelo computacional, predicción de lo que pueda pasar en las neuronas de MT
cuando se empiece a registrar.
La segunda relación de nuestro modelo con la psicofísica deriva de experimentos ya
realizados. De hecho, ya han sido descritas determinadas propiedades de estas funciones
psicométricas neuronales en el área MT para determinadas tareas de discriminación
(Britten et al., 1992; Purushothaman y Bradley, 2005). Los resultados entre estas
publicaciones son en cierto sentido contradictorias, así según Britten existen neuronas
individuales en MT que tienen un comportamiento igual o superior al del propio mono, es
decir son capaces de discernir con un menor umbral, un determinado movimiento
49
Mecanismos de modulación por atención selectiva entre áreas corticales: Un modelo biofísico de red
coherente inmerso en movimientos aleatorios. En cambio, Purushothaman obtiene que las
neuronas por sí solas discriminan mucho peor en término medio que el animal. La tarea
que utiliza es responder si un estímulo coherente se mueve inclinado a la derecha, o a la
izquierda, de otro presentado anteriormente que se movía de forma totalmente vertical.
Aunque todas las neuronas registradas tengan peor poder de discriminación existen
algunas neuronas concretas que discriminan mejor que otras. Por tanto, propone que es
la población conjunta de estas neuronas con mayor capacidad de discriminar la que
consigue un mayor refinamiento y obtener un comportamiento similar al del animal en su
conjunto. En cuanto a las diferencias entre un experimento y otro, argumenta que se
utilizan tareas diferentes, que las células que Britten ha utilizado en su experimento han
estado preseleccionadas como las más discriminatorias, y que la diferencia entre las
direcciones a discriminar en su tarea son tan pequeñas que son imposibles de discriminar
por una única neurona, desde el punto de vista de la teoría de la estimación, dada la
anchura de su curva de sintonía.Nuestro modelo puede ser testeado directamente para
estos dos tipos de tarea para ver en qué casos es suficiente la capacidad discriminatoria
de una sola neurona y cuándo se requiere un refinamiento de la selectividad de la
población, al estilo sharpening, comentado antes en esta discusión. (En la tarea de
Britten se ha de introducir la coherencia en la entrada sensorial de V1 a MT para generar
el estímulo ruidoso. No obstante, el estímulo que hemos modelizado incorpora ya esta
dependencia con la coherencia, aunque en nuestros resultados previos se haya obviado
por ser ésta siempre el 100%).
Comentar que estamos elaborando un póster sobre el modelo realizado, el cual vamos
a presentar en el próximo congreso: Annual Meeting of the American Society for
Neuroscience 12-16 de Noviembre, Washington DC, al mismo tiempo que estamos
escribiendo nuestros resultados en un artículo para publicarlos próximamente.
Queremos aprovechar de este modo todo el potencial del modelo que tenemos entre
manos, relacionando aspectos mecanísticos que pueden ser desde la participación de un
determinado receptor, la constante de tiempo de una sinapsis, u otras propiedades de
carácter sináptico o celular, con aspectos funcionales de la actividad electrofisiológica de
neuronas y poblaciones en cada área, hasta actividades globales que puedan registrarse
en experimentos de imagen y aspectos perceptuales que se manifiesten en experimentos
de psicofísica. La posibilidad única de integrar todos estos niveles tan distanciados a nivel
experimental es una propiedad muy atractiva de nuestro modelo que explotaremos para
mi trabajo de tesis.
Agradecimientos
Quisiera concluir el trabajo agradeciendo, en primer lugar, a mi tutor la orientación y
apoyo que me ha otorgado en todo momento, sin los cuales este trabajo no hubiera sido
posible. Así como a los investigadores con los que he tenido el gusto de colaborar y
discutir mi modelo ( Xiao-Jing Wang del grupo de Estados Unidos; y Stefan Treue y
Vladislav Kozyrev del grupo de Alemania); y especialmente a Alfonso Renart por sus
inestimables explicaciones y sugerencias en la aproximación mean-field. Gracias a sus
contribuciones el modelo ha evolucionado en gran medida. Y a todos mis familiares y
amigos más cercanos porqué siempre aparecen cuando hace falta.
Este trabajo ha sido posible gracias al apoyo financiero de la Fundación Volkswagen, al
Ministerio de Educación y Ciencia (BFI2002-02378) y al Fondo Europeo de Desarrollo
Regional.
50
Referencias
Abeles M. (1991) Corticonics. Cambridge University Press, New York.
Albright T. D. (1984) Direction and orientation selectivity of neurons in visual area
MT of the macaque. Journal of Neurophysiology, 52:1106-1130.
Bandettini P. A., Jesmanowicz A. J., Wong E. C., Hyde J. S. (1993) Processing
strategies for time-course data sets in functional MRI of the human brain.
Magnetic Resonance in Medicine, 30:161–173.
Bauer R. H., Fuster J.M. (1976) Delayed-matching and delayed-response deficit
from cooling dorsolateral prefrontal cortex in monkeys. Journal of
Comparative and Physiological Psychology, 90:293-302.
Beauchamp M. S., Cox R. W., DeYoe E. A. (1997) Graded effects of spatial and
featural attention on human area MT and associated motion processing
areas. Journal of Neurophysiology, 78:516-520.
Boynton G. M. (2005) Attention
Neurobiology, 15:465-469.
and
visual
perception.
Current
Opinion
in
Born R. T., Bradley D. C. (2005) Structure and function of visual area MT. Annual
Review of Neuroscience, 28:157-189.
Braitenberg V., Schütz A. (1991) Anatomy of the Cortex. Springer-Verlag, Berlin.
Britten K. H., Shadlen M. N., Newsome W. T., Movshon J. A. (1992) The analysis of
visual motion: a comparison of neuronal and psychophysical performance.
Journal of Neuroscience, 12:4745-65.
Britten K. H., Shadlen M. N., Newsome W. T., Movshon J. A. (1993) Responses of
neurons in macaque MT to stochastic motion signals. Visual Neuroscience,
10:1157-1169.
Mecanismos de modulación por atención selectiva entre áreas corticales: Un modelo biofísico de red
Britten K. H., Newsome W. T. (1998) Tuning bandwidth for near-threshold stimuli in
area MT. Journal of Neurophysiology, 80:762-770.
Bruce C. J., Goldberg M. E. (1985) Primate frontal eye fields. I. Single neurons
discharging before saccades. Journal of Neurophysiology, 53:603-635.
Cavada C., Goldman-Rakic P. S. (1989) Posterior parietal cortex in rhesus monkey:
II. Evidence for segregated cortico-cortical networks linking sensory and
limbic areas with the frontal lobe. Journal of Comparative Neurology, 287:422445.
Chafee M. V., Goldman-Rakic P. S. (1998) Matching patterns of activity in primate
prefrontal area 8a and parietal area 7ip neurons during a spatial working
memory task. Journal of Neurophysiology,79:2919-2940.
Chafee M. V. Goldman-Rakic P. S. (2000) Inactivation of parietal and prefrontal
cortex revealsinterdependence of neural activity during memory-guided
saccades. Journal of Neurophysiology, 83:1550-1566.
Chance F. S., Abbott L. F., Reyes A. D. (2002) Gain modulation from background
synaptic input. Neuron, 35:602.4.
Chelazzi L., Desimone R. (1994) Responses of V4 neurons during visual research.
Society for Neuroscience Abstract, 20:1054.
Chelazzi L., Miller E. K., Duncan J., Desimone R. (1993) A neural basis for visual
search in inferior temporal cortex. Nature, 363:345.7.
Chelazzi L., Duncan J., Miller E. K., Desimone R. (1998) Responses of neurons in
inferior temporal cortex during memory-guided visual search. Journal of
Neurophysiology, 80:2918-2940.
Cohen J. D., Perlstein W. M., Braver T. S., Nystrom L. E., Noll D. C., Jonides J., Smith E.
E. (1997) Temporal dynamics of brain activation during a memory working
task. Nature, 386:604.8.
Compte A., Brunel N., Goldman-Rakic P. S., Wang X-J. (2000) Synaptic mechanisms
and network dynamics underlying spatial working memory in a cortical
network model. Cerebral cortex, 10:910-923.
Compte A., Wang X-J. (2005) Tuning curve shift by attentional modulation in
cortical neurons: a computational study of its mechanisms. Cerebral cortex,
doi:10.1093/cercor/bhj021 (in press, published online).
Connor C. E., Gallant J. L., Preddie D. C., Van Essen D. C. (1996) Responses in area V4
depend on the spatial relationship between stimulus and attention. Journal
of Neurophysiology, 75:1306.8.
Connor C. E., Preddie D. C., Gallant J. L., Van Essen D. C. (1997) Spatial attention
effects in macaque area V4. Journal of Neuroscience, 17:3201-3214.
52
Referencias
Constantinidis C., Franowicz M. N., Goldman-Rakic P. S. (1999) Multiple electrode
analysis of local circuitry in the primate prefrontal cortex during spatial
working memory. Society for Neuroscience Abstract, 25:44-.1.
Corbetta M. Shulman G. L. (2002) Control of goal-directed and stimulus-driven
attention in the brain. Nature Reviews Neuroscience, 3:201-215.
Corchs S., Deco G. (2004) Feature-based attention in human visual cortex:
simulation of fRMI data. NeuroImage, 21:36-45.
Courtney S M., Ungerleider L. G., Keil K., Haxby J. V. (1997) Transient and sustained
activity in a distributed neural system for human working memory. Nature,
386:608-611.
Courtney S M., Petit L., Maison J. M., Ungerleider L. G., Haxby J. V.( 1998) An area
specialized for spatial working memory in human frontal cortex. Science,
279:1347:1351.
Desimone R. (1996) Neural mechanisms for visual memory and their role in
attention. Proc. Natn. Acad. Sci. USA, 93:13494.9.
Desimone R. (1998) Visual attention mediated by biased competition in
extrastriate visual cortex. Philosophical Transactions of the Royal Society of
London. Series B. Biological Sciences, 353:1245-1255.
Desimone R., Duncan J. (1995) Neural mechanisms of selective visual attention.
Annual Review of Neuroscience, 18:193-222.
Desimone R., Wessinger M., Thomas L., Schneider W. (1990) Attentional control of
visual perception: cortical and subcortical mechanisms. Cold Spring Harbor
Symposium on Quantitative Biology, 55:963-971
D'Esposito M., Aguirre G. K., Zarahn E., Ballard D., Shin R. K., Lease J. (1998)
Functional MRI studies of spatial and nonspatial working memory. Cognitive
Brain Research, 7:1-13.
Distler C., Boussaoud D., Desimone R., Ungerleider L. G. (1993) Cortical connections
of inferior temporal area TEO in macaques. Journal of Comparative Neurology,
334:125-150.
Duncan J. (1996) Cooperating brain systems in selective perception and action.
In: Attention and performance XVI (Inui T., McClelland J. L. Eds.), pp 549-578.
Cambridge, MA: MIT Press.
Duncan J., Nimmo-Smith I. (1996) Objects and attributes in divided attention:
Surface and boundary systems. Percepteption & Psychophysics, 58:1076-1084.
Felleman D. J., Van Essen D. C. (1991) Distributed hierarchical processing in the
primate cerebral cortex. Cerebral Cortex, 1:1-47.
Fries W., Keizer K., Kuypers H. G. J. (1985) Large layer V1 cells in macaque striate
cortex (Meynart cells) project to both superior colliculus and prestriate
visual area V5. Experimental Brain Research, 58:613.6.
53
Mecanismos de modulación por atención selectiva entre áreas corticales: Un modelo biofísico de red
Fries P., Reynolds J. H., Rorie A. E., Desimone R. (2001) Modulation of oscillatory
neuronal synchronization by selective visual attention. Science, 291:1560.3.
Friston K. J., Jezzard P., Turner R. (1994) Analysis of functional MRI time-series.
Human Brain Mapping, 1:153–171.
Funahashi S., Bruce S. J., Goldman-Rakic P. S. (1989) Mnemonic coding of visual
space in the monkey's dorsolateral prefrontal cortex. Journal of
Neurophysiology, 61:1-19.
Funahashi S., Bruce C. J., Goldman-Rakic P. S. (1993) Dorsolateral prefrontal lesions
and oculomotor delayed response performance: evidence for mneumonic
'scotomas'. Journal of Neurosciences, 13:1479-1497.
Fuster J. M. (1973) Unit activity in prefrontal cortex during delayed-response
performance: Neuronal correlates of transient memory. Journal of
Neurophysiology, 36:61-78.
Fuster J. M. (1988) The prefrontal cortex. New York: Raven Press.
Fuster J. M. (2001) The prefrontal cortex:An update: Time is of the essence.
Neuron, 30:319-333.
Fuster J. M., Alexander G. E. (1971) Neuron activity related to short-term memory.
Science, 173:652.4.
Fuster J. M., Jervey J. P. (1982) Neuronal firing in the inferotemporal cortex of the
monkey in a visual memory task. Journal of Neuroscience, 2:361-375.
Gnadt J. W., Andersen R. A. (1988) Memory related motor planning activity in
posterior parietal cortex of macaque. Experimental Brain Research, 70:216-220.
Goldman-Rakic P. S. (1987) Circuitry of primate prefrontal cortex and regulation of
behavior by representational memory. In: Handbook of physiology, vol. V: The
nervous system (Plum F., Mountcastle V. B., Eds.), chap. 9, pp. 373-417. Bethesda
MD: American Physiological Society.
Goldman-Rakic P. S. (1995) Cellular basis of working memory. Neuron, 14:477-485.
Goldman P. S., Rosvold H. E. (1970) Localization of function within the dorsolateral
prefrontal cortex of the rhesus monkey. Experimental Neurology, 27:291-304.
Hansel D., Mato G., Meunier C., Neltner (1998) On numerical simulations of
integrate-and-fire neural networks. Neural Computation, 10:467-483.
Hawken M. J., Parker A. J., Lund J. S. (1988) Laminar organization and contrast
sensitivity of direction-selective cells in the striate cortex of the Old World
monkey. Journal of Neuroscience, 8:3541.8.
Hestrin S., Sah P., Nicoll R. (1990) Mechanisms generating the time course of dual
component excitatory synaptic currents recorded in hippocampal slices.
Neuron, 5:247-253.
54
Referencias
Horwitz B., Husain F. T., Braun A. R., Tagamets M.-A. (2001) Simulating PET/fMRI
studies of visual and auditory pattern recognition using biologically realistic
large-scale neural models. In: Proceedings of the International Joint Conference
on Neural Networks, pp. 878–883. IEEE Press.
Jahr
C. E., Stevens C. F. (1990) Voltage dependence of NMDA-activated
macroscopic conductances predicted by single-channel kinetics. Journal of
Neuroscience, 10:3178-3182.
Jonides J., Schumacher E. H., Smith E. E., Koeppe R. A., Awh E., Reuter-Lorentz P. A.,
Marshuetz C., Willis C. R. (1998) The role of parietal cortex in verbal working
memory. Journal of Neuroscience, 18:5026-5034.
Kastner S., Ungerleider L. G. (2000) Mechanisms of visual attention in the human
cortex. Annual Review of Neuroscience, 23:315-341.
Kamitami Y., Tong F. (2005) Decoding the visual and subjective contents of the
human brain. Nature Neuroscience, 8:679-685.
Kubota K., Niki H. (1971) Prefrontal cortical unit activity and delayed alternation
performance in monkeys. Journal of Neurophysiology, 34:337-347.
Lorente de Nó R. (1938) The cerebral cortex: architecture, intracortical
connections and motor projections. In: Fulton J. F., Physiology of the Nervous
System, pp. 291-339. Oxford University Press, London.
Luck S. J., Chelazzi L., Hillyard S. A., Desimone R. (1997) Neural mechanisms of
spatial selective attention in areas V1, V2 and V4 of macaque visual cortex.
Journal of Neurophysiology, 77:24-42.
Martinez-Trujillo J. C., Treue S. (2002) Attentional modulation strength in cortical
area MT depends on stimulus contrast. Neuron, 35:365-370.
Martinez-Trujillo J. C., Treue S. (2004) Feature-based attention increases the
selectivity of population responses in primate visual cortex. Current Biology,
14:744-751.
Maunsell J. H. R., Van Essen D. C. (1983) Functional properties of neurons in middle
temporal visual area of the macaque monkey. I. Selectivity for stimulus
direction, speed, and orientation. Journal of Neurophysiology, 49:1127-1147.
McAdams C. J., Maunsell J. H. (1999) Effects of attention on orientation-tuning
functions of single neurons in macaque cortical area V4. Journal of
Neuroscience, 19:431-441.
Mesulam M. M. (1981) A cortical network for directed attention and unilateral
neglect. Annals of Neurology, 10:309-325.
Mesulam M. M. (1990) Large-scale neurocognitive networks and distributed
processing for attention, language and memory. Annals of Neurology, 28:597613.
55
Mecanismos de modulación por atención selectiva entre áreas corticales: Un modelo biofísico de red
Mikami A., Newsome W. T., Wurtz R. H. (1986) Motion selectivity in macaque visual
cortex. I. Mechanisms of direction and speed selectivity in extrastriate area
MT. Journal of Neurophysiology, 55:1308-1327.
Moran J., Desimone R. (1985) Selective attention gates visual processing in the
extrastriate cortex. Science, 229:782.4.
Motter B. C. (1993) Focal attention produces spatially selective processing in
visual cortical areas V1, V2, and V4 in the presence of competing stimuli.
Journal of Neurophysiology, 70:909-919.
Motter B. C. (1994) Neuronal correlates of attentive selection for color or
luminance in extrastriate area V4. Journal of Neuroscience, 14:2178-2189.
Mountcastle V. B. (1957) Modality and topographic properties of single neurons of
cat's somatic sensory cortex. Journal of Neurophysiology. 20:408-434.
Mountcastle V. B. (1997) The columnar organization of the neocortex. Brain
120:701-722.
Murphy B. K., Miller K. D. (2003) Multiplicative gain changes are induced by
excitation or inhibition alone. Journal of Neuroscience, 23:10040-10051.
O'Craven K. M., Rosen B. R., Kwong K. K., Treisman A., Savoy R. L. (1997) Voluntary
attention modulates fRMI activity in human MT-MST. Neuron, 18:591.8.
Ó Scalaidhe S. P., Goldman-Rakic P. S. (1999) Functional organization of prefrontal
cortical columns in the macaque. Society for Neuroscience Abstract, 25:620.2.
Pessoa L., Kastner S., Ungerleider L. G. (2002a) Attentional control of the processing
of neural and emotional stimuli. Cognitive Brain Research, 15:31-45.
Pessoa L., Gutierrez E., Bandettini P. B., Ungerleider L. G. (2002b) Neural correlates of
visual working memory: fRMI amplitude predicts task performance. Neuron,
35:975-987.
Pessoa L., Ungerleider L. G. (2004) Top-down mechanisms for working memory and
attentional processes. In: Gazzaniga M. S., Ed. The New Cognitive Neurosciences,
3rd Edition, pp 919-930, MIT Press.
Petit L., Courtney S. M., Ungerleider L. G., Haxby J. V. (1998) Sustained activity in the
medial wall during working memory delays. Journal of Neuroscience, 18:94299437.
Posner M. I., Fan J. (2004) Attention as an organ system. In: Pomerantz J. R., Crai M.
C., Eds. Topics in integrative neuroscience: from cells to cognition. Cambridge:
Cambridge University Press.
Posner M. I., Petersen S. E. (1990) The attention system of the human brain. Annual
Review of Neuroscience, 13:25-42.
56
Referencias
Posner M. I. (2004) Tenth annual George A. Miller distinguished lecture at the
annual meeting of the Cognitive Neuroscience Society. San Francisco,
Cognitive Neuroscience Society.
Purushothaman G., Bradley D. C. (2005) Neural population code for fine perceptual
decisions in area MT. Nature Neuroscience, 8:99-106.
Qian N., Andersen R. A. (1994) Transparent motion perception as detection of
unbalanced motion signals. II. Physiology. Journal of Neuroscience, 14:73677380.
Rao S. G., Williams G. V., Goldman-Rakic P. S. (1999) Isodirectional tuning of
adjacent interneurons and pyramidal cells during working memory:
evidence for microcolumnar organization in PFC. Journal of Neurophysiology,
81:1903-1916.
Raz A. (2004) Anatomy of attentional networks. Anatomical record (part B: new
anat.), 281B:21-36.
Remington R. W., Johnson J. C., Yantis S. (1992) Involuntary attentional capture by
abrupt onset. Perception & Psychophysics, 51:279-290.
Renart A., Brunel N., Wang X-J (2003) Mean-Field theory of irregularly spiking
neuronal populations and working memory in recurrent cortical networks.
In: Computational Neuroscience: A Comprehensive Approach (Feng J., Ed.), chap.
15, pp. 431-490. CRC Press, Boca Raton.
Rensink R. A., O'Regan J. K., Clark J. J. (1997) To see or not to see: the need for
attention to perceive changes in scenes. Psychol. Sci., 8:368-373.
Reynolds J. H., Chelazzi L., Desimone R. (1999) Competitive mechanisms subserve
attention in macaque areas V2 and V4. Journal of Neuroscience, 19:1736-1753.
Reynolds J. H., Desimone R. (1997) Attention and contrast have similar effects on
competitive interactions in macaque area V4. Society for Neuroscience
Abstract, 23:302.
Reynolds J. H., Pasternak T., Desimone R. (2000) Attention increases sensitivity of
V4 neurons. Neuron, 26:703-714.
Rosenkilde C. E., Bauer R. H., Fuster J. M. (1981) Single cell activity in ventral
prefrontal cortex of behaving monkeys. Brain Research, 209:375-394.
Rowe J. B., Toni I., Josephs O., Frackowiak R. S., Passingham R. E. (2000) The
prefrontal cortex: Response selection or maintenance within working
memory? Science, 288:1656-1660
Saenz M., Buracas G. T., Boynton G. M. (2002) Global effects of feature-based
attention in human visual cortex. Nature Neuroscience, 5:631.2.
Salin P. A., Prince D. A. (1996) Spontaneous GABAA receptor mediated inhibitory
currents in adult rat somatosensory cortex. Journal of Neurophysiology,
75:1573-1588.
57
Mecanismos de modulación por atención selectiva entre áreas corticales: Un modelo biofísico de red
Salzman C. D., Britten K. H., Newsome W. T. (1990) Cortical microstimulation
influences perceptual judgements of motion direction. Nature, 346:174.7.
Shipp S., Zeki S. (1985) Segregated output to area V5 from layer 4B of macaque
monkey striate cortex. Journal of Physiology (London), 369:32P.
Simoncelli E. P., Heeger D. J. (1998) A model of neuronal responses in visual area
MT. Vision Research, 38:743-761.
Snowden R. J., Treue S., Erikson R. G., Andersen R. A. (1991) The response of area MT
and V1 to transparent motion. Journal of Neuroscience, 11:2768-2785.
Snowden R. J., Treue S., Andersen R. A. (1992) The response of neurons in areas V1
and MT of the alert rhesus monkey to moving random dot patterns.
Experimental Brain Research, 88:389-400.
Spruston N., Jonas P., Sakmann B. (1995) Dendritic glutamate receptor channel in
rat hippocampal CA3 and CA1 pyramidal neurons. Journal of Physiology
(London), 482:325-352.
Treue S., Martinez-Trujillo J. C. (1999) Feature-based attention influences motion
processing gain in macaque visual cortex. Nature, 399:575-.9.
Treue S., Hol K., Rauber S. J. (2000) Seeing multiple directions of motionphisiology and psychophysics. Nature Neuroscience, 3:270.6.
Troyer T. W., Miller K. D. (1997) Physiological gain leads to high ISI variability in a
simple model of a cortical regular spiking cell. Neural Computation, 9:971-983.
Tuckwell H. C. (1988) Introduction to theoretical neurobiology.
Cambridge University Press.
Cambridge:
Ungerleider L. G., Gaffan D., Pelak V. S. (1989) Projections from inferior temporal
cortex to prefrontal cortex via the uncinate fascicle in rhesus monkeys.
Experimental Brain Research, 76:473-484.
Valdes-Sosa M., Bobes M. A., Rodriguez V., Pinilla T. (1998) Switching attention
without shifting the spotlight: Object-based attentional modulation of brain
potentials. Journal of Cognitive Neuroscience, 10:137-151.
Wang X-J (1999) Synaptic basis of cortical persistent activity: the importance of
NMDA receptors to working memory. Journal of Neuroscience, 19:9587-9603.
Watanabe M. (1986) Prefrontal unit activity during delayed conditional go/no-go
discrimination in the monkey I. Relation to the stimulus. Brain Research,
382:1-14.
Webster M. J., Bachevelier J., Ungerleider L. G. (1993) Subcortical connections of
inferior temporal areas TE and TEO in macaques. Journal of Comparative
Neurology, 335:73-91.
58
Referencias
Webster M. J., Bachevelier J., Ungerleider L. G. (1994) Connections of inferior
temporal areas TEO and TE with parietal and frontal cortex in macaque
monkeys. Cerebral Cortex, 4:470-483.
Xiang Z., Huguenard J. R., Prince D. A. (1998) GABAA receptor mediated currents in
interneurons and pyramidal cells of rat visual cortex. Journal of Physiology
(London), 506:715-730.
Yantis S., Johnson D.N. (1990) Mechanisms of attentional priority. Journal of
Experimental Psychology: Human Perception & Performance, 16:812-825.
Zeki S. M. (1974) Functional organization of a visual area in the posterior bank of
the superior temporal sulcus of the rhesus monkey. Journal of Physiology
(London), 263:549-573
59
 0
0
Anuncio


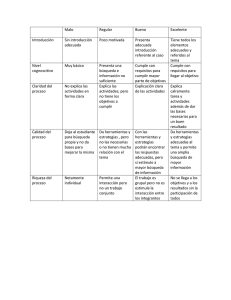

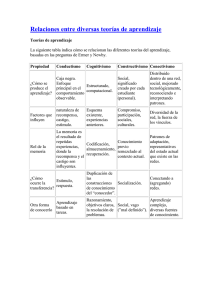

Descargar
Anuncio
Añadir este documento a la recogida (s)
Puede agregar este documento a su colección de estudio (s)
Iniciar sesión Disponible sólo para usuarios autorizadosAñadir a este documento guardado
Puede agregar este documento a su lista guardada
Iniciar sesión Disponible sólo para usuarios autorizados